URA3 - Farmaciaunina2.it
URA3 - Farmaciaunina2.it
URA3 - Farmaciaunina2.it

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
Fonti naturali:<br />
Proteine di interesse<br />
farmaceutico<br />
Prodotti difficili da isolare<br />
Possono comportare reazioni immunitarie (diff. sp.)<br />
Contaminazione virale e patogenica<br />
Difficile soddisfare la domanda<br />
Proteine ricombinanti:<br />
Più economiche<br />
Più “safe”<br />
Rifornimento infinito
Processo a sei step:<br />
Isolamento del gene di interesse<br />
Introduzione del gene in un vettore di espressione<br />
Trasformazione nella cellula ospite<br />
Crescita delle cellule mediante processi di<br />
fermentazione<br />
Isolamento & purificazione della proteina<br />
Formulazione del prodotto finale
Batteri<br />
Vantaggi Svantaggi<br />
Crescono velocemente (8 hrs<br />
per produrre una proteina)<br />
Rese elevate (50-500 mg/L)<br />
Basso costo del terreno<br />
Costi di fermentazione<br />
contenuti<br />
Difficoltà di esprimere proteine di<br />
grosse dimensioni (>50 kD)<br />
Incapacità di glicosilare e rimuovere<br />
il peptide segnale<br />
Proteine eucariote a volte tossiche<br />
Incapacità di gestire proteine ricche<br />
di S-S<br />
Vantaggi Altri vantaggi<br />
Rese elevate (50-5000 mg/L)<br />
Basso costo del terreno<br />
Costi di fermentazione<br />
contenuti<br />
Lieviti<br />
Possono esprimere proteine di<br />
grosse dimensioni (>50 kD)<br />
Glicosilazione e rimozione del<br />
peptide segnale<br />
Presenza di chaperonine che<br />
consentono un folding corretto<br />
Gestiscono proteine ricche di S-S
LIEVITO:<br />
L’ E. coli degli Eucarioti
Complessità genomica<br />
Organism Size (MB) Genes genes/MB<br />
E. coli 4.6 4400 950<br />
Lievito 12.1 5800 480<br />
Drosophila 180 13700 80<br />
Uomo 2900 27000 9.3
Utilizzo del lievito come modello sperimentale:<br />
vantaggi<br />
• Terreno economico - LB simile<br />
• Duplicazione rapida - lievito - 90 min., E. coli - 20 min., uomo anni<br />
• Tool genetici ben sviluppati – Stati: aploide/diploide;<br />
Ricombinazione e isolamento mutanti<br />
• Geneticamente conservato – I lieviti sono più “vicini” all’uomo di E. coli<br />
• Completamente sequenziato- 5800 genes, 40% ORFs di funzione sconosciuta<br />
• Pochi introni- cDNA<br />
• Trasformabile – Modifica numero copie di geni; Eliminazione geni specifici<br />
• Buon sistema modello- Molti geni sono conservati tra uomo e lievito
Saccharomyces cerevisiae:<br />
Adattabilità a svariate condizioni ambientali<br />
Produzione vino e birra: tolleranza basso pH, alta conc etanolo<br />
Panificazione: produzione di CO 2 rapidamente da zucchero<br />
Cell factory: GRAS, facile da ingegnerizzare<br />
Drug screening: eucariota, ma facile da usare come batteri<br />
Sistema modello: studio del funzionamento delle cellule
Saccharomyces cerevisiae come SISTEMA MODELLO<br />
1997 – PRIMO eucariota sequenziato<br />
6607 ORFs<br />
Saccharomyces Genome Database<br />
http://www.yeastgenome.org/<br />
Genoma mitocondriale – 2micron circle<br />
Telomeri e centromeri semplici<br />
Ridondanza<br />
4885 ORFs, 73.94% 912 ORFs, 13.8% 810 ORFs, 12.26%
Candida albicans<br />
•Lievito patogeno;<br />
•ife
Schizosaccharomyces pombe:<br />
(Fission yeast)<br />
È utilizzato come organismo modello in biologia molecolare e<br />
cellulare<br />
Cancro<br />
Diabete<br />
Fibrosi cistica<br />
Danni DNA/errori replicazione<br />
Si riproduce mediante scissione binaria<br />
