Animali 6 - Liceo Classico Psicopedagogico Cesare Valgimigli
Animali 6 - Liceo Classico Psicopedagogico Cesare Valgimigli
Animali 6 - Liceo Classico Psicopedagogico Cesare Valgimigli

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
1. I Lofoforati presentano affinità sia con i protostomi che con i<br />
deuterostomi.<br />
I Lofoforati sono un raggruppamento di animali che presentano alcuni caratteri comuni ai protostomi ed altri comuni ai<br />
deuterostomi. Essi sono caratterizzati dalla presenza del lofoforo che permette la nutrizione. Quest’ultimo è un organo filtratore,<br />
dotato di una piega anteriore a forma di ferro di cavallo che sorregge tentacoli ciliati e cavi. Il celoma si forma con modalità<br />
intermedie tra quelle dei protostomi e quelle dei deuterostomi.<br />
I Lofoforati presentano il corpo<br />
suddiviso in tre parti.<br />
Due di questi phyla, i più importanti, dal punto di<br />
vista del numero di specie, sono i Briozoi e i<br />
Brachiopodi.<br />
I Lofoforati rappresentano un gruppo di animali<br />
che presentano caratteri comuni. Il loro piano<br />
organizzativo è caratterizzato da tre parti che si<br />
succedono in senso antero-posteriore: il prosoma, il<br />
mesosoma e il metasoma; queste porzioni corporee<br />
in molte specie dispongono di una specifica cavità<br />
celomatica: il protocele, il mesocele e il metacele.<br />
I lofoforati di regola producono un rivestimento<br />
I Briozoi sono simili ai coralli.<br />
Gli organismi appartenenti a questo phylum, tutti<br />
acquatici e per la maggior parte marini, presentano<br />
abitudini coloniali e, al pari di altri animali del gruppo dei<br />
deuterostomi, «abitano» entro involucri secreti dal loro<br />
tegumento (▶figura 1).<br />
corporeo esterno, e costantemente presentano un<br />
intestino con percorso a U, che termina con un<br />
ano prossimo all'apertura orale, ma comunque<br />
esterno alle strutture accessorie di corredo alla<br />
bocca.<br />
Tali strutture specializzate rappresentano il tratto<br />
caratteristico di questi organismi, dal quale deriva il<br />
loro stesso nome; i lofoforati dispongono infatti<br />
di un lofoforo, una corona singola o duplice di<br />
tentacoli ciliati, in grado di muoversi grazie a una<br />
specifica muscolatura disposta intorno al mesocele.<br />
La funzione del lofoforo è quella di catturare gli<br />
organismi planctonici di cui si nutrono i lofoforati;<br />
questi sono infatti animali sessili (cioè fissati a un<br />
substrato e quindi immobili) in fase adulta. Secondo<br />
alcuni autori, i tessuti che rivestono i tentacoli cavi del<br />
lofoforo consentirebbero gli scambi gassosi tra<br />
l'organismo e l'ambiente. Anche nei primitivi<br />
vertebrati acquatici, come vedremo, la struttura<br />
ancestrale devoluta alla raccolta del cibo svolge<br />
un'accessoria funzione respiratoria. Ai lofoforati, che<br />
attualmente comprendono 4500 specie, si ascrivono<br />
tre phyla, tutti praticamente comprendenti organismi<br />
marini, con poche eccezioni rappresentate da forme<br />
adattate all'acqua dolce.<br />
Figura 1. Le caratteristiche dei Briozoi.<br />
In alto: espanso o retratto che sia, il lofoforo rappresenta la<br />
struttura predominante nell’anatomia dei Briozoi. I Briozoi,<br />
spesso agitano il loro lofoforo per aumentare la sua efficienza.<br />
In basso: particolare del lofoforo.
Il singolo componente della colonia viene definito polipide<br />
o zooide, mentre la colonia stessa è detta zooario.<br />
Dotati di notevoli specializzazioni legate alla loro condizione<br />
coloniale, i briozoi rappresentano, tra i lofoforati, l'unico<br />
esempio di organismi in grado di protrarre, di ruotare e<br />
esterno, dove avviene la fecondazione. Le larve dei<br />
brachiopodi rappresentano una componente dello<br />
zooplancton, ma la fase larvale è breve, poiché dopo pochi<br />
giorni l'organismo si fa sedentario e compie la<br />
metamorfosi, trasformandosi in adulto.<br />
di retrarre la struttura tentacolata periorale; in questo<br />
modo risulta potenziata la funzione di raccolta del cibo, che<br />
altrimenti richiederebbe l'afflusso di notevoli quantità di<br />
acqua verso il lofoforo.<br />
Lo zooario si forma per opera di un esemplare fondatore,<br />
che si riproduce in modo asessuale; la struttura sociale che<br />
ne deriva può contenere un numero variabile di organismi a<br />
seconda della specie, arrivando sino a un massimo di 2<br />
milioni di individui.<br />
In alcuni casi la vita coloniale comporta la presenza di<br />
organismi specializzati, ognuno con una funzione diversa:<br />
la nutrizione, la riproduzione sessuale (che si affianca a<br />
quella asessuale) o il sostegno. Gli spermatozoi vengono<br />
rilasciati nell'ambiente esterno, e da questo sono<br />
successivamente raccolti per portare a termine la<br />
fecondazione che è interna; gli individui si liberano solo in<br />
fase larvale, allorché si muovono attivamente alla ricerca di<br />
un substrato adatto al loro ancoraggio, sul quale si<br />
trasformeranno in adulti.<br />
I Brachiopodi sono simili ai bivalvi.<br />
Figura 2. Le caratteristiche dei Brachiopodi.<br />
(A) Lingula anatina è una specie bentonica che vive infossata<br />
nella sabbia grazie all’azione di un peduncolo muscolare e che<br />
si alimenta con un lofoforo. (B) Le due valve della conchiglia dei<br />
Brachiopodi sono una dorsale e una ventrale e non una destra e<br />
una sinistra come nei molluschi bivalvi.<br />
II phylum comprende organismi marini solitari che<br />
superficialmente ricordano i molluschi dei bivalvi,<br />
presentando una porzione mantellare e una conchiglia a<br />
due valve (▶figura 2); queste si muovono però secondo<br />
piani dorso-ventrali e non latero-laterali. Il loro lofoforo si<br />
presenta organizzato in due rami e risulta protetto dalla<br />
conchiglia. L'acqua carica dei microscopici organismi da<br />
filtrare viene richiamata all'interno della camera delimitata<br />
dalle due valve socchiuse. Si tratta di animali fissi,<br />
ancorati a un substrato o immersi in sedimenti di<br />
consistenza molle; talvolta si fissano tramite un peduncolo<br />
flessibile che consente loro di elevarsi rispetto al substrato<br />
stesso. Gli scambi gassosi avvengono attraverso porzioni<br />
non specializzate della superficie corporea o in<br />
corrispondenza dei tentacoli del lofoforo. La maggior<br />
Numerosissimi nel Paleozoico (o Primario) e nel Mesozoico<br />
(o Secondario), come testimoniano 12000 specie fossili, i<br />
brachiopodi sono attualmente rappresentati da un numero<br />
non elevato di specie (circa 350) che, comunque, sono<br />
abituali in numerosi ambienti marini. Si ritiene, a livello di<br />
ipotesi, che il loro declino a partire dal Mesozoico sia<br />
dovuto alla competizione con i molluschi, rispetto ai quali<br />
condividono le medesime fonti alimentari.<br />
Le parole:<br />
Briozoi deriva dal greco brýon, «muschio», e zoion,<br />
«animale».<br />
Brachiopodi deriva dal greco brakhýs «corto», e póuspodós,<br />
«piede».<br />
parte dei brachiopodi libera i propri gameti nell'ambiente
2. I deuterostomi condividono molte caratteristiche embriologiche e<br />
strutturali.<br />
Saranno ora passati in rassegna gli organismi animali appartenenti alla linea evolutiva dei deuterostomi che, come abbiamo<br />
visto precedentemente, presentano caratteristiche di sviluppo diverse, se non alternative, rispetto ai protostomi. Nel corso di<br />
questa trattazione verranno messe in risalto le fasi critiche della filogenesi dei vari gruppi appartenenti a questa fondamentale<br />
categoria sistematica. Tali punti cruciali dell'evoluzione dei deuterostomi saranno correlati ai cambiamenti delle strategie<br />
alimentari e a quelli di tipo ecologico (cioè relativi all'habitat). In rapporto ai meccanismi di alimentazione, verranno trattati in<br />
modo dettagliato i processi evolutivi che hanno coinvolto gli apparati specializzati per l'assunzione del cibo.<br />
I deuterostomi.<br />
Tra tutti i gruppi riferibili ai deuterostomi verranno<br />
descritti il phylum degli Echinodermi (▶figura 3) e quello<br />
dei Cordati (▶figura 4), una categoria sistematica di<br />
grandissimo rilievo per l'ampio spettro adattativo che<br />
la caratterizza e per il fatto di comprendere la nostra<br />
Alcune specie possiedono un corpo segmentato,<br />
sebbene i singoli segmenti siano meno evidenti rispetto<br />
a quanto si osserva negli anellidi e negli artropodi.<br />
I due phyla più importanti della linea evolutiva dei<br />
deuterostomi sono gli Echinodermi e i Cordati.<br />
specie. I caratteri embriologici essenziali dei<br />
deuterostomi sono i seguenti:<br />
<br />
<br />
lo sviluppo dell'uovo è di tipo sostanzialmente<br />
regolativo;<br />
il blastoporo da origine all’ano mentre la bocca si<br />
forma all’estremità opposta rispetto al blastoporo;<br />
il celoma si forma a partire da tasche<br />
di mesoderma che si separano dalla cavità<br />
della gastrula (enterocelia), invece che per scissione<br />
del mesoderma (schizocelia), come avviene nei<br />
protostomi.<br />
Gli stretti rapporti evolutivi tra echinodermi e cordati (i<br />
due phyla più importanti dei deuterostomi) sono<br />
accertati anche dalle analisi genetiche; infatti le<br />
sequenze del DNA di molti geni indicano una loro<br />
Figura 3. Riccio di mare: un Echinoderma.<br />
Gli Echinodermi sono animali acquatici, esclusivamente marini<br />
dotati di uno scheletro dermico di materiale calcareo. Da adulti<br />
presentano simmetria radiale ma questa condizione può essere<br />
considerata secondaria in quanto le larve di questi animali<br />
presentano simmetria bilaterale.<br />
comune discendenza.<br />
Difficilmente riconducigli a un progenitore comune<br />
(i primi fossili sono già riferibili alle principali linee del<br />
gruppo), i deuterostomi rivelano un piano<br />
organizzativo fondamentalmente simile, basato<br />
sulla presenza di tre foglietti embrionali e di cavità<br />
celomatiche ben definite.<br />
Per quanto non così ricchi in numero di specie come i<br />
protostomi, i deuterostomi rivelano una maggiore<br />
varietà di piani strutturali. Tutti i deuterostomi sono<br />
celomati, triblastici e provvisti di endoscheletro.<br />
Figura 4. Rana: un Cordato.<br />
I Cordati si sono diversificati notevolmente e tra essi, i<br />
Vertebrati sono quelli che hanno colonizzato praticamente tutti<br />
gli ambienti del pianeta.
