Il potenziale di membrana - CPRG
Il potenziale di membrana - CPRG Il potenziale di membrana - CPRG
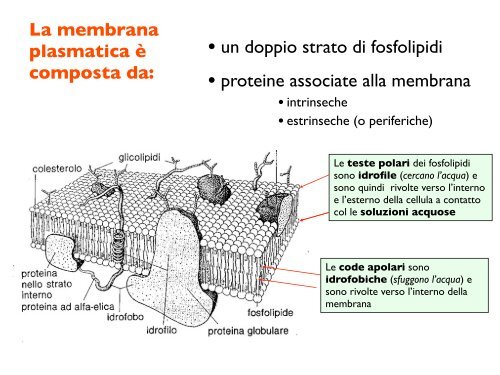
La membrana plasmatica è composta da: • un doppio strato di fosfolipidi • proteine associate alla membrana • intrinseche • estrinseche (o periferiche) Le teste polari dei fosfolipidi sono idrofile (cercano l’acqua) e sono quindi rivolte verso l’interno e l’esterno della cellula a contatto col le soluzioni acquose Le code apolari sono idrofobiche (sfuggono l’acqua) e sono rivolte verso l’interno della membrana
- Page 2 and 3: Le proteine intrinseche sono blocca
- Page 4 and 5: Che cosa sono gli ioni Quando un sa
- Page 7 and 8: Gli ioni che hanno maggior importan
- Page 9 and 10: Ci sono vari tipi di canali ad acce
- Page 11 and 12: i 4 livelli strutturali delle prote
- Page 13 and 14: Metodi di studio dei canali ionici
- Page 15 and 16: Immunocitochimica È possibile otte
- Page 17 and 18: Basi ioniche dei potenziali Tutte l
- Page 19 and 20: Lo stesso accade quando rendiamo un
- Page 21 and 22: La forze elettriche La forze elettr
- Page 23 and 24: Il flusso di corrente in un sistema
- Page 25 and 26: Nei neuroni come in tutte le cellul
- Page 27 and 28: Il rapporto scritto sopra la frecci
- Page 29 and 30: La maggior parte dei fenomeni che a
- Page 31 and 32: Si supponga ad esempio che inizialm
- Page 33 and 34: Il potenziale di equilibrio per uno
- Page 35 and 36: EK = 61,54 mV x ln 1 20 = -80 mV ne
- Page 37: Il potenziale di membrana (indicato
La <strong>membrana</strong><br />
plasmatica è<br />
composta da:<br />
• un doppio strato <strong>di</strong> fosfolipi<strong>di</strong><br />
• proteine associate alla <strong>membrana</strong><br />
• intrinseche<br />
• estrinseche (o periferiche)<br />
Le teste polari dei fosfolipi<strong>di</strong><br />
sono idrofile (cercano l’acqua) e<br />
sono quin<strong>di</strong> rivolte verso l’interno<br />
e l’esterno della cellula a contatto<br />
col le soluzioni acquose<br />
Le code apolari sono<br />
idrofobiche (sfuggono l’acqua) e<br />
sono rivolte verso l’interno della<br />
<strong>membrana</strong>
Le proteine intrinseche sono<br />
bloccate all’interno della<br />
<strong>membrana</strong> perché la<br />
porzione che attraversa la<br />
<strong>membrana</strong> ha residui<br />
aminoaci<strong>di</strong>ci apolari che non<br />
possono uscire a contatto<br />
con il mezzo acquoso<br />
si<br />
no<br />
si<br />
Nella maggior parte delle cellule le<br />
proteine intrinseche possono<br />
muoversi in orizzontale<br />
liberamente scorrendo dentro la<br />
<strong>membrana</strong><br />
no<br />
Nei neuroni tuttavia la maggior parte delle proteine <strong>di</strong> <strong>membrana</strong><br />
sono fissate in posizione dato che porzioni <strong>di</strong>fferenti del neurone hanno<br />
composizione proteica e proprietà <strong>di</strong>fferenti
Caratteristiche <strong>di</strong> permeabilità del doppio strato fosfolipi<strong>di</strong>co<br />
In una <strong>membrana</strong> artificiale<br />
costituita da soli fosfolipi<strong>di</strong>:<br />
NB: La scala è logaritmica per<br />
cui quando ad esempio si passa da<br />
10 -8 a 10 -6 la permeabilità<br />
<strong>di</strong>venta 100 volte maggiore<br />
• Passano facilmente i gas, le<br />
molecole liposolubili e<br />
idrofobiche (O 2 , lipi<strong>di</strong>)<br />
