Lezioni 4 - 8
Lezioni 4 - 8 Lezioni 4 - 8
- Page 6 and 7: La sequenza degli eventi elettrofis
- Page 14 and 15: Fase 2: il rilascio La depolarizzaz
- Page 16 and 17: In che modo il Calcio determina la
- Page 19: L’azione dei neurotrasmettitori a
- Page 30 and 31: Fase 3: il cambiamento del potenzia
- Page 32 and 33: La trasmissione sinaptica eccitator
- Page 34 and 35: Il risultato finale di tutta l’at
- Page 36 and 37: Recettori a secondo messaggero: glu
- Page 38 and 39: La trasmissione sinaptica a secondo
- Page 40 and 41: Come dimostrare che una sostanza è
- Page 42 and 43: Il glutammato lega recettori-canale
- Page 44 and 45: I recettori non-NMDA (che si suddiv
- Page 46 and 47: La neuromodulazione Le trasmissioni
- Page 48 and 49: Un esempio di neuromodulazione: la
- Page 50: I sistemi a proiezione diffusa Sist
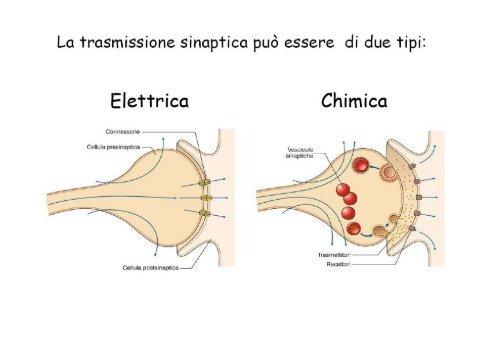
La sequenza degli eventi elettrofisiologici alla base della<br />
trasmissione sinaptica. Il ritardo sinaptico.<br />
Fatto …<br />
Le correnti Na +<br />
e K + NON<br />
sono<br />
necessarie per<br />
il rilascio del Nt
Esperimento Katz no Calcio extracellulare
Fase 2: il rilascio<br />
La depolarizzazione del terminale presinaptico causata<br />
dal potenziale di azione apre i canali calcio voltaggiodipendenti.<br />
Lo ione calcio (Ca++) ha un potenziale di equilibrio<br />
positivo e ha quindi una f.e.m. che lo spinge ad entrare<br />
nel terminale.<br />
Una volta entrato, il Ca lega delle proteine presenti sulla<br />
superficie delle vescicole contenenti il neurotrasmettitore<br />
(vescicole sinaptiche) determinando la fusione delle<br />
vescicole con la membrana cellulare e la liberazione nel<br />
vallo sinaptico del neurotrasmettitore contenuto nelle<br />
vescicole.
Per minimizzare i tempi di questo processo le<br />
vescicole sono già giustapposte alla membrana<br />
prima dell’arrivo del potenziale di azione. Inoltre<br />
le vescicole sono radunate in una zona del<br />
terminale, la zona attiva, in cui si trovano<br />
concentrati i canali calcio voltaggio-dipendenti.<br />
In questo modo il Ca che entra trova<br />
immediatamente il suo bersaglio sulla vescicola<br />
sinaptica e la vescicola può immediatamente<br />
completare la fusione e rilasciare il<br />
neurotrasmettitore.
In che modo il Calcio determina la fusione delle<br />
vescicole??<br />
V-SNARE<br />
Interazioni con il complesso SNARE<br />
Indicazioni da esperimenti con tossine<br />
clostridiche (botulino e tetano)<br />
T-SNARE<br />
+ Ca 2+
Affinchè la sinapsi sia pronta a rispondere a nuovi stimoli,<br />
una fase cruciale della trasmissione sinaptica è la<br />
rimozione dei neurotrasmettitori subito dopo il loro<br />
rilascio. Ciò può avvenire secondo tre meccanismi:<br />
diffusione, degradazione e riassunzione.<br />
La diffusione permette l’allontanamento del<br />
neurotrasmettitore dal sito di rilascio; la degradazione è<br />
utilizzata solo per alcuni neurotrasmettitori (ad es.<br />
l’acetilcolina) ed è eseguita da enzimi specifici posti<br />
extracellularmente tra il versante presinaptico e quello<br />
postsinaptico; la riassunzione del neurotrasmettitore è un<br />
meccanismo abbastanza comune tra i diversi<br />
neurotrasmettitori ed avviene tramite proteine trasportatrici<br />
poste sulla membrana del neurone o delle cellule gliali che<br />
internalizzano i neurotrasmettitori rilasciati.