3. Gli Echinodermi: deuterostomi a simmetria radiale con rivestimento<br />
spinoso e scheletro interno.<br />
Di regola la simmetria bilaterale è associata ad abitudini di vita che prevedono attivi movimenti, mentre la simmetria raggiata è tipica di<br />
organismi sessili o, al più, mediocremente mobili. Tra i deuterostomi è comunque presente un numero notevole di organismi di grande<br />
successo evolutivo, che combinano la simmetria raggiata con peculiari adattamenti motori.<br />
I caratteri distintivi degli<br />
Echinodermi.<br />
Questo phylum comprende organismi molto complessi che<br />
hanno messo a punto un elaborato scheletro superficiale<br />
dermico (dermascheletro) a base di sali di calcio<br />
(prevalentemente calcarei), rivestito da tessuti molli:<br />
tegumento e muscoli. Un'ulteriore, notevole<br />
specializzazione è rappresentata dall'idrocele, un derivato<br />
del celoma (per lo più del mesocele), che consente loro di<br />
modificare la forma del corpo e di compiere movimenti.<br />
L'idrocele comprende una serie di canali che dispongono di<br />
estensioni terminali definite pedicelli ambulacrali, devolute<br />
alla locomozione, all'alimentazione e agli scambi<br />
respiratori. L'acqua marina penetra nell'idrocele tramite un<br />
sistema di pori in corrispondenza della placca madreporica,<br />
L'acqua dell'idrocele è mantenuta in movimento, grazie alla<br />
spinta di strutture muscolari specializzate, in particolare di<br />
quelle associate ai pedicelli.<br />
Le cinque classi degli Echinodermi.<br />
La documentazione fossile sugli echinodermi è notevole: se<br />
ne conoscono 17 classi estinte e 5 tuttora esistenti; 6000<br />
sono le specie attualmente viventi alle quali se ne devono<br />
aggiungere altre 13 000 fossili, per altro rappresentative<br />
solo di una parte di quelle scomparse.<br />
Quasi tutte le specie prevedono una fase larvale<br />
caratterizzata da un individuo ciliato a simmetria bilaterale<br />
(▶figura 6), il quale vive nello zooplancton prima di<br />
acquisire abitudini sedentarie e metamorfosare,<br />
trasformandosi nell'adulto a simmetria raggiata.<br />
che in alcune specie è in connessione con un canale con<br />
decorso anulare che circonda l'esofago. Da questo canale<br />
anulare si dipartono altri canali, definiti radiali, uno per<br />
ognuno delle braccia ambulacrali. Con questo termine si<br />
indicano le caratteristiche appendici che, in alcuni gruppi di<br />
echinodermi, garantiscono i movimenti sul substrato con<br />
il concorso dei pedicelli connessi ai canali radiali (▶figura<br />
5).<br />
Figura 6. La larva degli Echinodermi è dotata di simmetria<br />
bilaterale.<br />
Questa curiosa “candela” è una larva di Luidia ciliaris, una delle<br />
tante specie di stelle marine.<br />
Il phylum degli Echinodermi viene suddiviso in due subphyla:<br />
i pelmatozoi e gli eleuterozoi. I due sub-phyla sono<br />
distinguibili in base alla struttura dell'idrocele e al numero<br />
delle appendici del corpo.<br />
I pelmatozoi comprendono la classe dei Crinoidei, gli<br />
Figura 5. Le caratteristiche degli echinodermi.<br />
Una visione dorsale della stella marina illustra lo scheletro<br />
interno calcificato e il sistema vascolare acquifero composto dai<br />
canali acquiferi e dai pedicelli ambulacrali.<br />
eleuterozoi comprendono le restanti classi: Ofiuroidei,<br />
Asteroidei, Echinoidei ed Oloturoidei.
I crinoidei: i gigli di mare.<br />
Nei Crinoidei, l’unica classe di pelmatozoi, il lofoforo ha<br />
subito, al pari delle altre parti del corpo, un processo di<br />
calcificazione.<br />
Attualmente ne sopravvivono solo due gruppi: i<br />
pentacrinidi e gli antedonidi, per non più di 80 specie.<br />
I pentacrinidi (▶figura 7) vengono definiti gigli di mare<br />
per la loro forma; vivono ancorati al substrato tramite un<br />
peduncolo che è flessibile, in quanto il suo scheletro<br />
calcareo è formato da dischi.<br />
Al peduncolo segue la porzione espansa del corpo, simile a<br />
una coppa, dalla quale si dipartono da cinque a centinaia di<br />
braccia (di regola il numero è comunque multiplo di<br />
cinque). Le braccia presentano uno scheletro articolato e<br />
dunque possono piegarsi. Ogni braccio è percorso da un<br />
solco che porta sui due lati una serie di pedicelli corredati<br />
Figura 8. Gli antedonidi sono simili ai gigli di mare.<br />
Antedon mediterranea, una specie molto diffusa nel<br />
mediterraneo.<br />
Gli ofiuroidei: le stelle serpentine.<br />
Gli ofiuroidei comprendono le cosiddette stelle serpentine<br />
(▶figura 9) che ricordano i crinoidei liberi per la presenza<br />
di appendici flessibili, poiché le placchette calcaree<br />
dell'endoscheletro risultano reciprocamente articolate.<br />
da elementi cellulari secernenti muco.<br />
Il meccanismo di acquisizione del cibo prevede la cattura di<br />
particelle alimentari o di minuti organismi tramite i<br />
pedicelli; il cibo è quindi trasferito al solco e di qui alla<br />
cavità orale grazie al movimento di un epitelio ciliato. I<br />
pedicelli servono anche agli scambi gassosi e all'escrezione.<br />
Figura 7. I Pentacrinidi sono definiti gigli di mare.<br />
I Pentacrinidi vengono spesso definiti “gigli di mare”.<br />
Gli antedonidi (▶figura 8) sono simili ai gigli di mare, a<br />
parte l'assenza del peduncolo e la presenza di braccia<br />
flessibili e piumose. Essi utilizzano le loro braccia per<br />
nutrirsi, ma anche per ancorarsi al substrato, per<br />
spostarsi su questo e per nuotare.<br />
Figura 9. Gli Ofiuroidei vengono definiti stelle serpentine.<br />
In alto, Ophiura ophiura presenta cinque braccia allungate ed è<br />
per questo che gli Ofiuroidei vengono definiti “stelle<br />
serpentine”. In basso, una stella serpentina nelle mani di un<br />
ricercatore viene confrontata con una stella marina che si vede<br />
sullo sfondo.<br />
Durante il movimento vengono agitate le braccia, che sono<br />
sempre cinque anche se provviste di rami, e in certi casi
anche i pedicelli sono coinvolti nel movimento. Come<br />
caratteristica anatomica peculiare, gli ofiuroidei presentano<br />
una sola apertura del canale digerente, attraverso la quale<br />
essi assumono il cibo, particelle del sedimento presenti in<br />
braccia e spingendo nel senso dell'apertura le due valve,<br />
gli asteroidei possono alla lunga rendere i muscoli del<br />
mollusco totalmente esausti e scardinare dunque la pur<br />
ermetica chiusura della conchiglia.<br />
sospensione e piccoli organismi animali, oppure eliminano<br />
i residui dei processi alimentari.<br />
Oltre a ciò i pedicelli funzionano come organi locomotori e<br />
di scambio respiratorio. Nel corso della predazione sui<br />
Gli asteroidei: le stelle marine.<br />
bivalvi le stelle di mare sono in grado di estroflettere la<br />
porzione gastrica del loro tratto digerente dall'apparato<br />
orale, e di introdurla nella cavità della conchiglia attraverso<br />
Gli asteroidei comprendono le comuni stelle di mare<br />
(▶figura 10). Simili agli ofiuroidei, ma provvisti di<br />
appendici meno flessibili, le stelle marine presentano per<br />
lo più abitudini predatorie anche nei confronti dei<br />
molluschi e dei pesci (esse causano spesso danni ingenti<br />
agli allevamenti di ostriche). I loro pedicelli rappresentano<br />
efficienti organi adesivi, grazie a un meccanismo che<br />
funziona come una ventosa a depressione, reso più<br />
efficiente dalla presenza di secreti adesivi (▶figura 10).<br />
le due valve divaricate. In questo modo vengono emessi<br />
all'esterno gli enzimi digestivi che agiscono sui tessuti<br />
molli indifesi del mollusco. Altri asteroidei, che si nutrono<br />
di prede minute, praticano una digestione più tradizionale,<br />
mantenendo lo stomaco all'interno del corpo.<br />
Gli echinoidei: i ricci di mare.<br />
Gli echinoidei comprendono i ricci di mare (▶figura 11)<br />
sono sprovvisti di braccia e sono ben noti per la loro forma<br />
emisferica e per le caratteristiche spine che risultano<br />
applicate allo scheletro sottostante, formato da placche<br />
reciprocamente fuse. Il rapporto tra spine e scheletro<br />
dermico è regolato da un'articolazione analoga all'enartrosi<br />
dell'anatomia umana. Gli echinoidei si nutrono di alghe o di<br />
minuti detriti alimentari e presentano una tipica struttura<br />
raschiante che essi utilizzano per alimentarsi “brucando” le<br />
alghe del fondale: la lanterna di Aristotele (▶figura 12). Molti<br />
autori rinvengono omologie tra le stelle marine e i ricci di<br />
mare, i quali corrisponderebbero ad asteroidei con le<br />
braccia ripiegate e fuse sul dorso.<br />
Figura 10. Gli Asteroidei vengono definiti stelle marine.<br />
In alto, una stella marina tropicale che presenta delle sculture e<br />
una colorazione vivace. In basso, si vedono molto bene i<br />
pedicelli ambulacrali con i quali le stelle marine riescono a<br />
spostarsi e ad alimentarsi.<br />
La possibilità di usare simultaneamente centinaia di<br />
pedicelli a ventosa rende le stelle marine predatori<br />
potentissimi: ancorandosi a un bivalve con le proprie<br />
Figura 11. Gli Echinoidei vengono definiti ricci di mare.<br />
Un riccio di mare dall’aspetto globoso e con le tipiche spine che<br />
caratterizzano questa classe.