4800 ORF, 3 cromosomi, solitamente aploide<br />
Disacidificazione vini
Altri lieviti importanti:<br />
Kluyveromyces lactis: milk yeast; organismo modello<br />
sfruttato per la fermentazione su lattosio<br />
Pichia pastoris, Hansenula polymorpha, Yarrovia lipolytica:<br />
sistemi usati per la produzione di proteine<br />
Altri funghi importanti:<br />
Cryptococcus, Aspergillus, Neurospora....: Funghi<br />
filamentosi, microrganismi modello
Ciclo vitale dei lieviti<br />
Budding Yeast Fission Yeast
•Condensazione dei cromosomi non visibile<br />
•Inizio fuso fase S non G2<br />
•Membrana nucleare intatta<br />
•Dimensioni<br />
Ciclo cellulare del lievito
•Dimensioni aploide vs<br />
diploide<br />
•Homothallic<br />
Ciclo vitale del lievito<br />
Nitrogen starvation
Il sesso nei lieviti!<br />
Risposta ai fattori a- o alpha-<br />
Componenti conservate tra uomo e lievito<br />
Legame dei fattori a- o alpha- ad un recettore<br />
specifico<br />
Riorientamento della cellula<br />
Cascata MAP<br />
Arresto del ciclo cellulare<br />
Espressione geni necessari per la fusione<br />
1<br />
2<br />
Recettori transmembrana a<br />
Associate a proteine-G<br />
1: Orientamento della cellula<br />
2: Arresto del ciclo cellulare<br />
3: Espressione geni per la fusione<br />
3
•In natura…2n!<br />
•Mating type switch<br />
•Funzione del mating<br />
Formazione di un lievito diploide:
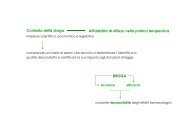
Processo a sei step:<br />
Isolamento del gene di interesse<br />
Introduzione del gene in un vettore di espressione<br />
Trasformazione nella cellula ospite<br />
Crescita delle cellule mediante processi di<br />
fermentazione<br />
Isolamento & purificazione della proteina<br />
Formulazione del prodotto finale
Come si sceglie il vettore<br />
giusto????<br />
Compatibile con la cellula ospite<br />
Necessita di una buona combinazione di:<br />
Promotori forti<br />
ribosome binding sites<br />
Sequenze di terminazione<br />
affinity tag o sequenze per rendere le proteine solubili<br />
Siti di restrizione multi-enzyme
Clonaggio in lievito:<br />
Clonaggio e preparazione di plasmidi<br />
molto inefficiente (shuttle-vectors)<br />
Numero di copie di plasmidi per cellula: 1-<br />
50<br />
Può contenere più di un plasmide<br />
contemporaneamente<br />
Sistema di ricombinazione omologa<br />
affidabile ed efficiente
Vettori Shuttle<br />
Integrato, low copy, molto stabile<br />
High copy (20-40/cell),<br />
relativamente stabile<br />
High copy, instabile<br />
Low copy (2), stabile
Integrative plasmids (YIp):<br />
Backbone di un vettore<br />
di E. coli (pBR322,<br />
pUC19, pBLUESCRIPT)<br />
Marcatore di selezione<br />
per lievito (<strong>URA3</strong>, HIS3,<br />
TRP1, LEU2)<br />
Origine di replicazione per<br />
lievito: assente<br />
Propagati mediante<br />
integrazione genomica<br />
Vettori-shuttle Lievito-E. coli<br />
Amp-resistance<br />
Pst I (4795)<br />
Apa LI (3971)<br />
Apa LI (5217)<br />
PMB1<br />
Apa LI (3473)<br />
YIp5: pBR322 più gene <strong>URA3</strong><br />
YIp5<br />
5541bp<br />
Eco R I (2)<br />
Cla I (28)<br />
Xma I (2541)<br />
Sma I (2543)<br />
Hin d III (33)<br />
Bam H I (379)<br />
<strong>URA3</strong><br />
Ava I (2541)<br />
Tet-resistance<br />
Pst I (1644)<br />
Nco I (1867)
Integrazione del plasmide nel genoma di lievito:<br />
Ricombinazione omologa<br />
Duplicazione della regione target<br />
La linearizzazione del plasmide incrementa il tasso di<br />