• Passano più lentamente le<br />
molecole polari (H 2 O, CO 2 )<br />
• Non passano le molecole<br />
cariche (ioni: K + , Na + , Cl - )<br />
• A parità <strong>di</strong> solubilità nei lipi<strong>di</strong> le<br />
molecole piccole passano più<br />
facilmente delle gran<strong>di</strong> (H 2 O<br />
<strong>di</strong>ffonde più rapidamente <strong>di</strong><br />
monosaccari<strong>di</strong> o aminoaci<strong>di</strong>)
Che cosa sono gli ioni<br />
Quando un sale, ad esempio cloruro <strong>di</strong> so<strong>di</strong>o (NaCl), il comune sale<br />
da cucina, viene <strong>di</strong>sciolto in acqua, le sue molecole si <strong>di</strong>ssociano in ioni<br />
Na + Cl -<br />
Nel sale NaCl il legame chimico è molto forte (in quanto le<br />
cariche opposte <strong>di</strong> Na + e Cl - si attraggono)<br />
La separazione dei due ioni in soluzione è resa possibile dal fatto che<br />
in sostituzione del legame forte tra Na + e Cl - si formano<br />
tanti legami deboli tra le molecole <strong>di</strong> acqua e gli ioni (il<br />
cosiddetto guscio <strong>di</strong> idratazione)
L’acqua infatti è un <strong>di</strong>polo cioè<br />
una molecola con una parziale carica<br />
positiva e una parziale carica negativa<br />
in grado quin<strong>di</strong> <strong>di</strong><br />
formare legami H (+)<br />
sia con Na +<br />
(-)<br />
O<br />
che con Cl -<br />
H<br />
H<br />
O<br />
O H H<br />
Na +<br />
O H H<br />
O H H<br />
H (+) O H H<br />
O H H<br />
Cl -<br />
H<br />
H<br />
H<br />
H<br />
O<br />
O
Gli ioni che hanno maggior importanza<br />
per la fisiologia del neurone:<br />
Cationi<br />
carica + verso il catodo<br />
Anioni<br />
carica - verso l’anodo<br />
K + Na + Ca ++ Cl -<br />
Potassio<br />
Cloro<br />
So<strong>di</strong>o<br />
Calcio
I canali ionici:<br />
ØPermettono il passaggio degli ioni<br />
ØSono estemamente selettivi (lasciano passare <strong>di</strong><br />
solito un solo tipo <strong>di</strong> ione)<br />
Alcuni canali sono<br />
sempre aperti<br />
Canali<br />
passivi<br />
Altri si aprono e si<br />
chiudono in risposta a<br />
determinati stimoli<br />
Canali ad<br />
accesso<br />
variabile
Ci sono vari tipi <strong>di</strong> canali ad accesso variabile<br />
Alcuni variano la loro permeabilità a seconda<br />
della presenza all’esterno <strong>di</strong> messaggeri<br />
chimici (ormoni, neurotrasmettitori)<br />
Est<br />
Int<br />
Alcuni variano la loro permeabilità in risposta ad<br />
un messaggero intracellulare<br />
(II° messaggero)<br />
Alcuni variano la loro permeabilità quando vi è una<br />
variazione del voltaggio<br />
Alcuni variano la loro permeabilità in risposta ad<br />
una sollecitazione meccanica sulla cellula<br />
(me<strong>di</strong>ata dal citoscheletro)
I canali ionici<br />
I canali ionici sono<br />
proteine <strong>di</strong> <strong>membrana</strong><br />
Essi sono troppo<br />
piccoli per essere<br />
stu<strong>di</strong>ati con meto<strong>di</strong><br />
tra<strong>di</strong>zionali<br />
Tuttavia:<br />
Est<br />
Int<br />
5 nanometri<br />
5 nanometri<br />
6 nanometri<br />
2 nanometri<br />
Appartengono a non più <strong>di</strong> 3-4 <strong>di</strong>verse famiglie e i <strong>di</strong>versi tipi <strong>di</strong> canali si<br />
assomigliano nella struttura<br />