Privi di spine, dispongono di serie di pedicelli disposte su<br />
ambo i lati di cinque solchi che si sviluppano<br />
longitudinalmente sul corpo; i pedicelli servono per<br />
garantire adesione e ancoraggio al substrato piuttosto che<br />
per il movimento, e in certe specie possono risultare del<br />
tutto assenti. Quando presenti, i pedicelli appaiono<br />
modificati in corrispondenza dell'estremità orale, dove<br />
formano voluminosi tentacoli finemente ramificati e<br />
provvisti di una sostanza adesiva deposta superficialmente.<br />
Questi tentacoli vengono periodicamente estroflessi e<br />
Figura 12. La lanterna di Aristotele.<br />
E’ la tipica struttura dell’apparato raschiatore dei ricci di mare<br />
che permette loro di brucare il substrato. La lanterna é<br />
composta da cinque placche calcaree, dette piramidi, unite da<br />
fibre muscolari trasverse. Lungo la faccia interna di ogni<br />
piramide scorre un lungo dente dall'estremità appuntita.<br />
introflessi dalla bocca: nell'ambiente esterno catturano<br />
minuti organismi e detriti che vengono poi digeriti e<br />
assimilati, una volta che i tentacoli vengono reintrodotti<br />
nella bocca.<br />
Alcune specie, quando vengono disturbate, possono<br />
Gli oloturoidei: i cetrioli di mare.<br />
Gli oloturoidei comprendono i cetrioli di mare (▶figura 13)<br />
anch’essi sprovvisti di braccia. Essi possono essere considerati<br />
emettere lunghi filamenti vischiosi e, al tempo stesso,<br />
espellere per autotomia, parti dei loro organi interni<br />
(polmoni acquiferi), dall’orifizio cloacale al fine di<br />
disorientare il loro aggressore. Esse, in attesa di rigenerare<br />
le parti perdute, respirano tramite la cavità corporea.<br />
ricci di mare dal corpo flessibile (per la riduzione del proprio<br />
scheletro) e allungato.<br />
Le parole:<br />
Echinoderma deriva dal greco echínos «riccio», e derma<br />
«pelle» e fa riferimento alla presenza di placche<br />
scheletriche nella pelle.<br />
Nei termini che seguono, la desinenza -idei deriva dal<br />
greco eidés «simile a».<br />
Crinoideo deriva dal greco krínon «giglio ».<br />
Ofiuoroideo deriva dal greco óphis «serpente», e ourá,<br />
«coda».<br />
Asteroidei deriva dal greco astér «stella».<br />
Echinoidei deriva dal greco echínos «riccio».<br />
Oloturoidei<br />
mare».<br />
deriva dal greco olothurion «cetriolo di<br />
Figura 13. Gli Oloturoidei vengono definiti cetrioli di mare.<br />
In alto, un cetriolo di mare dall’aspetto allungato mostra un<br />
aspetto quasi cilindrico. In basso, un’altra specie di oloturoideo<br />
dall’aspetto ancora più allungato e quasi vermiforme.
4. L’evoluzione della faringe: i Cordati.<br />
Un'altra linea evolutiva dei deuterostomi, per utilizzare le abbondanti risorse alimentari del plancton marino, ha messo a<br />
punto un complesso dispositivo fessurato in corrispondenza della faringe, in grado, come i lofofori, di raccogliere per<br />
filtrazione le minute prede planctoniche. In un secondo tempo queste fessure faringee, definite anche branchiali, hanno<br />
assunto il significato di dispositivi per lo scambio respiratorio, probabilmente attraverso una fase transitoria nella quale le<br />
funzioni di scambio gassoso e quelle di filtraggio coesistevano. La trasformazione dell'apparato branchiale filtrante nel<br />
corso della filogenesi, costituisce un processo evolutivo di grandissimo rilievo che caratterizza un gruppo peculiare di<br />
deuterostomi: i cordati.<br />
Le caratteristiche dei Cordati.<br />
I cordati presentano simmetria bilaterale e, in<br />
sostituzione dei lofofori, possiedono una regione<br />
faringea (o branchiale) estesamente fessurata, o<br />
almeno, caratterizzata da abbozzi di fessure in stadi<br />
Il phylum dei Cordati si divide in tre<br />
sub-phyla.<br />
La classificazione dei Cordati tiene conto della<br />
localizzazione della corda e della sua permanenza o<br />
meno nell’adulto. I sub-phyla sono:<br />
precoci di sviluppo. Il piano organizzativo dei cordati<br />
comprende un endoscheletro assile in posizione<br />
SUB-PHYLUM Quando è<br />
presente:<br />
Dove<br />
localizzata:<br />
è<br />
dorsale, interposto tra due apparati parimenti assili<br />
che gli sono adiacenti e paralleli: ventralmente il<br />
canale alimentare e dorsalmente il sistema nervoso.<br />
Urocordati temporanea o coda<br />
permanente<br />
Cefalocordati permanente in tutto il<br />
corpo<br />
Per quanto l'endoscheletro definitivo nei cordati<br />
adulti possa variare notevolmente in consistenza e<br />
struttura, tutti possiedono, in qualche fase di<br />
sviluppo, un esclusivo organo di sostegno definito<br />
corda dorsale, (notocorda). Essa è formata da un asse<br />
di cellule con vacuoli colmi di liquido, che rendono l’intera<br />
struttura rigida ma flessibile. Questa struttura può<br />
avere estensioni diverse e può essere permanente o<br />
transitoria a seconda dei sub-phyla. In diversi casi,<br />
Vertebrati quasi sempre<br />
Le parole:<br />
temporanea<br />
tronco e coda<br />
negli adulti, essa viene rimpiazzata da formazioni<br />
scheletriche più consistenti.<br />
Riassumendo, i caratteri peculiari dei cordati sono:<br />
una struttura dorsale di sostegno definita notocorda;<br />
delle fessure branchiali situate a livello della<br />
faringe, la porzione del tubo digerente posta subito<br />
dopo la bocca.<br />
un tubo neurale dorsale;<br />
La notocorda delle larve degli urocordati non arriva fino<br />
all’estremità anteriore del corpo; da questo deriva il<br />
nome del gruppo (in greco ourá significa «coda»).<br />
Il contrario di ciò che accade per icefalocordati (kephalé,<br />
«testa»).<br />
Anfiosso, dal greco amphí, «da due parti», e oxýs,<br />
«appuntito», significa «animale con due punte».<br />
In italiano si chiama anche (impropriamente!) «pesce<br />
lanceolato» e in inglese lancelet, da lancet, bisturi. Tutti<br />
i termini si rifanno alla forma caratteristica del corpo di<br />
questo animale.<br />
<br />
una coda che si estende oltre l’ano;
Gli urocordati comprendono le<br />
ascidie, le salpe e le appendicolarie.<br />
Il movimento dell'acqua attraverso le numerose<br />
fessure branchiali è assicurato dall'epitelio ciliato<br />
della faringe, internamente rivestita da muco, il quale<br />
I tunicati, appartenenti al sub-phylum urocordati,<br />
rappresentano cordati ancestrali esclusivamente<br />
marini e spesso, da adulti, sono adattati a vivere<br />
ancorati a un substrato. In alcuni casi è presente una<br />
distinta fase larvale mobile che possiede le<br />
caratteristiche anatomiche del tipo: una regione<br />
branchiale filtrante, un sistema nervoso assile e, nella<br />
sola regione della coda, una corda dorsale, sulla quale<br />
si inseriscono i muscoli responsabili del movimento<br />
serve a intrappolare le particelle alimentari. L'acqua<br />
penetra nella faringe tramite un sifone orale, passa<br />
attraverso il cestello per raggiungere uno spazio<br />
definito atrio, compreso tra il cestello stesso e la<br />
tunica, e viene quindi esalata attraverso un sifone<br />
atriale. Alcune ascidie formano colonie mediante<br />
gemmazione (riproduzione asessuata) a partire da un<br />
individuo fondatore; queste colonie possono<br />
raggiungere un diametro di diversi metri.<br />
(▶figura 14A). Questi muscoli vengono definiti<br />
segmentali poiché hanno una struttura modulare<br />
formando una serie in senso cefalo-caudale, e poiché<br />
derivano da mesoderma segmentato. Dopo una breve<br />
fase di vita, caratterizzata da attivi movimenti, la larva<br />
si trasforma in un adulto sedentario. Lo stato di<br />
immobilità viene compensato dall'ampliamento<br />
dell'apparato filtrante, divenuto simile a un cestello<br />
estesamente fessurato.<br />
Più del 90% delle specie note di tunicati appartengono<br />
alla classe degli ascidiacei i cui adulti, solitari o<br />
coloniali, presentano un corpo sacciforme rivestito da<br />
un involucro (tunica) di natura proteica e glucidica<br />
Un secondo gruppo di tunicati comprende le salpe che<br />
possono vivere singolarmente oppure formare colonie<br />
simili a catenelle, che raggiungono diversi metri di<br />
lunghezza (▶figura 14C). Questi animali fluttuano negli<br />
oceani tropicali e subtropicali fino a 1500 m di<br />
profondità.<br />
In un terzo gruppo di tunicati, i larvacei o<br />
appendicolarie vengono mantenute costantemente<br />
aspetto e abitudini larvali (neotenia); queste forme,<br />
riconducibili a specie poco numerose ma molto<br />
diffuse, raggiungono la maturità sessuale in fase<br />
larvale (pedogenesi).<br />
(▶figura 14B).<br />
Figura 14. Gli urocordati.<br />
(A) Uno schema della larva di un’ascidia, in cui è ben visibile la notocorda. (B) L’aspetto iridescente della tunica è evidente in queste ascidie<br />
trasparenti. Nella fotografia sono illustrate due diverse specie appartenenti a uno stesso genere. (C) Una colonia di salpe che galleggia nelle acque<br />
tropicali.
I cefalocordati sono filtratori a vita<br />
libera.<br />
Per la maggior parte del tempo, questi animali si<br />
trovano infossati nella sabbia, con il capo sporgente dal<br />
sedimento, sebbene in realtà essi siano anche capaci di<br />
Le trenta specie di anfiossi sono piccoli animali che<br />
nuotare.<br />
raramente superano la lunghezza di 5 cm. La notocorda<br />
si estende per l’intera lunghezza del corpo e persiste<br />
per tutta la vita (▶figura 15). Gli anfiossi vivono nelle<br />
acque salate e salmastre poco profonde di tutto il<br />
mondo.<br />
Gli anfiossi estraggono le prede dall’acqua grazie ai<br />
cestelli branchiali. Durante la stagione riproduttiva<br />
entrambi i sessi liberano uova e spermatozoi nell’acqua,<br />
dove ha luogo la fecondazione.<br />
Figura 15. I cefalocordati.<br />
Questo animale, chiamato anfiosso, è un tipico esempio di cefalocordato.<br />
I vertebrati e le loro caratteristiche.<br />
I vertebrati presentano un più efficiente dispositivo di<br />
sostegno del tronco rispetto alla corda: una colonna<br />
Un'estesa fessurazione faringea rappresenta un<br />
adattamento assai efficiente per raccogliere, tramite<br />
filtrazione, le particelle alimentari e i piccoli organismi<br />
contenuti in sospensione nell'acqua. Le numerose<br />
aperture del cestello branchiale consentono infatti<br />
l'eliminazione dell'acqua e delle particelle non utilizzabili<br />
ingerite dalla bocca. Nel tardo Cambriano, oltre<br />
500 milioni di anni fa, i primi cordati hanno messo a<br />
punto tale meccanismo piuttosto innovativo per<br />
ricavare cibo dal fondo fangoso o sabbioso. I loro<br />
discendenti noti come vertebrati hanno mantenuto<br />
inizialmente tali abitudini alimentari utilizzando un<br />
apparato branchiale filtrante, nel quale le singole<br />
fessure risultavano sorrette da specifici sostegni<br />
scheletrici (archi).<br />
vertebrale, costituita da elementi scheletrici tra loro<br />
articolati.<br />
I vertebrati costituiscono un gruppo di cordati che<br />
devono il proprio nome alla colonna vertebrale, una<br />
nuova struttura dorsale di supporto che sostituisce la<br />
notocorda durante le prime fasi di sviluppo embrionale.<br />
Essi comprendono due grandi suddivisioni:<br />
gli agnati (privi di mandibole e di mascelle) e<br />
gli gnatostomi (i pesci cartilaginei, i pesci ossei, gli<br />
anfibi, i rettili, gli uccelli e i mammiferi).<br />
Prima di prendere in considerazione le varie classi di<br />
Vertebrati, passiamo in rassegna i principali sistemi ed<br />
apparati che si riscontrano nei vertebrati facendo un<br />
breve cenno sulle loro funzioni generali.