L’azione dei neurotrasmettitori a livello postsinaptico è<br />
stata inizialmente investigata a livello della sinapsi<br />
neuromuscolare.<br />
Si è osservato che l’acetilcolina lega selettivamente<br />
recettori sinaptici, i recettori per l’acetilcolina di tipo<br />
nicotinico, che sono localizzati sulla membrana<br />
postsinaptica.<br />
Oltre a legare il neurotrasmettitore questi recettori<br />
formano anche un canale e fanno quindi parte della<br />
categoria dei canali ligando-dipendenti.
Per studiare quali ioni attraversino i recettori-canale dell’acetilcolina,<br />
sono state misurate le correnti sinaptiche evocate nel terminale<br />
postsinaptico dalla stimolazione degli assoni presinaptici imponendo<br />
sperimentalmente diversi valori del potenziale di membrana alla cellula<br />
postsinaptica.<br />
Individuare a quale potenziale di membrana le correnti ioniche che<br />
attraversano i recettori-canali si azzerano dà un’importante informazione<br />
in quanto, a seconda del suo valore, indica se la corrente sinaptica è<br />
mediata dal sodio, dal potassio o da entrambi.<br />
Gli esperimenti hanno mostrato che la corrente sinaptica che attraversa<br />
i recettori nicotinici per l’acetilcolina diventa nulla se si impone un<br />
potenziale di membrana di 0 mV. Questo potenziale viene chiamato<br />
potenziale di inversione.<br />
Un potenziale di inversione di 0 mV non corrisponde né al potenziale di<br />
equilibrio del potassio E K né a quello del sodio E Na . In effetti, i recettori<br />
nicotinici per l’acetilcolina hanno permeabilità mista sodio-potassio.
Fase 3: il cambiamento del potenziale di membrana<br />
postsinaptica<br />
La trasmissione sinaptica eccitatoria<br />
Si definisce eccitatoria una sinapsi la cui attivazione<br />
aumenta la probabilità che il neurone postsinapico<br />
produca potenziali di azione.<br />
Il risultato di una trasmissione sinaptica eccitatoria nella<br />
maggior parte dei casi è una depolarizzazione del<br />
terminale postsinaptico (EPSP). Nel sistema nervoso<br />
centrale la gran parte delle sinapsi eccitatorie utilizza il<br />
glutammato mentre alla sinapsi neuromuscolare viene<br />
utilizzata l’acetilcolina.
Oltre a suggerire quali ioni formino la corrente sinaptica, il fatto<br />
che il potenziale di inversione sia 0 mV ci indica anche la natura<br />
eccitatoria della sinapsi neuromuscolare. Difatti, se il potenziale di<br />
membrana fosse minore di 0 mV, la f.e.m. spingerebbe le cariche<br />
positive che attraversano i canali aperti dall’acetilcolina ad entrare<br />
fino a che Vm non raggiungesse il valore del potenziale di<br />
inversione. A quel valore la f.e.m. si annullerebbe e di<br />
conseguenza si annullerebbe anche la corrente sinaptica.<br />
Ciò significa che l’attivazione di questa sinapsi tende a portare il<br />
potenziale di membrana a 0 mV, un valore molto superiore al<br />
valore soglia per l’innesco del potenziale di azione e, in definitiva,<br />
ad aumentare la probabilità che la cellula postsinaptica generi un<br />
potenziale di azione.<br />
È quindi evidente che l’azione eccitatoria o inibitoria di un<br />
neurotrasmettitore dipende dal tipo di recettore sinaptico cui<br />
esso si lega ed in particolare dal potenziale di inversione<br />
della corrente sinaptica che da esso è determinata.
La<br />
trasmissione<br />
sinaptica<br />
eccitatoria:
La trasmissione sinaptica inibitoria<br />
Il GABA è il più importante<br />
neurotrasmettitore inibitorio nel<br />
cervello<br />
La corrente inibitoria postsinaptica<br />
(IPSC) che media gli<br />
IPSP inverte a -70mV e cioe’<br />
intorno al potenziale di riposo<br />
del Cloro
Il risultato finale di tutta l’attivita’ sinaptica inibitoria ed eccitatoria<br />
e’ rappresentata dalla frequenza di scarica in uscita lungo<br />
l’assone.<br />
L’inibizione modula la scarica impedendo l’instaurarsi di potenziali<br />
di azione o diminuendo la probabilità che vengano generati.<br />
Eccitatorio: avvicina V m alla soglia per la generazione di un<br />
potenziale di azione<br />
Inibitorio: diminuisce la probabilita’che si superi la soglia per la<br />
generazione di un potenziale di azione.
La localizzazione preferenzialmente somatica delle sinapsi<br />
inibitorie favorisce la loro azione di blocco dell’azione di<br />
sinapsi eccitatorie simultaneamente attive.