Sono ben conservati a livello filogenetico: lo stesso canale si può trovare<br />
nella drosophila (moscerino della frutta) o nell’uomo
i 4 livelli strutturali delle proteine:<br />
struttura primaria: aminoaci<strong>di</strong> legati tra loro da legami pepti<strong>di</strong>ci (carbossilicoaminico)<br />
struttura secondaria: <strong>di</strong>stribuzione alfa-elica<br />
struttura terziaria: la proteina assume forma globulare, il polipeptide si piega<br />
su se stesso<br />
struttura quaternaria: pepti<strong>di</strong> <strong>di</strong>fferenti si legano tra loro creando una<br />
proteina <strong>di</strong> <strong>di</strong>mensioni maggiori<br />
11
I canali ionici sono composti da più sub-unità<br />
(<strong>di</strong> solito da 4 a 6)<br />
La porzione che attraversa<br />
la <strong>membrana</strong> ha una<br />
struttura ad α-elica con<br />
aminoaci<strong>di</strong> polari rivolti<br />
verso il canale e aminoaci<strong>di</strong><br />
apolari rivolti verso lo<br />
strato fosfolipi<strong>di</strong>co
Meto<strong>di</strong> <strong>di</strong> stu<strong>di</strong>o dei canali ionici<br />
La sequenza degli amminoaci<strong>di</strong> ci può dare molte<br />
informazioni<br />
La struttura secondaria ci<br />
può dare informazione sulla<br />
forma del canale<br />
La mappa <strong>di</strong> idrofobicità<br />
ci da in<strong>di</strong>zi sulla struttura e<br />
sulla funzione dei canali<br />
È anche possibile causare mutazioni mirate in certe porzioni del<br />
canale per vedere qual è la funzione <strong>di</strong> quella porzione
Patch-clamp<br />
È possibile stu<strong>di</strong>are il<br />
comportamento elettrico<br />
<strong>di</strong> un solo canale per<br />
mezzo del metodo del<br />
patch-clamp
Immunocitochimica<br />
È possibile ottenere anticorpi che rispondono selettivamente ad un<br />
canale ionico o a sue porzioni<br />
Questi anticorpi vengono poi marcati (con ra<strong>di</strong>oattivo o con coloranti<br />
speciali) e si va a vedere dove gli anticorpi si vanno ad attaccare a livello<br />
<strong>di</strong> una cellula o <strong>di</strong> un tessuto
<strong>Il</strong> <strong>potenziale</strong> <strong>di</strong> <strong>membrana</strong><br />
Nel citoplasma della cellula, così come nel liquido extracellulare<br />
sono <strong>di</strong>sciolti <strong>di</strong>versi tipi <strong>di</strong> ioni. La loro concentrazione non è però<br />
uguale dai due lati della <strong>membrana</strong> citoplasmatica<br />
<strong>Il</strong> <strong>potenziale</strong> <strong>di</strong> <strong>membrana</strong> ( Vm ) corrisponde alla <strong>di</strong>fferenza<br />
<strong>di</strong> <strong>potenziale</strong> elettrico tra interno ed esterno della cellula, cioè alla<br />
<strong>di</strong>fferenza tra interno ed esterno nel numero <strong>di</strong> cariche positive e<br />
negative portate dagli ioni<br />
I segnali trasmessi da sistema nervoso sono costituiti<br />
da rapide variazioni del <strong>potenziale</strong> <strong>di</strong> <strong>membrana</strong> delle<br />
cellule nervose
Basi ioniche dei potenziali<br />
Tutte le attività del sistema nervoso, dalla generazione dei<br />
potenziali <strong>di</strong> <strong>membrana</strong> alla trasmissione sinaptica,<br />