Il sistema tegumentario è deputato alla protezione del<br />
corpo può presentare vari tipi di strutture come scaglie<br />
ossee, squame cornee, unghie, penne e peli. Nelle<br />
specie che hanno un sistema respiratorio ridotto, è<br />
formato da pelle nuda che consente gli scambi gassosi.<br />
o quattro cavità, definite atri (se accolgono il sangue<br />
proveniente dalla periferia) o ventricoli (se inviano il<br />
sangue verso la periferia). I vasi sanguigni che<br />
trasportano il sangue dal cuore alla periferia sono detti<br />
arterie, esse si ramificano in arterie a calibro via via<br />
inferiore fino a che si risolvono nei capillari a livello dei<br />
Lo scheletro dei vertebrati può essere cartilagineo<br />
oppure osseo, ma fondamentalmente, esso presenta lo<br />
stesso modello di base (▶figura 16). E’ suddiviso in<br />
scheletro assile (presente lungo l’asse del corpo) e<br />
quali avvengono gli scambi tra il sangue e le cellule. I<br />
capillari confluiscono nelle vene che riportano il sangue<br />
al cuore (▶figura 17).<br />
scheletro appendicolare (che costituisce le appendici).<br />
Lo scheletro assile è formato dal cranio che protegge<br />
l’encefalo e dalla colonna vertebrale che protegge il<br />
midollo spinale. Le appendici possono essere<br />
rappresentate da pinne nelle forme acquatiche o da arti<br />
nelle forme terrestri e in qualche caso, si può assistere<br />
alla perdita, parziale o totale degli arti. Le ossa o i<br />
segmenti cartilaginei che costituiscono lo scheletro<br />
sono articolati tra loro tramite articolazioni più o meno<br />
mobili. Sullo scheletro si inseriscono i muscoli che<br />
tramite la contrazione, permettono il movimento. Lo<br />
scheletro, unitamente alle articolazioni e ai muscoli<br />
scheletrici forma il sistema locomotore.<br />
Figura 17. Il sistema circolatorio dei Vertebrati.<br />
Il sistema circolatorio dei Vertebrati è chiuso e il sangue scorre<br />
sempre all’interno dei vasi sanguigni.<br />
L’apparato respiratorio, sempre strettamente connesso<br />
al sistema circolatorio, è rappresentato da branchie<br />
nelle forme acquatiche e da polmoni, nelle forme<br />
terrestri. In alcuni casi, la pelle rappresenta un organo<br />
con funzione respiratoria che coadiuva o addirittura<br />
sostituisce gli organi respiratori veri e propri.<br />
L’apparato digerente è dotato di bocca posta<br />
sull’estremità cefalica e di ano, che si trova in<br />
Figura 16. L’endoscheletro dei Vertebrati.<br />
Lo scheletro dei Vertebrati si suddivide in scheletro assile e<br />
scheletro appendicolare.<br />
Il sistema circolatorio dei Vertebrati è di tipo chiuso<br />
perché il sangue non esce mai dai vasi sanguigni e si<br />
possono distinguere diversi tipi di circolazione. La<br />
distinzione principale è basata sul fatto che il sangue<br />
possa passare una sola volta o due volte dal cuore<br />
durante il suo ciclo. Nel primo caso, la circolazione è<br />
detta semplice, mentre nel secondo caso è detta<br />
doppia. Un’altra distinzione importante tiene conto del<br />
fatto che il sangue ossigenato si mescoli anche<br />
parzialmente con il sangue povero di ossigeno e ricco<br />
di anidride carbonica, oppure no. In questi due casi si<br />
parla di circolazione incompleta e completa,<br />
rispettivamente. Il cuore rappresenta la pompa del<br />
sistema circolatorio e può essere costituito da due, tre<br />
prossimità della coda (ma non proprio alla sua<br />
estremità). Il tubo digerente è formato da organi cavi<br />
all’interno dei quali avvengono le varie fasi della<br />
digestione ed è posizionato sempre ventralmente<br />
rispetto alla corda. Al tubo digerente sono annesse<br />
ghiandole che producono diversi enzimi digestivi. Nella<br />
bocca possono essere presenti, squame cornee o veri e<br />
propri denti che vengono utilizzati inizialmente solo<br />
per trattenere il cibo oppure per la masticazione.<br />
Quando i denti sono tutti uguali tra loro come forma, si<br />
ha una condizione detta omodonzia, nel caso in cui i<br />
denti siano diversificati, si parla di eterodonzia.<br />
L’assenza di denti è detta anodonzia.<br />
Il sistema immunitario consente ai vertebrati di<br />
difendersi dalle aggressione dei microrganismi<br />
patogeni tramite la produzione di cellule immunitarie e<br />
di molecole proteiche, le immunoglobuline (anticorpi).
Il sistema nervoso dei Vertebrati è suddiviso in centrale<br />
(SNC) e periferico (SNP). Il primo è costituito<br />
dall’encefalo (protetto all’interno della scatola cranica)<br />
e dal midollo spinale (protetto all’interno della colonna<br />
vertebrale). Il secondo è costituito dai nervi che<br />
collegano il SNC alla periferia. Il sistema nervoso dei<br />
Vertebrati si forma a partire dal tubo neurale che a sua<br />
volta si forma dorsalmente rispetto alla corda. Durante<br />
Nei vertebrati possono essere presenti vari tipi di<br />
recettori spesso riuniti per formare veri e propri organi<br />
di senso. In base al tipo di stimoli che vengono<br />
percepiti si distinguono: fotorecettori (sensibili alla<br />
luce), chemiorecettori (sensibili a stimolazioni<br />
chimiche), meccanorecettori (sensibili a stimolazioni<br />
meccaniche), termorecettori (sensibili a stimolazioni<br />
termiche), etc..<br />
lo sviluppo embrionale, la porzione di tubo neurale che<br />
si sviluppa nel capo, si suddivide prima in tre ed poi in<br />
cinque vescicole encefaliche. Allo stadio di tre<br />
vescicole, si distinguono in senso cefalo-caudale:<br />
prosencefalo, mesencefalo e romboencefalo. In seguito<br />
dal prosencefalo si formano telencefalo e diencefalo, il<br />
mesencefalo non si suddivide e dal romboencefalo si<br />
sviluppano metaencefalo e mielencefalo (▶figura 18).<br />
Il sistema endocrino è un sistema di controllo costituito<br />
dalle ghiandole endocrine che producono ormoni. Tali<br />
molecole, di natura lipidica o proteica, regolano<br />
importanti funzioni come l’accrescimento, la<br />
riproduzione e l’omeostasi. Esso è strettamente<br />
collegato funzionalmente al sistema nervoso.<br />
L’ apparato escretore è costituito dai reni che filtrano il<br />
sangue e eliminano nell’urina<br />
le scorie metaboliche<br />
prodotte dal metabolismo cellulare. Le scorie azotate<br />
prodotte dal metabolismo delle proteine e degli acidi<br />
nucleici possono essere eliminate come ammoniaca,<br />
urea o acido urico e gli animali possono essere definiti<br />
rispettivamente ammoniotelici, ureotelici ed uricotelici.<br />
L’ammoniaca è prodotta dai vertebrati che vivono in un<br />
ambiente acquatico perché è molto tossica e quindi<br />
deve essere eliminata con un grande dispendio idrico. I<br />
Vertebrati terrestri eliminano principalmente urea<br />
(meno tossica dell’ammoniaca) o acido urico (che<br />
cristallizza e può permettere di effettuare un grande<br />
risparmio idrico). Nel corso dell’evoluzione dei<br />
Vertebrati, si sono evoluti tre tipi diversi di reni che,<br />
nell’ordine di comparsa sono: il prorene, il mesorene<br />
ed il metarene. Quest’ordine di comparsa non riguarda<br />
solo l’evoluzione, ma si ripete anche nello sviluppo di<br />
ogni singolo individuo: nei vertebrati primitivi compare<br />
solo il prorene e tale organo funziona come organo<br />
escretore nell’adulto, in altri vertebrati, invece,<br />
esso<br />
funziona solo durante la vita embrionale mentre<br />
nell’adulto viene sostituito dal mesorene. In vertebrati<br />
ancora più evoluti, anche il mesorene viene sostituito<br />
dal metarene.<br />
La riproduzione dei vertebrati è quasi sempre sessuata<br />
e i due sessi sono separati (il maschio produce gameti<br />
maschili all’interno dei testicoli, mentre la femmina<br />
Figura 18. Lo sviluppo delle vescicole encefaliche.<br />
Lo sviluppo delle vescicole encefaliche nell’uomo.<br />
produce gameti femminili, all’interno delle ovaie).<br />
Spesso esiste dimorfismo sessuale.
I caratteri derivati dei cordati.<br />
una struttura dorsale di sostegno definita<br />
notocorda;<br />
In base a una combinazione di dati fossili, anatomici e<br />
molecolari, i biologi hanno formulato ipotesi<br />
sull’evoluzione del phylum dei cordati.<br />
La ▶figura 19 illustra l’idea più condivisa che inquadra i<br />
vari gruppi di cordati, assieme ad alcuni dei caratteri<br />
derivati che li contraddistinguono e la (▶figura 20)<br />
mostra l’evoluzione dei Vertebrati con uno schema<br />
cronologico.<br />
Riassumendo, sarebbero comparsi in sequenza, i<br />
seguenti caratteri:<br />
<br />
<br />
<br />
<br />
<br />
<br />
<br />
<br />
un encefalo<br />
un vero e proprio capo ed una colonna vertebrale<br />
una bocca dotata di mascelle e mandibole distinte<br />
i polmoni<br />
le pinne lobate<br />
gli arti<br />
l’uovo dotato di amnios<br />
la capacità di allattare i piccoli<br />
Figura 19. L’albero filogenetico dei cordati.<br />
Lo schema mette in evidenza i caratteri derivati condivisi.<br />
Figura 20. Filogenesi dei Vertebrati.<br />
Lo schema rappresenta l’evoluzione dei Vertebrati nelle varie ere geologiche.
