centro activo AMP sÃtio alostérico AMP
centro activo AMP sÃtio alostérico AMP
centro activo AMP sÃtio alostérico AMP

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
Um modelo possível para explicar a inibição de tipo não competitivo é admitir a<br />
existência na enzima de um local de ligação diferente do <strong>centro</strong> <strong>activo</strong> cuja<br />
ligação ao inibidor impedisse a formação do produto.<br />
A par da classificação que divide os inibidores em competitivos e não<br />
competitivos existe uma outra que divide os modificadores (inibidores ou<br />
activadores) em isostéticos ou alostéricos.<br />
Um modificador isostérico liga-se ao<br />
<strong>centro</strong> <strong>activo</strong> e é sempre inibidor:<br />
é um inibidor isostérico.<br />
Um ligando alostérico liga-se à enzima num sítio diferente do <strong>centro</strong> <strong>activo</strong> (um<br />
sítio alostérico) provocando uma alteração conformacional na enzima.<br />
Sítios alostéricos<br />
Inibidor alostérico<br />
Se a alteração<br />
conformacional diminuir a<br />
actividade da enzima<br />
dizemos que há inibição<br />
alostérica.<br />
De acordo com este modelo tudo se passaria como se<br />
as moléculas de enzima ligadas ao inibidor estivessem excluídas do processo<br />
catalítico, isto é, a concentração de enzima tivesse baixado.<br />
37<br />
Activador alostérico<br />
Se a alteração<br />
conformacional aumentar a<br />
actividade da enzima<br />
dizemos que há activação 38<br />
alostérica.<br />
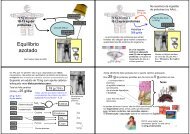
Quando uma fibra muscular se contrai aumenta a velocidade de hidrólise do<br />
ATP e aumenta a concentração de ADP (e <strong>AMP</strong>; 2 ADP → <strong>AMP</strong> + ATP).<br />
O <strong>AMP</strong> liga-se à fosforílase muscular induzindo uma conformação mais activa<br />
⇔ o <strong>AMP</strong> é um activador alostérico da fosforílase muscular.<br />
Fosforílase<br />
muscular na<br />
conformação<br />
tensa (inactiva)<br />
<strong>AMP</strong><br />
sítio alostérico<br />
Fosforílase<br />
muscular na<br />
conformação<br />
relaxada (activa)<br />
A cínase-1 da frutose-6-P (ATP+ Frutose-6-P→ ADP + Frutose-1,6-bisfosfato)<br />
o ATP é, simultaneamente, substrato e inibidor alostérico. Para concentrações<br />
fisiológicas de ATP, o <strong>AMP</strong> é um activador alostérico.<br />
(3) O <strong>AMP</strong><br />
compete com o<br />
ATP pelo mesmo<br />
sítio alostérico<br />
impedindo a sua<br />
acção inibidora.<br />
<strong>AMP</strong><br />
<strong>centro</strong> <strong>activo</strong><br />
39<br />
(1) Um dos substratos da cínase da frutose-6-P é o ATP.<br />
(2) Contudo, existe na<br />
enzima um<br />
sítio alostérico onde o<br />
ATP se liga inibindo-a.<br />
Para concentrações<br />
fisiológicas de ATP essa<br />
acção inibidora é muito<br />
40<br />
marcada.