<strong>di</strong>pendono strettamente dal flusso dei <strong>di</strong>versi tipi <strong>di</strong> ioni<br />
attraverso i flui<strong>di</strong> come il citoplasma e il liquido<br />
extracellulare e attraverso la <strong>membrana</strong><br />
citoplasmatica<br />
<strong>Il</strong> flusso degli ioni è<br />
regolato da due<br />
meccanismi:<br />
• La <strong>di</strong>ffusione<br />
• Le forze elettriche
La <strong>di</strong>ffusione<br />
La <strong>di</strong>ffusione regola il movimento <strong>di</strong> tutte le molecole<br />
solubili (e quin<strong>di</strong> non solo gli ioni) e consiste nello spostamento<br />
delle molecole dalla zona dove esse sono più concentrate alla<br />
zona dove sono meno concentrate<br />
Se in una stanza è riunito un numero<br />
consistente <strong>di</strong> persone e ad un certo<br />
punto apriamo la porta <strong>di</strong><br />
comunicazione col la stanza attigua, le<br />
persone tenderanno a <strong>di</strong>stribuirsi<br />
equamente nelle due stanze migrando<br />
dalla zona a maggior densità a quella a<br />
densità minore
Lo stesso accade quando ren<strong>di</strong>amo una<br />
<strong>membrana</strong> permeabile agli ioni
In una cellula nervosa la <strong>di</strong>ffusione<br />
<strong>di</strong>pende da:<br />
• <strong>Il</strong> gra<strong>di</strong>ente <strong>di</strong> concentrazione <strong>di</strong><br />
quella molecola (quanto <strong>di</strong>fferente è la concentrazione<br />
dai due lati)<br />
• La permeabilità della <strong>membrana</strong> nei<br />
confronti <strong>di</strong> quella molecola<br />
Nell’esempio precedente la <strong>di</strong>ffusione <strong>di</strong>penderebbe 1) da quanto affollata<br />
la stanza prima che si apra la porta e 2) da quanto grande è la porta che<br />
viene aperta
La forze elettriche<br />
La forze elettriche influenzano il movimento solamente degli ioni<br />
Esse <strong>di</strong>pendono da:<br />
• La <strong>di</strong>fferenza <strong>di</strong> <strong>potenziale</strong> (o voltaggio)<br />
• La conduttanza elettrica (o come spesso si<br />
preferisce, dal suo reciproco, la resistenza)
la legge <strong>di</strong> Ohm<br />
relazione tra la <strong>di</strong>fferenza <strong>di</strong> <strong>potenziale</strong> ai capi <strong>di</strong> un<br />
conduttore elettrico e la corrente che lo attraversa
<strong>Il</strong> flusso <strong>di</strong> corrente in un sistema<br />
segue la legge <strong>di</strong> Ohm<br />
Flusso <strong>di</strong><br />
corrente (si<br />
misura in Ampere)<br />
I = g x V<br />
Voltaggio o<br />
<strong>di</strong>fferenza <strong>di</strong><br />
<strong>potenziale</strong> (si<br />
misura in Volt)<br />
Conduttanza (si<br />
misura in Siemens)<br />
Spesso la legge <strong>di</strong> Ohm viene<br />
espressa rispetto alla resistenza<br />
(R=V/I)<br />
La resistenza è semplicemente<br />
il reciproco della conduttanza<br />
g =1/R<br />
I = V / R<br />
Resistenza<br />
(si misura in<br />
Ohm)
Me<strong>di</strong>ante la legge <strong>di</strong> Ohm, possiamo prevedere il<br />
comportamento <strong>di</strong> uno ione soggetto a forze elettriche<br />
I = g x V<br />
In una cellula nervosa il voltaggio è legato al valore del<br />
<strong>potenziale</strong> <strong>di</strong> <strong>membrana</strong><br />
La conduttanza elettrica è invece proporzionale al numero <strong>di</strong><br />