5. I vertebrati acquatici.<br />
Le prime tre classi di vertebrati comprendono animali quasi esclusivamente acquatici comunemente denominati “pesci”. Va<br />
però chiarito che questo termine non ha un significato sistematico ben preciso in quanto comprende animali molto diversi<br />
tra loro e non necessariamente vicini dal punto di vista filogenetico.<br />
I Ciclostomi dovrebbero essere considerati separatamente dai pesci rispetto ai quali presentano importanti differenze: la<br />
bocca è priva di strutture articolate e la narice è impari e mediana (monorrini). Le altre due classi, invece comprendono i<br />
pesci propriamente detti, con arco orale suddiviso nell’arcata mascellare (superiore) e mandibolare (inferiore) e narici<br />
doppie (anfirrini). Essi si distinguono in Condroitti (pesci cartilaginei) e in Osteoitti (pesci ossei).<br />
Gli agnati presentano scheletro<br />
cartilagineo e arco orale non<br />
suddiviso.<br />
.<br />
I primi vertebrati, gli ostracodermi (▶figura 21),<br />
disponevano anche di uno scheletro esterno<br />
costituito da tavolati ossei che era in grado di<br />
proteggerli dagli attacchi dei loro predatori,<br />
artropodi giganteschi, e che viene ricordato nel<br />
nome stesso di questa classe, che letteralmente<br />
significa «animali dotati di scheletro cutaneo simile a<br />
una conchiglia». Gli ostracodermi rappresentano una<br />
classe della più ampia categoria (superclasse) degli<br />
agnati, definiti in tal modo per l'assenza di uno<br />
scheletro orale incernierato (si veda in seguito). II<br />
robusto esoscheletro consentiva ai piccoli<br />
ostracodermi (6-30 cm di lunghezza) di muoversi<br />
sul fondo piuttosto che scavare gallerie per sfuggire<br />
ai predatori; per quanto lenti nuotatori questi primi<br />
vertebrati erano in grado di esplorare il loro ambiente<br />
cercando substrati alimentari più ricchi e più adatti<br />
alla filtrazione. Il movimento autonomo<br />
nell'ambiente rappresenta, nel quadro<br />
comportamentale complessivo dei vertebrati, un<br />
elemento preso in considerazione dai paleontologi<br />
per ipotizzare la derivazione di questo gruppo da<br />
cordati ancestrali. Tra questi, le larve dei tunicati<br />
Figura 21. Ostracodermi.<br />
Ricostruzione di un Ostracodermo: Cephalaspis.<br />
I Ciclostomi: gli unici agnati attuali.<br />
Tornando ai vertebrati agnati, possiamo citare una<br />
classe attuale, i ciclostomi, comprendente un gruppo<br />
che, oltre a esplorare attivamente l'ambiente, ha<br />
escogitato un sistema più economico per muoversi e<br />
per nutrirsi. Le lamprede (petromizonti) (▶figura 22) si<br />
applicano con la loro bocca agnata, che funziona come<br />
una ventosa, al corpo di vertebrati più evoluti, si<br />
fanno da questi trasportare e li parassitano in un<br />
modo piuttosto cruento. Con una sorta di lingua<br />
dotata di dentelli cornei lacerano i tessuti superficiali<br />
dell'ospite (di solito un rappresentante dei pesci a<br />
scheletro osseo) e si nutrono del loro sangue e del<br />
liquido interstiziale.<br />
rappresentano attualmente gli organismi più spesso<br />
indiziati di tale paternità filogenetica, poiché<br />
contrariamente agli adulti, tali forme giovanili sono<br />
attive nuotatrici. Basta ammettere la concomitanza<br />
di due eventi non insoliti tra gli animali: il<br />
mantenimento delle caratteristiche larvali (neotenia) e<br />
la capacità di una riproduzione sessuale in fase<br />
larvale (pedogenesi), per accettare la nostra<br />
discendenza da forme immature di questi barilotti<br />
filtranti!<br />
Figura 22. Le lamprede.<br />
Molte specie sono parassiti di altri pesci e utilizzano la loro<br />
grande bocca circolare per raschiare la carne e succhiare il<br />
sangue.
Le circa 50 specie di lamprede vivono sia nelle acque<br />
dolci sia in quelle costiere, per poi trasferirsi nei fiumi<br />
durante la stagione riproduttiva. Le lamprede<br />
possiedono una scatola cranica completa e vertebre<br />
vere (sebbene rudimentali) formate da cartilagine. Prima<br />
di trasformarsi in adulti, le larve delle lamprede si<br />
nutrono per filtrazione e hanno un aspetto piuttosto<br />
simile agli anfiossi adulti. Gli adulti di molte specie di<br />
lamprede sono parassiti, sebbene numerose linee<br />
evolutive abbiano dato origine ad adulti che non si<br />
nutrono. In quest’ultimo caso, dopo la metamorfosi la<br />
lampreda adulta sopravvive per poche settimane, giusto<br />
il tempo per potersi riprodurre. Nelle specie che da<br />
adulte conducono una vita parassita, la bocca circolare<br />
è trasformata in un organo che svolge le funzioni di una<br />
ventosa, usata per fissarsi alla preda e raschiarne la<br />
carne (▶figura 23).<br />
Figura 23. Bocca di una lampreda.<br />
Si possono notare sia il labbro circolare, tipico delle lamprede,<br />
sia i dentelli cornei con i quali, questi ectoparassiti lacerano i<br />
tessuti delle loro vittime.<br />
Figura 24. Le missine.<br />
Le missine vivono poggiate sul fango dei fondali, da cui<br />
estraggono piccole prede.<br />
Quadro anatomico dei ciclostomi:<br />
il corpo è allungato e cilindrico<br />
lo scheletro è interamente cartilagineo<br />
la corda dorsale è persistente<br />
l’arco orale non è suddiviso in mascella e<br />
mandibola (agnati)<br />
la bocca è dotata, in alcune specie, di un<br />
labbro disposto circolarmente<br />
le branchie, poste in tasche branchiali<br />
indipendenti, sono dotate di fori<br />
branchiali<br />
il cuore possiede un atrio e un ventricolo<br />
la circolazione è semplice e completa<br />
il prorene spesso è funzionale anche<br />
nell’adulto<br />
l’N è eliminato come ammoniaca<br />
(ammoniotelici)<br />
la temperatura corporea è identica a<br />
quella dell’ambiente circostante<br />
il midollo allungato (bulbo) è il centro che<br />
controlla l’attività nervosa<br />
l’olfatto e il gusto sono gli organi di senso<br />
più sviluppati<br />
la fecondazione è esterna<br />
Altri ciclostomi, i missinoidei, dispongono di una<br />
bocca agnata più primitiva e vivono dei tessuti in<br />
disfacimento delle carogne di altri vertebrati.<br />
Le missine (▶figura 24) sono animali marini, quasi<br />
ciechi ma con quattro tentacoli sensoriali disposti<br />
intorno alla bocca. Questi animali sono provvisti di una<br />
struttura simile a una lingua, dotata di dentelli adatti a<br />
raschiare, che vengono usati per strappare pezzi di<br />
carne da organismi morti (carogne) e per catturare<br />
piccoli invertebrati. Le missine hanno sviluppo diretto<br />
(cioè privo di uno stadio larvale); gli individui inoltre<br />
possono cambiare sesso da un anno all’altro (da<br />
maschio a femmina e viceversa).<br />
Le parole:<br />
Agnati deriva da a privativa e da gnáthos, «mascella» e<br />
letteralmente significa, privi di mascelle.<br />
Gnatostomi viene da gnáthos, «mascella», e stóma,<br />
«bocca». Il termine pone l’accento sulla più importante<br />
innovazione evolutiva di questo gruppo.<br />
Ciclostoma deriva dal greco kýklos, «cerchio», e stóma,<br />
«bocca». Il nome pone l’accento sulla caratteristica<br />
tipica di questi animali, che possono sembrare pesci,<br />
ma hanno una bocca circolare e sempre aperta.<br />
Ostracodermi deriva dal greco ostráx, «conchiglia»,<br />
e derma, «pelle» e fa riferimento alla pelle corazzata di<br />
questi animali.
Caratteristiche generali dei pesci<br />
propriamente detti.<br />
Solo più tardi nel corso dell'evoluzione i denti saranno<br />
utilizzati per la masticazione. La presenza di un arco<br />
orale dotato di denti dischiude agli gnatostomi la<br />
I pesci hanno subito diversi milioni di anni<br />
di evoluzione negli ambienti acquatici prima che i primi<br />
vertebrati iniziassero a colonizzare le terre emerse e a<br />
diversificarsi in un numero enorme di specie. II periodo<br />
possibilità di nutrirsi di organismi voluminosi.<br />
L'attività predatoria si associa inoltre a notevoli progressi<br />
nel sistema nervoso, in quello sensoriale e in quello<br />
muscolare.<br />
del Paleozoico noto come Devoniano viene definito<br />
«età dei pesci», poiché in quel tempo comparve sugli<br />
scenari evolutivi una straordinaria varietà di tali<br />
vertebrati, sia nelle acque interne che nel mare. Il gruppo<br />
dei pesci non rappresenta una categoria sistematica<br />
unitaria; essa comprende i pesci a scheletro osseo e<br />
quelli a scheletro cartilagineo, oltre agli estinti placodermi.<br />
Occorre rilevare che anche attualmente i pesci ossei<br />
costituiscono il gruppo dei vertebrati più rappresentativo<br />
in termini numerici assoluti e per diversità di specie. Nel<br />
Primario agli agnati si aggiunsero altri vertebrati nei quali<br />
una coppia degli archi scheletrici di sostegno alle fessure<br />
branchiali (la seconda o la terza coppia) si era trasformata<br />
in una sorta di tagliola, definita arco orale, con un<br />
elemento dorsale (mascella) e uno ventrale (mandibola), in<br />
grado di aprire e chiudere la fessura (rima) buccale<br />
(▶figura 25A). L'arco orale incernierato rappresenta<br />
un'acquisizione rivoluzionaria. Il valore adattativo di tale<br />
apparato muscolo-scheletrico risiede nella capacità di<br />
Sono pervenuti resti paleontologici dei primi vertebrati<br />
gnatostomi (cioè con bocca provvista di fauci incernierate),<br />
appartenenti al gruppo dei placodermi (▶figura 26). Si<br />
tratta di organismi che condividevano il robusto<br />
esoscheletro dei progenitori ostracodermi, ma che, oltre<br />
alla presenza di un arco orale articolato, disponevano<br />
anche di una sagoma più idrodinamica e delle prime<br />
pinne pari. Queste strutture, sia pure nella loro<br />
primitività, attestano una migliore manovrabilità in<br />
acqua, consentita anche da una certa flessibilità del<br />
l'esoscheletro del tronco, che era costituito da elementi più<br />
piccoli e numerosi rispetto a quello degli ostracodermi.<br />
Alcuni placodermi raggiungevano dimensioni<br />
ragguardevoli e rappresentavano, con i calamari, i<br />
predatori più attivi negli oceani del Devoniano. Nonostante<br />
il loro temporaneo successo evolutivo la maggior parte<br />
dei placodermi non ha superato tale periodo e nessuno ha<br />
valicato il Paleozoico.<br />
afferrare saldamente la preda, che viene poi trasferita nella<br />
faringe e nei tratti posteriori del canale digerente. A tale<br />
funzione concorrono i denti (▶figura 25B) che<br />
consentono di trattenere la preda in modo più efficace.<br />
Figura 26. Placodermi.<br />
(A) Ricostruzione intera di Coccosteus.<br />
(B) Cranio di Dunkleosteous nel quale si possono apprezzare i<br />
denti e le placche che circondano l’occhio.<br />
Figura 25. Le mascelle e i denti hanno aumentato l’efficienza<br />
nella ricerca di cibo.<br />
(A) Questa serie di schemi illustra un possibile percorso<br />
evolutivo dell’arcata mandibolare a partire dagli archi branchiali<br />
anteriori dei pesci. (B) L’arcata mandibolare dello squalo gigante<br />
(Carcharodon megalodon), oggi estinto, è un buon esempio di<br />
dentatura tipica di uno stile di vita predatorio<br />
Le parole:<br />
Placodermi deriva dal greco pláx, «placca», e derma,<br />
«pelle» e fa sempre riferimento alla pelle corazzata di<br />
questi animali.