ricombinazione<br />
Intergazione stabile<br />
X<br />
ura3<br />
X<br />
plasmide<br />
ura3<br />
plasmide<br />
<strong>URA3</strong><br />
<strong>URA3</strong><br />
genoma<br />
genoma
Costruzione di un null mutant
Vettori-shuttle Lievito-E. coli<br />
Plasmidi episomali (YEp):<br />
Backbone di u vettore<br />
di E. coli (pBR322,<br />
pUC19, pBLUESCRIPT)<br />
Marcatore di selezione<br />
per lievito (<strong>URA3</strong>, HIS3,<br />
TRP1, LEU2)<br />
Origine di replicazione del<br />
plasmide 2micron del<br />
lievito<br />
Propagati stabilmente<br />
high copy number ( 20-50<br />
per cellula)<br />
L’uso di marker ”difettosi”<br />
può spingere il numero di<br />
plasmidi fino a 200/cell<br />
PMB1<br />
Apa LI (5701)<br />
Amp-resistance<br />
Apa LI (6199)<br />
Pst I (7023)<br />
Ava I (4835)<br />
Apa LI (7445)<br />
Tet-resistance<br />
EcoR I (2)<br />
YEp24<br />
7769bp<br />
BamH I (3785)<br />
Hind III (106)<br />
<strong>URA3</strong><br />
Xma I (3379)<br />
Ava I (3379)<br />
Sma I (3381)<br />
Hind III (3439)<br />
2micron ORI<br />
Ava I (1391)<br />
Pst I (2001)<br />
EcoR I (2242)<br />
Cla I (2268)<br />
Hind III (2273)<br />
Pst I (2482)<br />
Nco I (2705)<br />
YEp24: pBR322 plus the <strong>URA3</strong> gene, plus 2micron origin
Replicative centromeric plasmids<br />
(YCp) :<br />
backbone di u vettore di E. coli<br />
(pBR322, pUC19, pBLUESCRIPT)<br />
Marcatore di selezione per lievito<br />
(<strong>URA3</strong>, HIS3, TRP1, LEU2)<br />
origine di replicazione cromosomale<br />
ARS (autonomously replicating<br />
sequence)<br />
centromero CEN di cromosoma di<br />
lievito<br />
Propagati stabilmente<br />
Basso numero di copie (1 per cellula)<br />
Vettori-shuttle Lievito-E. coli<br />
Amp-resistance<br />
Apa LI (6380)<br />
PMB1<br />
Apa LI (5882)<br />
Apa LI (5457)<br />
Pst I (5451)<br />
Pst I (7204)<br />
ARS1<br />
Ava I (4703)<br />
Apa LI (7626)<br />
CEN4<br />
EcoR I (2)<br />
Cla I (28)<br />
YCp50<br />
7950bp<br />
Hind III (33)<br />
BamH I (379)<br />
Tet-resistance<br />
POLY<br />
Pst I (1644)<br />
Nco I (1867)<br />
<strong>URA3</strong><br />
Xma I (2541)<br />
Ava I (2541)<br />
Sma I (2543)<br />
POLY<br />
YCp50: pBR322 plus the <strong>URA3</strong> gene, plus CEN4, plus ARS1
Delezione genica in lievito<br />
1) In vitro: realizzazione costrutto (gene marcatore affiancato da porzioni del<br />
gene da eliminare)<br />
2) In vivo: Inserimento del costrutto in lievito dove sostituisce il gene target<br />
3) Analisi genotipica e fenotipica per confermare la delezione<br />
in vitro<br />
in vivo<br />
GDI<br />
<strong>URA3</strong><br />
<strong>URA3</strong><br />
Gene di interesse<br />
X X<br />
<strong>URA3</strong><br />
Inserimento del GDI in un plasmide<br />
Ricombinazione omologa in lievito<br />
Delezione del GDI dal genoma
Delezione genica in lievito<br />
Come generare il costrutto per la delezione:<br />
(1)Digestione con enzimi di restrizione e successiva ligazione<br />
(2)PCR in due volte senza step di clonaggio<br />
(3)Unico step di PCR con primer lunghi
ResI<br />
ResI<br />
Delezione genica in lievito (1)<br />
Digestione con enzimi di restrizione e successiva ligazione<br />
<strong>URA3</strong><br />
<strong>URA3</strong><br />
ResII<br />
ResI<br />
ResII<br />
<strong>URA3</strong><br />
ResI<br />
ResII<br />
ResI<br />
GDI<br />
ResII<br />
GDI<br />
ResII
Delezione genica in lievito (2)<br />