canali ionici (aperti) presenti nella <strong>membrana</strong><br />
Se la <strong>membrana</strong> non è<br />
permeabile agli ioni (g = 0)<br />
oppure se la <strong>di</strong>fferenza <strong>di</strong><br />
<strong>potenziale</strong> è nulla (V = 0) non<br />
ci sarà flusso <strong>di</strong> corrente (in<br />
entrambi i casi I <strong>di</strong>venta 0)
Nei neuroni come in tutte le cellule dell’organismo ci sono gran<strong>di</strong> <strong>di</strong>fferenze<br />
nella concentrazione <strong>di</strong> ciascun ione tra interno ed esterno della cellula<br />
La <strong>di</strong>fferenza <strong>di</strong> concentrazione degli ioni tra interno ed esterno della<br />
cellula è mantenuta dal lavoro incessante della pompa so<strong>di</strong>o-potassio<br />
(e da quello delle altre pompe, pompa del calcio, pompa del cloro)
Ad ogni ciclo, la pompa so<strong>di</strong>o-potassio<br />
- espelle tre ioni Na + (So<strong>di</strong>o)<br />
- trasporta all’interno due ioni K + (Potassio)<br />
consumando una molecola <strong>di</strong> ATP<br />
L’80% dell’ATP consumato dal neurone è dovuto al funzionamento<br />
delle pompe ioniche
<strong>Il</strong> rapporto scritto sopra la<br />
freccia in<strong>di</strong>ca quanto<br />
maggiormente è<br />
concentrato uno ione da un<br />
lato rispetto all’altro<br />
La freccia in<strong>di</strong>ca la <strong>di</strong>rezione<br />
in cui gli ioni sono sospinti<br />
dal loro gra<strong>di</strong>ente <strong>di</strong><br />
concentrazione<br />
K + K +<br />
Na +<br />
Cl - 20:1<br />
Ca ++<br />
1:10<br />
1:11<br />
1:10.000<br />
Na +<br />
Cl -<br />
Ca ++
Ci sono 4 ioni importanti:<br />
K + , Na + , Ca ++ , Cl -<br />
Tre sono più concentrati all’esterno,<br />
uno (K + ) all’interno
La maggior parte dei fenomeni che avvengono<br />
nel neurone possono essere spiegati stu<strong>di</strong>ando il<br />
comportamento <strong>di</strong> due soli ioni , K + e Na +<br />
<strong>Il</strong> potassio, K + è uno ione positivo (catione). Dato che è<br />
molto più concentrato all’interno tenderà ad uscire<br />
rendendo l’interno della cellula ancor più negativo<br />
Anche il so<strong>di</strong>o, Na + è uno ione positivo (catione). Dato<br />
che è molto più concentrato all’esterno tenderà ad entrare<br />
rendendo l’interno della cellula meno negativo<br />
Le variazioni <strong>di</strong> permeabilità della <strong>membrana</strong> a<br />
questi due ioni sono in grado <strong>di</strong> determinare<br />
gran<strong>di</strong> cambiamenti nel <strong>potenziale</strong> <strong>di</strong> <strong>membrana</strong>
Potenziale <strong>di</strong> equilibrio per uno ione<br />
In molti casi il gra<strong>di</strong>ente elettrico può sospingere uno ione in una<br />
<strong>di</strong>rezione mentre il gra<strong>di</strong>ente <strong>di</strong> concentrazione lo sospinge in<br />
<strong>di</strong>rezione opposta<br />
Quando queste due forze si equivalgono, un ugual numero <strong>di</strong> ioni<br />
entra ed esce dalla cellula e si ha quin<strong>di</strong> un equilibrio. <strong>Il</strong> <strong>potenziale</strong><br />
<strong>di</strong> <strong>membrana</strong> a cui questo avviene si chiama <strong>potenziale</strong> <strong>di</strong><br />
equilibrio per uno ione
Si supponga ad esempio che<br />