Il piano strutturale di un pesce, con le caratteristiche<br />
tipiche dei vertebrati in evidenza, è illustrato<br />
nella ▶figura 27: lo scheletro assile costituito<br />
dal cranio che protegge l’encefalo, posto anteriormente<br />
e dalla colonna vertebrale che protegge il midollo<br />
spinale che fa seguito al cranio.<br />
un’enorme superficie allo scambio dei gas, le branchie<br />
sono presenti non soltanto nei pesci, ma anche in molti<br />
altri gruppi animali, tra cui gli artropodi, i molluschi, gli<br />
anellidi e, tra i vertebrati, gli anfibi (nelle forme larvali e<br />
anche in alcune forme adulte). Nei pesci, le branchie<br />
sono organizzate intorno agli archi branchiali, strutture<br />
rigide che forniscono supporto e protezione, oltre a<br />
comprendere le vene e le arterie in cui scorre il sangue<br />
fino ai filamenti branchiali (▶figura 28). I filamenti<br />
(o lamelle) branchiali sono disposti fittamente in file<br />
lungo gli archi (di solito vi sono quattro archi branchiali<br />
per ciascun lato del pesce) e formano un vero e proprio<br />
filtro in cui scorre, in modo forzato, l’acqua. Nei<br />
Figura 27. Il piano strutturale dei pesci.<br />
Il disegno riassume gli elementi strutturali comuni a tutti i<br />
vertebrati. Oltre alle pinne pelviche pari (le pinne posteriori),<br />
questi pesci possiedono pinne pettorali (le «pinne anteriori») su<br />
entrambi i lati del corpo (che non sono visibili nella figura).<br />
I pesci sono vertebrati dotati di pinne che permettono<br />
loro di stabilizzare la propria posizione e di nuotare. La<br />
maggior parte dei pesci possiede una coppia di<br />
pinne pettorali (che corrispondono agli arti anteriori dei<br />
tetrapodi) appena posteriormente alle fessure branchiali<br />
e una coppia di pinne pelviche (che corrispondono agli<br />
arti posteriori dei tetrapodi), anteriormente alla zona<br />
anale.<br />
Oltre alle pinne pari e simmetriche, ci sono poi, pinne<br />
impari e mediane, come la pinna dorsale e<br />
filamenti branchiali scorre un fitto sistema<br />
di capillari sanguigni, la sede in cui avviene<br />
effettivamente lo scambio dei gas.<br />
Il sistema di ventilazione, quel meccanismo per cui<br />
l’acqua si trova a scorrere a ritmo forzato lungo i<br />
filamenti, fa in modo che per un pesce sia sufficiente<br />
aprire la bocca per far penetrare l’acqua che andrà a<br />
fluire nelle branchie. L’apertura della bocca da una<br />
parte e la chiusura alternata degli opercoli branchiali (i<br />
«coperchietti» rigidi che ricoprono le branchie<br />
all’esterno) dall’altra mantengono e modulano il flusso<br />
dell’acqua secondo le esigenze della respirazione.<br />
Questo è il motivo per cui i pesci continuano a<br />
«boccheggiare».<br />
quella caudale, che contribuiscono a stabilizzare il<br />
pesce promuovendo il movimento, o ancora<br />
permettendo all’animale di cambiare rapidamente<br />
direzione. Spesso è presente anche una pinna anale<br />
posta in prossimità dell’apertura anale.<br />
I pesci dispongono, inoltre, di una struttura sensoriale<br />
detta sistema della linea laterale, costituita da una fila<br />
di recettori che corre lungo ciascun lato del corpo<br />
dell’animale; questo sistema è sensibile alle variazioni<br />
di pressione e consente di registrare le minime<br />
vibrazioni provocate dai pesci che nuotano nelle<br />
vicinanze, oppure di rilevare gli ostacoli.<br />
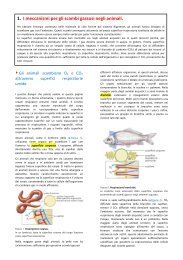
Le branchie sono un sistema efficientissimo nella<br />
cattura dell’ossigeno presente nell’acqua, che è soltanto<br />
1/20 di quello percentualmente presente nell’aria, a<br />
parità di volume. Conformate in modo da offrire<br />
Figura 28. Le branchie dei pesci.<br />
(A) L’acqua fluisce sulle branchie di un pesce in maniera<br />
unidirezionale. (B) Le lamelle branchiali di primo ordine sono<br />
caratterizzate da un’ampia superficie di scambio e da tessuti<br />
molto sottili. (C) Il sangue scorre attraverso le lamelle di<br />
secondo ordine nella direzione opposta (da sinistra a destra) a<br />
quella in cui fluisce l’acqua (da destra a sinistra).
I condroitti sono pesci con<br />
scheletro cartilagineo.<br />
L'endoscheletro dei condroitti è costituito da tessuto<br />
cartilagineo opportunamente infiltrato di sali; si tratta di<br />
un connettivo specializzato per il sostegno, caratterizzato<br />
I condroitti o pesci cartilaginei costituiscono una classe<br />
da robustezza, da leggerezza e da flessibilità.<br />
di pesci comparsa durante il Devoniano che attualmente<br />
comprende gli squali, le razze e le chimere (per un<br />
totale di circa 1000 specie).<br />
Di regola gli squali sono eccellenti nuotatori grazie anche<br />
alla presenza di due coppie di pinne pari, una posteriore<br />
alla regione branchiale, l'altra anteriore alla regione<br />
Come gli agnati, anche questi pesci possiedono uno<br />
scheletro interamente formato da un materiale rigido<br />
ma flessibile, la cartilagine. Immediatamente dietro<br />
all'arco orale, si differenzia dal «cestello» faringeo<br />
un'ulteriore struttura scheletrica, definita arco ioideo.<br />
L'apparato cutaneo di queste forme, quasi esclusivamente<br />
marine, non è apparentemente dotato di strutture ossee<br />
di protezione e appare del tutto nudo. In realtà,<br />
cloacale, che consentono all'animale di dirigere<br />
efficacemente il nuoto (▶figura 30). Le pinne pari<br />
corrispondono agli arti delle forme terrestri e<br />
appartengono a tutti gli effetti al piano strutturale dei<br />
vertebrati. Il movimento degli squali è garantito dai potenti<br />
movimenti della porzione caudale del tronco provvista di<br />
una caratteristica pinna a lobo dorsale più sviluppata<br />
(coda eterocerca).<br />
soprattutto gli squali dispongono di particolari scaglie<br />
definite placoidi o dentelli cutanei, poiché risultano<br />
conformati e strutturati come veri denti; la loro pelle,<br />
opportunamente trattata, può essere impiegata come<br />
un'eccellente carta vetrata (zigrino).<br />
I denti posti nella<br />
bocca sono a crescita continua (▶figura 29).<br />
Figura 30. Squalo bianco.<br />
Il grande squalo bianco, Carcharodon carcharias, chiamato<br />
anche carcarodonte o talvolta semplicemente pescecane, è il più<br />
grande pesce predatore del pianeta.<br />
Le razze, le mante e le torpedini, che hanno una coda<br />
filiforme, nuotano invece utilizzando le ampie pinne<br />
pettorali (▶figura 31 e 32).<br />
Figura 29. Dente di squalo.<br />
Dente di squalo bianco, Carcharodon carcharias (lato interno).<br />
Inconfondibile per le sue caratteristiche uniche: dente grande,<br />
triangolare, dritto, fortemente seghettato ai margini. I denti<br />
delle mandibole superiore ed inferiore sono molto simili.<br />
Il notevole alleggerimento dello scheletro cutaneo è stato<br />
favorito dal contemporaneo miglioramento delle capacità<br />
cinetiche che rende loro possibile di sfuggire celermente<br />
ai predatori. D'altra parte il ruolo dei condroitti nel<br />
proprio ecosistema è quello dei predatori piuttosto che<br />
quello delle prede. La notevole riduzione dello scheletro<br />
cutaneo si è realizzata contemporaneamente alla<br />
scomparsa secondaria del tessuto osseo interno.<br />
Quasi tutti i pesci cartilaginei sono marini, ma alcuni<br />
vivono nelle acque di estuario o migrano in laghi e<br />
fiumi. Gli squali più grandi si nutrono di plancton, come<br />
lo squalo balena che con i suoi 18 m rappresenta il più<br />
grande pesce vivente. La maggior parte delle specie<br />
comprende formidabili predatori dotati di un corpo<br />
affusolato, di una notevole velocità, di mascelle potenti<br />
e di organi di senso estremamente sensibili; oltre alla<br />
vista acuta, all’olfatto molto sviluppato e all’organo<br />
della linea laterale, gli squali possiedono infatti speciali<br />
elettrosensori capaci di rivelare i debolissimi campi<br />
elettrici prodotti dalle contrazioni muscolari delle<br />
potenziali prede.