√ PCR in due volte senza step di clonaggio<br />
GDI<br />
<strong>URA3</strong><br />
<strong>URA3</strong><br />
<strong>URA3</strong>
Delezione genica in lievito (3)<br />
√ Unico step di PCR con primer lunghi:<br />
<strong>URA3</strong>
Altri usi dell’integrazione genica (1)<br />
Inserimento nella cassetta di delezione di un gene reporter (lacZ) che<br />
lo pone sotto il controllo del promotore del GDI per studiarne<br />
l’espressione<br />
Usando tale costrutto in un diploide si può studiare l’espressione<br />
genica (monitorando l’attivita’ della beta–galattosidase) prima nel<br />
diploide in caso di un gene essenziale<br />
Promotore X<br />
lacZ<br />
GDI1<br />
<strong>URA3</strong><br />
Cellula diploide
Altri usi dell’integrazione genica (2)<br />
Taggaggio genico: Inserimento nella cassetta del GDI in frame con<br />
lacZ/ GFP/immuno-tag per ottenere una proteina di fusione<br />
facilmente rilevabile o purificabile.<br />
GDI<br />
GFP<br />
<strong>URA3</strong>
POP-out del marcatore<br />
Riutilizzare il marcatore per successive delezioni<br />
Eliminazione di tracce di DNA esogeno<br />
Up REPEAT <strong>URA3</strong> REPEAT Dw<br />
Up OLD Dw<br />
Terreno contente un composto che impedisce<br />
la crescita alle cellule contenenti il gene <strong>URA3</strong><br />
Terrene non contenente uracile<br />
OLD<br />
Up<br />
<strong>URA3</strong><br />
REPEAT<br />
REPEAT<br />
Up REPEAT Dw<br />
Dw
Applicazioni della ricombinazione<br />
omologa in lievito: clonaggio mediante<br />
gap-repair<br />
1) Cotrasformazione del gene di interesse con il plasmide<br />
2) Cotrasformazione di 2 geni di interesse con il plasmide<br />
3) Ricombinazione tra il plasmide scelto e genoma<br />
1<br />
X<br />
gapped plasmid<br />
GDI<br />
X<br />
repair fragment<br />
2<br />
gapped plasmid<br />
X<br />
YFG1 GDI<br />
X<br />
3<br />
gapped plasmid<br />
YFG1 GDI<br />
X X<br />
Copia genomica usata per riparare il gap;<br />
Il templato viene duplicato
Cloning Mutants by Gapped<br />
Plasmid Repair
Promotori: costitutivi<br />
ADH1 – alcohol dehydrogenase<br />
GPD – glyceraldehyde-3-phosphate dehydrogenase<br />
PMA1 – plasma membrane H + -ATPase<br />
PDR5 – pleiotropic drug resistant pump<br />
PMA1 & PDR5 –L’ espressione può raggiungere<br />
anche il 10% delle proteine della membrana<br />
plasmatica
Gal1<br />
Promotori: Inducibili<br />
CUP1 – metalothionein gene promoter<br />
Inducibile grazie all’aggiunta di ioni rame<br />
PHO 5 – indotto da basse concentrazioni di fosfato<br />
inorganico extracellulare<br />
HSE – tandem heat shock elements<br />
Indotto da incremento della temperatura fino a 37 o C
Gal1<br />
Promotori: Inducibili<br />
GAL genes – metabolismo del galattosio<br />
Inducibile dal galattosio<br />
Represso in presenza di glucosio<br />
Antimalarian drug
Pichia pastoris<br />
Similitudini con Saccharomyces cerevisiae:<br />
Facile da manipolare<br />
Più veloce, più semplice, meno costoso di altri sistemi<br />
eucariotici<br />
Vantaggi rispetto a Saccharmoyces cerevisiae:<br />
Livello di espressione di proteine eterologhe 10-100 volte<br />
SUPERIORE!!!!<br />
Pichia è un lievito metilotrofico (metabolizza metanolo)<br />
CH 3 OH<br />
AOX<br />
O 2 H 2 O 2<br />
HCHO
2 geni codificano per l’alcohol oxidase:<br />
AOX1e AOX2<br />
Pichia pastoris<br />
AOX1 principale responsabile dell’attività<br />
AOX1 inducibile con metanolo<br />
Può raggiungere fino al 30% delle proteine totali solubili in<br />