inizialmente uno ione sia molto<br />
più concentrato da un lato della<br />
<strong>membrana</strong> e che non vi sia<br />
alcuna <strong>di</strong>fferenza <strong>di</strong> <strong>potenziale</strong><br />
tra i due lati<br />
+<br />
+<br />
+<br />
+<br />
+<br />
+<br />
+ +<br />
Gra<strong>di</strong>ente <strong>di</strong><br />
concentrazione<br />
Gra<strong>di</strong>ente<br />
elettrico<br />
Inizialmente lo ione si muoverà in <strong>di</strong>rezione del<br />
lato dove è meno concentrato. Tuttavia man mano<br />
che lo ione passa, andranno accumulandosi da<br />
quel lato sempre più cariche positive (che saranno<br />
invece sottratte dall’altra parte)<br />
+<br />
+<br />
+<br />
+ +<br />
+ +<br />
-<br />
-<br />
+<br />
+<br />
+
Gra<strong>di</strong>ente <strong>di</strong><br />
concentrazione<br />
Gra<strong>di</strong>ente<br />
elettrico<br />
La <strong>di</strong>fferenza <strong>di</strong> <strong>potenziale</strong> andrà aumentando e inizierà<br />
a ri-sospingere lo ione dentro la cellula<br />
All’equilibrio le due forze si equivarranno senza che si<br />
possa mai raggiungere né ugual concentrazione<br />
dai due lati, né neutralità elettrica<br />
+<br />
+<br />
+<br />
+<br />
+ +<br />
-<br />
-<br />
-<br />
+<br />
+<br />
+<br />
+<br />
+
<strong>Il</strong> <strong>potenziale</strong> <strong>di</strong> equilibrio per uno ione in una data<br />
cellula può essere calcolato me<strong>di</strong>ante una equazione,<br />
l’equazione <strong>di</strong> Nernst, e <strong>di</strong>pende essenzialmente dalle<br />
concentrazioni interna ed esterna <strong>di</strong> quello ione.
Ex =<br />
RT<br />
zF<br />
ln<br />
[X]e<br />
[X]i<br />
Ex <strong>potenziale</strong> <strong>di</strong> equilibrio<br />
R costante dei gas<br />
T temperatura (in Kelvin)<br />
z carica ione<br />
F costante <strong>di</strong> Faraday
EK =<br />
61,54 mV<br />
x ln<br />
1<br />
20<br />
= -80 mV<br />
nei neuroni a riposo, il K + è venti volte più concentrato<br />
all’interno della cellula e il suo <strong>potenziale</strong> <strong>di</strong> equilibrio<br />
(E K ) calcolato con l’equazione <strong>di</strong> Nernst, è <strong>di</strong> –80 mV.<br />
Al contrario Na + è <strong>di</strong>eci volte più concentrato fuori e il suo<br />
<strong>potenziale</strong> <strong>di</strong> equilibrio (E Na ) calcolato con<br />
l’equazione <strong>di</strong> Nernst, è <strong>di</strong> +62 mV.
In molti casi le forze elettriche e la <strong>di</strong>ffusione spingono uno ione<br />
nella stessa <strong>di</strong>rezione<br />
Ad esempio a riposo (Vm = -65 mV) sia la <strong>di</strong>ffusione che le<br />
forze elettriche tendono a sospingere dentro il so<strong>di</strong>o<br />
Viceversa all’apice <strong>di</strong> un <strong>potenziale</strong> d’azione (Vm = +40 mV)<br />
<strong>di</strong>ffusione e le forze elettriche tendono entrambe a sospingere<br />
fuori il potassio
<strong>Il</strong> <strong>potenziale</strong> <strong>di</strong> <strong>membrana</strong> (in<strong>di</strong>cato con Vm) è uguale<br />
alla <strong>di</strong>fferenza <strong>di</strong> <strong>potenziale</strong> tra esterno e interno e cioè alla<br />
<strong>di</strong>fferenza nel numero delle cariche dalle due parti della<br />
<strong>membrana</strong> plasmatica.