A differenza dei pesci ossei, gli squali non sono dotati<br />
di una muscolatura faringea in grado di creare una forte<br />
corrente d’acqua sulle branchie: affinchè gli scambi<br />
gassosi possano avvenire, però, lo scorrimento<br />
dell’acqua attraverso le branchie deve essere continuo,<br />
altrimenti lo squalo rischierebbe di morire per asfissia.<br />
Per questo motivo, le modalità di respirazione sono<br />
diverse a seconda delle specie: gli squali pelagici, che<br />
nuotano in mare, devono muoversi in continuazione<br />
tenendo la bocca semiaperta per permettere il<br />
passaggio dell’acqua e l’ossigenazione delle branchie.<br />
Recenti studi hanno dimostrato che alcune specie<br />
rimangono immobili disponendosi controcorrente: in<br />
questo modo sfruttano le correnti per garantire un<br />
flusso costante di acqua sulle branchie.<br />
Figura 32. Torpedine.<br />
Torpedo torpedo ha forma appiattita ed è caratterizzata dalla<br />
presenza, ai lati del corpo, di organi elettrogeni, derivati da<br />
muscoli, in grado di produrre scariche elettriche sensibili anche<br />
per l'uomo.<br />
Le chimere sono i pesci cartilaginei meno noti, e vivono<br />
nelle acque profonde e fredde (▶figura 33).<br />
Gli squali bentonici, cioè che nuotano a stretto contatto<br />
con il fondo, possono anche rimanere fermi per lunghi<br />
periodi, aspirando l’acqua con la bocca e spingendola<br />
forzatamente attraverso le branchie.<br />
Alcuni squali hanno recuperato abitudini alimentari basate<br />
sulla filtrazione e vivono di plancton, una fonte alimentare<br />
che ha consentito loro di raggiungere dimensioni<br />
ragguardevoli; si pensi al gigantesco squalo-balena<br />
(Rhincodon typhus) che può raggiungere una lunghezza di<br />
15 m e un peso di 9000 kg!<br />
Un componente collegato alla respirazione degli squali<br />
è lo spiracolo, un piccolo foro posto dietro l’occhio che<br />
si apre e si chiude grazie all’azione di un muscolo<br />
involontario. Lo spiracolo viene utilizzato dal pesce<br />
come una pompa, per aspirare l’acqua e spingerla verso<br />
le branchie integrando l’azione della bocca.<br />
La maggior parte delle razze, caratterizzate da un corpo<br />
fortemente appiattito, vive invece sui fondali<br />
dell’oceano, dove si nutre di molluschi e di altri animali<br />
infossati nei sedimenti.<br />
Figura 31. Manta gigante.<br />
Manta birostris, ha il corpo costituito da un disco, distinto dalla<br />
coda, piatto e depresso. Gli occhi sono sul dorso mentre sul<br />
ventre ci sono la bocca e le fessure branchiali. Quest'ultime<br />
sono munite di un filtro per il plancton e sono larghe. La pelle è<br />
zigrinata, con piccole o grandi spine.<br />
Figura 33. Chimera.<br />
Chimaera monstrosa, o pesce ratto, vive nelle profondità<br />
oceaniche e possiede spesso pinne dorsali modificate che<br />
contengono tossine.<br />
Quadro anatomico dei condroitti:<br />
la cute è rivestita da scaglie dure e appuntite<br />
(squame placoidi)<br />
lo scheletro è interamente cartilagineo<br />
l’arco orale è suddiviso in mascella e<br />
mandibola (gnatostomi)<br />
la bocca è in posizione ventrale<br />
la coda è eterocerca (è cioè suddivisa in due<br />
lobi asimmetrici)<br />
le branchie, in tasche branchiali indipendenti,<br />
hanno fessure branchiali<br />
nell’intestino è presente una lunga piega che<br />
aumenta l’assorbimento (valvola spirale)<br />
il cuore possiede un atrio e un ventricolo<br />
la circolazione è semplice e completa<br />
il prorene è sostituito dal mesorene<br />
l’N è eliminato come urea (ureotelici)<br />
la regolazione osmotica si attua mantenendo<br />
la concentrazione dei soluti nei liquidi interni<br />
uguale a quella esterna<br />
la temperatura corporea è identica a quella<br />
dell’ambiente circostante<br />
il midollo allungato (bulbo) è il centro che<br />
controlla l’attività nervosa dell’animale e il<br />
cervelletto è molto sviluppato<br />
l’olfatto e il gusto sono gli organi di senso più<br />
sviluppati<br />
la fecondazione è interna; il maschio possiede<br />
organi copulatori<br />
le uova sono grandi, ricche di tuorlo e protette<br />
da una dura guaina
Gli osteoitti sono pesci con<br />
scheletro osseo.<br />
Per quanto la storia evolutiva dei pesci conduca sempre al<br />
mare, si ritiene che il loro ambiente ancestrale fosse<br />
rappresentato dalle acque dolci, o al più dalle acque di<br />
estuario, a concentrazione salina intermedia. Questo vale<br />
tanto per i condroitti che per gli osteoitti o pesci ossei,<br />
l'altra classe degli gnatostomi a esclusivo e primitivo<br />
habitat acquatico attualmente viventi. Il nome di questi<br />
organismi deriva dal fatto che essi dispongono di uno<br />
scheletro interno osseo, in contrapposizione a quello<br />
cartilagineo dei condroitti.<br />
L'ambiente ancestrale degli osteoitti era rappresentato da<br />
raccolte d'acqua stagnante, esposte a rischi di<br />
prosciugamento e di regola poco ossigenate. In tali<br />
condizioni gli osteoitti hanno messo a punto un elaborato<br />
apparato branchiale, con un opercolo sorretto da uno<br />
scheletro specifico, che esclude ed espone<br />
alternativamente le fessure nei confronti dell'ambiente,<br />
coadiuvando la loro funzione e facilitando il flusso<br />
dell'acqua attraverso di esse.<br />
In passato un opercolo simile, ma non derivato dalle<br />
medesime strutture ossee, proteggeva la camera<br />
branchiale dei placodermi, mentre i condroitti ne sono<br />
sprovvisti, con l'eccezione delle chimere, che tuttavia<br />
presentano un opercolo puramente carnoso.<br />
I primitivi pesci ossei, e forse gli stessi placodermi,<br />
possedevano organi ancestrali per coadiuvare la<br />
respirazione branchiale, difficoltosa in acque poco<br />
ossigenate: le sacche polmonari. Queste strutture, connesse<br />
con il cavo orale, potevano essere riempite d'aria, che il<br />
pesce abboccava sulla superficie dell'acqua, e scambiare<br />
gas con il sangue contenuto nei capillari arteriosi aderenti<br />
alla loro sottile parete. Attualmente limitate a poche<br />
forme arcaicizzanti, nelle quali talvolta non presentano più<br />
funzioni respiratorie, le sacche polmonari hanno consentito a<br />
un gruppo di primitivi pesci ossei la conquista<br />
dell'ambiente subaereo.<br />
Nelle forme più moderne, i teleostei , le sacche si sono<br />
trasformate in un efficacissimo organo idrostatico impari, la<br />
vescica natatoria. I ruoli di questo organo cavo sono<br />
molteplici, potendo funzionare come camera di risonanza<br />
per la percezione del suono, come organo di senso<br />
recettore della pressione e, infine, come dispositivo<br />
variabile di equilibrio e di galleggiamento a varie<br />
profondità, tramite la regolazione della quantità e della<br />
qualità dei gas contenuti.<br />
Gli osteoitti si sono suddivisi in tre linee evolutive:<br />
1) Attinopterigi (hanno pinne pari con raggi<br />
disposti da un solo lato: pinne uniseriali)<br />
2) Dipnoi (hanno pinne pari peduncolate)<br />
3) Crossopterigi (hanno pinne pari con raggi<br />
disposti da entrambi i lati: pinne biseriali)<br />
Dipnoi e Crossopterigi vengono inclusi nella sottoclasse<br />
dei Sarcopterigi (con pinne carnose).<br />
Gli attinopterigi, pesci ossei a pinne<br />
raggiate, sono il gruppo più<br />
rappresentato tra i vertebrati attuali.<br />
Gli attinopterigi possiedono oltre ad uno scheletro<br />
formato da tessuto osseo, anche pinne pari a scheletro<br />
raggiato, molto efficienti per dirigere il nuoto<br />
che li<br />
distinguono dai sarcopterigi. Queste pinne pari<br />
presentano i raggi disposti da un solo lato dell’asse e<br />
sono pertanto definite pinne uni seriali.<br />
Gli attinopterigi si sono diversificati durante il Terziario,<br />
fino a dare origine a circa 30 000 specie di acque dolci e<br />
marine caratterizzate da taglia, forma, tipo di<br />
alimentazione e ciclo biologico estremamente variabili<br />
(▶dalla figura 34 alla figura 45): questa straordinaria<br />
abbondanza di specie fa degli attinopterigi il gruppo più<br />
rappresentato di vertebrati moderni.<br />
La superficie esterna del corpo degli acttinopterigi è<br />
rivestita da squame sottili e leggere, che forniscono un<br />
certo grado di protezione e facilitano il movimento<br />
nell’acqua; le branchie si aprono in una singola camera<br />
coperta da un rigido lembo di tessuto, detto opercolo: il<br />
movimento dell’opercolo aumenta il flusso dell’acqua<br />
attraverso le branchie, dove hanno luogo gli scambi<br />
gassosi.<br />
Negli antenati dei pesci attuali, la funzione respiratoria<br />
delle branchie era aumentata da sacche simili a<br />
polmoni, che permettevano loro di sopravvivere in<br />
acque povere di ossigeno. Nella maggior parte dei pesci<br />
ossei queste sacche hanno dato origine alla vescica<br />
natatoria, un organo di spinta idrostatica: regolando la<br />
quantità di gas presente nella vescica natatoria, il pesce<br />
è in grado di controllare la profondità, spendendo poca<br />
energia per mantenere la propria posizione.<br />
Gli attinopterigi rappresentano i vertebrati dominanti nei<br />
mari e nelle acque interne, date le loro notevoli capacità<br />
adattative.
Gli attinopterigi più evoluti corrispondono ai teleostei: si<br />
tratta di organismi con forme, dimensioni e quadri<br />
comportamentali estremamente variabili, che vivono<br />
predando tutti gli altri organismi marini, vertebrati<br />
compresi, con l'eccezione dei cetacei.<br />
Teleostei notevolmente adattati si rinvengono anche nelle<br />
acque interne. Essi presentano notevoli specializzazioni<br />
nell'apparato genitale e strategie riproduttive talvolta<br />
complesse. Per quanto si descrivano nei due sessi<br />
comportamenti elaborati che tendono a rendere non<br />
casuale la fecondazione delle uova, l'incontro dei gameti<br />
avviene nell'ambiente esterno, dove si compie anche lo<br />
sviluppo embrionale, con un alto tasso di mortalità per gli<br />
zigoti e per gli embrioni.<br />
Figura 37. Pesce pagliaccio.<br />
Il pesce pagliaccio, Amphiprion percula. Diffuso nell'Oceano<br />
Indiano e pacifico, frequenta le acque calme delle barriere<br />
coralline ed è particolarmente frequente nelle lagune degli atolli<br />
Vive in simbiosi mutualistica con anemoni di mare al cui veleno<br />
è immune. Comune ospite degli acquari marini e vi si può<br />
riprodurre.<br />
Figura 34. Storione.<br />
Lo storione, Acipenser sturio, è il più grande pesce d’acqua<br />
dolce e salmastra diffuso in Europa. E’ famoso per le sue carni<br />
pregiate e per le sue uova (caviale).<br />
Figura 38. Cavalluccio marino.<br />
Il cavalluccio marino Hippocampus, è così chiamato per via della<br />
testa che ricorda quella di un piccolo cavallo. I cavallucci marini<br />
si trovano in tutte le acque del mondo tranne quelle glaciali,<br />
prevalentemente in prossimità delle coste dove trovano rifugio e<br />
sostegni dove potersi ancorare durante i movimenti con la lunga<br />
coda prensile. Sono particolarmente diffusi nelle barriere<br />
coralline. La femmina depone le uova in una speciale sacca<br />
incubatrice nel ventre del maschio, situata vicino all'apertura<br />
anale. Alla schiusa, il maschio espelle gli avannotti con delle<br />
contrazioni addominali simili al parto femminile, evento<br />
piuttosto insolito in natura, chiamato gravidanza maschile.<br />
Figura 35. Salmone.<br />
Il salmone dell'Atlantico, Salmo salar, è un pesce predatore<br />
tipico dei mari freddi del nord Atlantico. È un<br />
pesce anàdromo (dal greco anádromos, composto da aná<br />
'all'indietro' e drómos 'corsa') in quanto nasce in acqua dolce,<br />
passa la fase centrale della sua vita in mare e torna in acqua<br />
dolce per riprodursi<br />
Figura 36. Tonno rosso.<br />
Il tonno rosso, Thunnus thynnus, è un grande pesce pelagico.<br />
Conosciuto anche come tonno pinna blu.<br />
Figura 39. Pesce luna.<br />
Il pesce luna, Mola mola, è il più grande tra i pesci ossei. Abita il<br />
mare aperto, ma è localizzato prevalentemente lungo le acque<br />
costiere. E’ caratterizzato da una forma allungata, ovaloide,<br />
molto compressa ai fianchi. I denti sono fusi tra loro nella<br />
piccola bocca e formano una sorta di becco. La sua pelle può<br />
raggiungere lo spessore di 15 cm e ospita<br />
parassiti e microorganismi che possono provocare il fenomeno<br />
della bioluminescenza. Raggiunge la lunghezza di 3 m per<br />
un'altezza di 3 m ed un peso di oltre due tonnellate. Si tratta<br />
inoltre di un pesce estremamente longevo: presumibilmente<br />
può superare ampiamente i cento anni di età.