cellule cresciute con metanolo<br />
Controllato a livello trascrizionale<br />
Simile al promotore Gal1: Trascrizione repressa da glucosio,<br />
anche in presenza di metanolo
Ceppi di Pichia mutanti utilizzati nelle strategie produttive:<br />
Fenotipi :<br />
1) Mut + : AOX1 e AOX2 sono presenti e funzionali, crescono rapidamente su metanolo<br />
2) Mut s : “slow”, sono aox1- AOX2 più lenti (5-10% attività presente) (utilizzati, ad es, per<br />
produzione di antigeni contro epatite B)<br />
3) Mut - : mutanti aox1,2-, incapaci di crescere su metanolo. Crescono molto più<br />
lentamente, utili in alcuni casi nella produzione di proteine ricombinanti<br />
Sviluppati vettori soprattutto di integrazione (che avviene principalmente per<br />
ricombinazione omologa)<br />
Ottimizzato l’uso di promotori, soprattutto quelli della via di utilizzo del metanolo<br />
Possibilità di utilizzo di marcatori sia metabolici (principalmente HIS4) sia antibiotici<br />
(principalmente zeocina)<br />
Possibilità di produrre la proteina eterologa nel citoplasma oppure secreta nel mezzo. La<br />
sequenza “leader” pre-pro dell’alpha factor di S. cerevisiae viene utilizzata frequentemente<br />
per guidare la secrezione.<br />
Sviluppati ceppi deleti in proteasi, in associazione o meno con ottimizzazione di strategie<br />
di coltivazione
Vantaggi per P. pastoris<br />
Cresce fino a raggiungere densità cellulari estremamente<br />
elevate<br />
Intracellulare o Secrete<br />
Pichia secerne poche proteine native (0,5%)<br />
Poche proteasi nel mezzo<br />
Purificazione più semplice!!!<br />
Elevate rese di prodotto<br />
Esempio: Sea Raven anti-freeze protein<br />
40 mg/L (secreta)<br />
Pichia pastoris<br />
Esempio: Hepatitis B virus surface antigen<br />
Resa= 400 mg/L (intracellulare)
Produzione di proteine umane ricombinanti<br />
Problema: Glicosilazione inappropriata
Soluzione 1: eliminare il problema!!<br />
Insulina – proteina terapeutica espressa in E.coli e lievito<br />
Non richiede glicosilazione<br />
La maggior parte delle proteine umane richiedono la<br />
glicosilazione
Soluzione 2: Ingegnerizzare l’ospite<br />
Eliminare il pathway di glicosilazione endogeno<br />
(ceppo och1: delezione della alpha 1-6 mannosyltransferase)<br />
Introdurre il pathway dell’uomo attivo<br />
mannosidases I and II<br />
N-acetylglucosaminyl transferases I and II<br />
uridine 5'-diphosphate (UDP)-N-acetylglucosamine<br />
transporter.<br />
Verificare: Introduzione del gene reporter umano K3<br />
(ha un singolo sito di glicosilazione)
Isolati ceppi di lievito che<br />
mostrano alte rese di proteina<br />
reporter umana K3<br />
caratterizzata da glicosilazione<br />
human-like<br />
Hamilton, SR (2003) Production of Complex Human Glycoproteins in Yeast.<br />
Science 301: 1244 - 1246.<br />
Soluzione 2: Ingegnerizzare l’ospite.<br />
Questo lavoro descrive la “humanization” del pathway di glicosilazione di Pichia.
ANTIBODY DEPENDENT CELL CITOTOXICITY:<br />
3<br />
APPLICAZIONI: PRODUZIONE DI ANTICORPI IN<br />
PICHIA<br />
1<br />
2<br />
1) Il dominio variabile dell’anticorpo lega la cellula target<br />
2) La cellula killer NK lega la regione costante Fc dell’anticorpo<br />
1) La cellula killer distrugge la cellula target<br />
Problema: Anticorpi che non sono N-glicosilati<br />
possono legare l’antigene ma non mediano l’ADCC<br />
Utilizzo di ceppi di Pichia pastoris<br />
“glicoengineered”<br />
per produrre anticorpi funzionanti nell’uomo