O receptor da insulina pode ser visto como uma enzima alostérica com o seu<br />
<strong>centro</strong> alostérico activador virado para o lado extra-celular (onde se liga a<br />
insulina) e o <strong>centro</strong> <strong>activo</strong> no lado citoplasmático.<br />
O receptor da insulina é uma<br />
cínase (ATP + proteína →<br />
ADP + proteína-P) que<br />
catalisa a fosforilação de<br />
proteínas específicas (ISR –<br />
“insulin receptor substrates”)<br />
num resíduo de tirosina.<br />
Os IRS fosforilados pela<br />
acção catalítica do receptor<br />
da insulina vão funcionar<br />
como activadores alostéricos<br />
de outras enzimas estando na<br />
origem de uma cadeia de<br />
reacções que estão na base<br />
dos efeitos da insulina.<br />
A ligação<br />
entre os substratos e o <strong>centro</strong> <strong>activo</strong><br />
entre os inibidores competitivos e o <strong>centro</strong> <strong>activo</strong> ou<br />
entre os modificadores alostéricos e os sítios alostéricos<br />
é de tipo não covalente e reversível.<br />
sítio alostérico<br />
<strong>AMP</strong><br />
41<br />
42<br />
Um ligando diferente do substrato que se liga ao <strong>centro</strong> <strong>activo</strong> é um inibidor<br />
isostérico (se se liga no <strong>centro</strong> <strong>activo</strong> só pode inibir).<br />
Se a ligação entre um inibidor isostérico e o <strong>centro</strong> <strong>activo</strong><br />
for reversível (se poder se “deslocado” pelo substrato)<br />
esse inibidor comporta-se funcionalmente como inibidor competitivo<br />
... mas se a ligação do inibidor ao sítio <strong>activo</strong> for irreversível<br />
(não podendo ser “deslocado” por altas concentrações de substrato)<br />
as moléculas de enzima ligadas ao inibidor ficam excluídas do processo catalítico.<br />
A lípase pancreática catalisa a<br />
hidrólise de triacilgliceróis no<br />
intestino.<br />
No tratamento da obesidade pode<br />
usar-se um fármaco (orlistat; xenical)<br />
que é um inibidor da lípase<br />
pancreática.<br />
Neste caso o inibidor comporta-se<br />
funcionalmente como<br />
não competitivo.<br />
43<br />
O orlistat reage com uma serina<br />
(formando uma ligação covalente e<br />
irreversível)<br />
situada no <strong>centro</strong> <strong>activo</strong> da enzima<br />
bloqueando a sua actividade.<br />
44
A modificação da actividade de uma enzima pode envolver a sua modificação<br />
covalente (hidrólise irreversível ou fosforilação reversível) por acção catalítica de<br />
enzimas.<br />
Muitas enzimas são reguladas por mecanismos de fosforilação/desfosforilação<br />
catalisadas por enzimas (cínases e fosfátases).<br />
A fosfátase da desidrogénase<br />
do piruvato<br />
catalisa a desfosforilação da<br />
desidrogénase do piruvato<br />
que no estado desfosforilado<br />
fica activa.<br />
Forma desfosforilada<br />
(activa)<br />
A cínase da desidrogénase do<br />
piruvato<br />
Certas enzimas são activadas por hidrólise irreversível.<br />
São exemplos a activação dos zimogénios na digestão dos<br />
Forma fosforilada<br />
catalisa a fosforilação da<br />
desidrogénase do piruvato<br />
nutrientes. (inactiva)<br />
45<br />
que no estado fosforilado 46 fica<br />
inactiva.<br />
A transição do estado de saciedade para o de jejum induz aumento da glicagina que,<br />
ligando-se ao seu receptor nos hepatócitos, induz a síntese de <strong>AMP</strong>c, que activa a<br />
PKA, que promove a fosforilação de muitas enzimas: síntase do glicogénio, cínase do<br />
piruvato, fosfátase-1 de proteínas, cínase da fosforílase do glicogénio, enzima<br />
bifuncional (cínase-2 da frutose-6-P; frutose-2,6-bisfosfátase), etc.<br />
Nalguns casos, a modificação da<br />
actividade de uma enzima (se o seu<br />
substrato é outra enzima) pode<br />
depender da modificação alostérica<br />
do substrato.<br />
glicose<br />
fosfoenolpiruvato<br />
ADP<br />
ATP<br />
piruvato<br />
Se pensarmos que o papel da<br />
glicagina é<br />
(1) promover a formação de<br />
glicose pelo fígado<br />
(2) inibir o seu consumo neste<br />
órgão...<br />
podemos deduzir qual o efeito da<br />
fosforilação catalisada pela PKA<br />
numa determinada enzima...<br />
A cínase do piruvato hepática é inibida<br />
por fosforilação mas...<br />
a isoenzima muscular não é substrato da<br />
PKA nem é regulada por 47<br />
fosforilação/desfosforilação.<br />
A fosforílase do glicogénio é<br />
activada por fosforilação e<br />
inactivada por desfosforilação que é<br />
catalisada pela fosfátase-1.<br />
A actividade fosfatásica da<br />
fosfátase-1 na fosforílase do<br />
glicogénio aumenta quando a<br />
glicose se liga à fosforílase em<br />
sítios alostéricos.<br />
48
A regulação de uma via metabólica pode envolver activações e inibições<br />
alostéricas e/ou isostéricas assim como activações e inibições por fosforilação<br />
e desfosforilação reversível que operam em cadeia.<br />
O <strong>AMP</strong><br />
é activador alostérico<br />
de uma cínase<br />
que inactiva a<br />
carboxílase de acetil-<br />
CoA<br />
o que baixa a<br />
concentração de<br />
malonil-CoA.<br />
A descida do malonil-<br />
CoA “desinibe” a<br />
carnitina-palmitoil<br />
transférase I<br />
A carnitina-palmitoil<br />
transférase I é a enzima reguladora da oxidação dos ácidos gordos.<br />
49<br />
Desejo a todos os alunos<br />
as maiores felicidades<br />
na Frequência de Bioquímica<br />
e nas outras frequências.<br />
50