Figura 40. Murena.<br />
La murena mediterranea, Muraena helena è un pesce d'acqua<br />
salata diffusa nel Mar Mediterraneo e nell'Atlantico orientale.<br />
Essendo un animale schivo preferisce le coste rocciose ricche di<br />
anfratti, da dove esce solo per cacciare.<br />
Figura 43. Sogliola.<br />
Le sogliole, Solea solea, sono pesci la cui evoluzione li ha<br />
portati alla postura sdraiata su un fianco, con uno dei due occhi<br />
che si è spostato a fianco dell'altro, sul lato rivolto verso la<br />
superficie dell'acqua. Anche la colorazione ha seguito<br />
quest'evoluzione: il lato rivolto verso il fondo è bianco, quello<br />
verso l'alto è bruno, marezzato, mimetico e spesso<br />
camaleontico: le sogliole riescono infatti a cambiare colore per<br />
imitare l'ambiente circostante grazie ai cromatofori, cellule con<br />
pigmenti colorati dell'epidermide.<br />
Figura 41. Piranha.<br />
I piranha sono un gruppo di pesci d'acqua dolce che vivono<br />
in fiumi e lagune del Sudamerica. Normalmente sono lunghi dai<br />
15 ai 25 cm e raramente superano i 40 cm. Sono celebri per i<br />
loro denti affilati, capaci di tranciare un amo, e un appetito<br />
vorace per la carne.<br />
Figura 44 Remora.<br />
La remora, Echeneis naucrates, è caratterizzata dalla presenza<br />
di una vistosa ventosa dorsale con la quale si attacca a grandi<br />
animali marini come squali, tartarughe marine o cetacei da cui<br />
si fa trasportare. Si nutre di pesciolini o avanzi del pasto del suo<br />
ospite. Talvolta può attaccarsi ai subacquei.<br />
Figura 42. Pesce combattente.<br />
Il pesce combattente, Betta splendens, è un pesce d’acqua dolce<br />
che vive nel Sud-est asiatico. Abita acque stagnanti, risaie,<br />
canali, specchi lacustri e aree fluviali con deboli correnti,<br />
sopportando acque poco ossigenate. Il maschio è molto<br />
territoriale ed è aggressivo nei confronti di altri maschi. E’<br />
diventato comune tra gli acquariofili i quali comunemente<br />
tengono un solo esemplare maschi in ogni acquario.<br />
Figura 45. Carpa.<br />
La carpa, Cyprinus carpa, è un pesce d’acqua dolce molto<br />
diffuso in Italia che predilige i fiumi a corso lento e i laghi.<br />
Originario delle regioni orientali d’Europa e dell’Asia minore, è<br />
stato introdotto molti secoli fa dai romani per l’allevamento.
I sarcopterigi, pesci ossei a pinne<br />
lobate, comprendono gli antenati dei<br />
vertebrati terrestri.<br />
La presenza di sacche polmonari in un gruppo ancestrale<br />
di pesci ossei rappresenta il principale preadattamento alla<br />
conquista dell'ambiente subaereo. La disponibilità di tali<br />
organi non solo consentiva di integrare la respirazione<br />
branchiale in acque povere di ossigeno, ma rendeva anche<br />
possibili brevi spostamenti nell'ambiente terrestre, alla<br />
ricerca di raccolte d'acqua più ospitali. La pinna raggiata,<br />
efficiente per dirigere il nuoto, rappresenta un mediocre<br />
supporto sul terreno; i pesci che si impegnarono nei primi<br />
tragitti fuori dall'acqua erano in realtà provvisti di un tipo<br />
di appendice del tronco assai primitivo ma adatto a tali<br />
spostamenti. Si trattava di una pinna di aspetto carnoso e<br />
provvista di un asse scheletrico articolato, sul quale si<br />
impiantavano segmenti marginali, paralleli piuttosto che<br />
raggiati. Per l'aspetto carnoso della loro pinna tali pesci<br />
ossei si definiscono sarcopterigi.<br />
I crossopterigi sono dotati di pinne pari con i raggi<br />
disposti da entrambi i lati (pinne biseriali).<br />
Comprendono i celacanti e i progenitori (fossili) dei<br />
vertebrati terrestri.<br />
I crossopterigi celacanti furono abbondanti a partire dal<br />
Devoniano fino a circa 65 milioni di anni fa, quando si<br />
estinsero quasi completamente. Casualmente, nel 1938<br />
un peschereccio catturò un celacanto al largo delle<br />
coste del Sudafrica. Da allora sono stati pescati<br />
centinaia di individui della specie Latimeria chalumnae,<br />
che è considerata un «fossile vivente». Una seconda<br />
specie, L. menadoensis, è stata scoperta nel 1998 al<br />
largo dell’isola indonesiana di Sulawesi. I pesci del<br />
genere Latimeria sono predatori e raggiungono una<br />
lunghezza di circa 1,8 m e un peso fino a 82 kg<br />
(▶figura 47). Entrambe le specie sono forme aberranti di<br />
dimensioni considerevoli, che nel corso del loro<br />
adattamento alle acque marine si sono evolute in modo<br />
specialistico.<br />
Attualmente il gruppo comprende i dipnoi e i crossopterigi.<br />
I dipnoi, anch’essi provvisti di pinne lobate e articolate,<br />
erano importanti predatori nelle acque basse durante il<br />
periodo Devoniano. La maggior parte delle linee<br />
evolutive di questi pesci attualmente è estinta: le sole<br />
sei specie sopravvissute vivono nelle paludi e nelle<br />
acque fangose del Sudamerica, dell’Africa e<br />
dell’Australia (▶figura 46).<br />
Figura 47. I parenti più prossimi dei vertebrati terrestri.<br />
Questo celacanto, che vive nelle acque profonde dell’oceano<br />
Indiano, rappresenta una delle due specie di celacanti<br />
sopravvissute fino ai giorni nostri.<br />
Le parole:<br />
Actinopterigi deriva dal greco aktís, «raggio»<br />
e pterygion, diminutivo di pteryx «ala», qui nel senso di<br />
pinna. Questi pesci hanno infatti le pinne raggiate, con<br />
raggi di sostegno disposti a ventaglio.<br />
Sarcopterigi deriva dal greco sarx, «carne» Questi pesci<br />
hanno infatti le pinne lobate, carnose.<br />
Figura 46. I dipnoi sono pesci polmonati.<br />
Tutte le specie attuali di pesci polmonati (come i dipnoi) vivono<br />
nelle acque dolci dell’emisfero meridionale.<br />
I dipnoi sono animali dotati sia di polmoni (derivati dalle<br />
sacche simili ai polmoni presenti nei loro antenati) sia di<br />
branchie. Quando uno stagno si prosciuga, i dipnoi<br />
possono infossarsi nel fango e sopravvivere per molti<br />
mesi in uno stato di sostanziale inattività, respirando<br />
direttamente l’aria atmosferica.<br />
Crossopterigi deriva dal greco krossós, «frangia»<br />
Questi pesci hanno infatti le pinne frangiate.<br />
Dipnoi deriva dal greco dipnos, «doppia respirazione».<br />
Questi pesci hanno infatti hanno una doppia<br />
respirazione: branchiale e polmonare.<br />
Latimeria chalumnae fu scoperto presso la foce del<br />
fiume Chalumna, in Sudafrica, dalla naturalista Marjorie<br />
Courtenay-Latimer. Chi identificò questo pesce scelse<br />
di dedicarlo al luogo e alla protagonista del<br />
ritrovamento.
I veri progenitori dei primi vertebrati terrestri, gli anfibi,<br />
devono essere ricercati tra i crossopterigi ripidisti, presenti<br />
attualmente solo come fossili, che a partire dal Devoniano<br />
intrapresero la conquista dell'ambiente subaereo.<br />
In modo apparentemente paradossale, i paleontologi<br />
affermano che l'adattamento all'ambiente subaereo dei<br />
ripidisti, dal quale ha preso l'avvio la colonizzazione delle<br />
terre emerse da parte dei vertebrati, sia avvenuto nel<br />
corso delle loro escursioni sul terreno per cercare un<br />
Figura 49. Tiktaalik roseae.<br />
Ricostruzione di un esemplare.<br />
ambiente acquatico più idoneo.<br />
L’evoluzione dei polmoni e delle pinne articolate, in<br />
grado di sostenere il pesce nelle acque basse, diede via<br />
alla colonizzazione delle terre emerse; in seguito,<br />
cambiamenti strutturali delle pinne consentirono ad<br />
alcuni pesci di sostenersi meglio nell’acqua bassa e,<br />
successivamente, di muoversi sulla terraferma.<br />
Dall’acqua alla terraferma.<br />
Si ritiene che alcuni crossopterigi acquatici primitivi<br />
abbiano iniziato a utilizzare fonti di cibo terrestri,<br />
adattandosi alla vita sulla terraferma e, infine,<br />
evolvendosi nei progenitori dei tetrapodi (vertebrati<br />
provvisti di quattro arti).<br />
Possiamo chiederci dunque da che cosa fu<br />
accompagnata la transizione da un animale che nuota<br />
nell’acqua a uno che cammina sulla terraferma.<br />
All’inizio del 2006, alcuni ricercatori hanno descritto la<br />
scoperta di un fossile del Devoniano caratterizzato da<br />
appendici che rappresenterebbero uno stadio<br />
intermedio tra le pinne dei pesci e gli arti dei tetrapodi<br />
terrestri (▶figure 48 – 49 - 50).<br />
Sembra che arti capaci di sollevare un pesce voluminoso<br />
con il movimento antero-posteriore, indispensabile per<br />
la deambulazione sulla terraferma, si siano evoluti<br />
quando gli animali vivevano ancora nell’acqua.<br />
Figura 48. Tiktaalik roseae.<br />
Questa specie fossile, scoperta di recente, fornisce ulteriori dati<br />
relativi all’evoluzione degli arti a partire dalle pinne pettorali<br />
Figura 50.Il passaggio dall’acqua alla terraferma.<br />
In questa figura si può notare quali trasformazioni hanno<br />
permesso di trasformare le pinne in arti.<br />
Quadro anatomico degli osteoitti:<br />
la cute è rivestita da scaglie ossee (squame<br />
cosmoidi o ganoidi)<br />
lo scheletro è prevalentemente costituito da<br />
tessuto osseo<br />
l’arco orale è suddiviso in mascella e<br />
mandibola (gnatostomi)<br />
la bocca non è in posizione ventrale<br />
la coda è generalmente omocerca (è cioè<br />
suddivisa in due lobi simmetrici)<br />
le branchie, contenute in una camera<br />
branchiale, sono protette da un opercolo<br />
branchiale<br />
è presente una sacca polmonare o un organo<br />
idrostatico (vescica natatoria)<br />
il cuore possiede un atrio e un ventricolo<br />
la circolazione è semplice e completa<br />
gli organi escretori sono rappresentati<br />
generalmente da mesoreni<br />
l’N è eliminato come ammoniaca<br />
(ammoniotelici)<br />
la temperatura corporea è identica a quella<br />
dell’ambiente circostante<br />
il mesencefalo è il centro che controlla<br />
l’attività nervosa<br />
la vista è l’organo di senso più sviluppato<br />
la fecondazione è esterna<br />
le uova sono numerose e di piccole dimensioni<br />
in diverse specie la determinazione del sesso è<br />
cromosomica