Recettori a secondo messaggero: glutamato
La trasmissione sinaptica indiretta (o a secondo messaggero)<br />
Trasmettitori:<br />
GABA, glicina<br />
Glutammato<br />
Acetilcolina (nicotinici)<br />
Trasmettitori:<br />
GABA (GABAB)<br />
Glutammato (metabotropici)<br />
Acetilcolina (muscarinici)<br />
Noradrenalina<br />
Dopamina<br />
Serotonina<br />
Opioidi ed altri….
La trasmissione sinaptica a secondo messaggero segue uno schema stereotipato
Esempio: l’azione della serotonina<br />
su neuroni di invertebrato.<br />
L’eccitazione è indiretta e mediata<br />
dalla chiusura di canali Potassio
Come dimostrare che una sostanza è un<br />
neurotrasmettitore??
I neurotrasemttitori: acetilcolina
Il glutammato lega recettori-canaledi tipo NMDA e di tipo non-<br />
NMDA; questi due tipi di recettori, pur legando entrambi il<br />
glutammato, sono distinguibili sulla base della capacità di legarsi<br />
o meno al composto NMDA.<br />
A questa differenza farmacologica corrispondono più importanti<br />
differenze funzionali.<br />
I recettori non-NMDA hanno permeabilità ionica e potenziale di<br />
inversione analoghi a quelli descritti per i recettori nicotinici per<br />
l’acetilcolina; i recettori NMDA invece mostrano delle importanti<br />
peculiarità.<br />
L’apertura del recettore canale NMDA avviene solo se<br />
l’azione del glutammato coincide con la depolarizzazione del<br />
neurone postsinaptico. Questo recettore-canale è quindi un<br />
esempio pressoché unico di canale sia ligando-dipendente<br />
che voltaggio-dipendente ed è un perfetto detettore della<br />
correlazione fra l’attività del neurone pre e postsinaptico.
Trasmissione sinaptica glutamatergica: recettori NMDA e non-<br />
NMDA<br />
La trasmissione sinaptica eccitatoria nel SNC ha come principale trasmettitore il<br />
glutammato.<br />
I recettori postsinaptici per il glutammato ionotropici (recettori-canali) possono<br />
appartenere a due tipi diversi
I recettori non-NMDA (che si suddividono in AMPA e kainato a seconda<br />
della loro affinita’ per queste due molecole) sono bloccati dall acido<br />
quisqualico.<br />
I recettori NMDA sono bloccati dall’acido fosfoaminovalerico (APV).<br />
L’APV blocca una componente tardiva, V-dipendente della corrente<br />
sinaptica<br />
Se rimuovo il Magnesio extracellulare il recettore canale NMDa non e’ piu’ Vdipendente
Il recettore NMDA come detettore di correlazione fra l’attività dell’elemento<br />
presinaptico e dell’elemento postsinaptico<br />
L’Ipotesi di Hebb
La neuromodulazione<br />
Le trasmissioni sinaptiche a secondo messaggero<br />
influenzano l’eccitabilità della cellula postsinaptica<br />
causando l’attivazione di cascate biochimiche che<br />
regolano l’apertura o la chiusura di canali posti su<br />
tutta la membrana postsinaptica.<br />
Alcuni neurotrasmettitori possiedono solo recettori a<br />
secondo messaggero.<br />
L’attività di questi neurotrasmettittori<br />
(neuromodulatori) è poco evidente per se, ma si<br />
evidenzia con una facilitazione o depressione delle<br />
risposte del neurone postsinaptico ad altri ingressi.<br />
Questa è la neuromodulazione.
Es. di neuromodulazione: L’azione degli oppioidi sulla trasmissione del<br />
dolore è mediata da sinapsi che modulano l’efficaci della sinapsi tra la fibra<br />
nocicettiva afferente e il neurone che trasmette l’informazione dolorifica<br />
centralmente.
Un esempio di<br />
neuromodulazione:<br />
la noradrenalina<br />
rimuove<br />
l’adattamento<br />
chiudendo i canali<br />
potassio calcio<br />
dipendenti nei<br />
neuroni<br />
dell’ippocampo
Bersaglio privilegiato della trasmissione sinaptica<br />
a secondo messaggero (e quindi della<br />
neuromodulazione) sono i canali Potassio<br />
Passivi (che succede?)<br />
V-dip (che succede?)<br />
Ca 2+ dipendenti (che succede?)<br />
ed i canali Ca 2+ (modulazione presinaptica) (che<br />
succede?)
I sistemi a proiezione diffusa<br />
Sistema noradrenergico<br />
Sistema dopaminergico
I recettori dei neuromodulatori sono anche bersaglio di farmaci o di<br />
sostanze d’abuso:<br />
Serotonina: Prozac, LSD<br />
Dopamina: antipsicotici (clorpromazina, clozapina, spiperone, antipsicotici<br />
di nuova generazione, SSRI)<br />
Dopamina-noradrenalina: cocaina, amfetamina<br />
Noradrenalina-dopamina-serotonina: antidepressivi (triciclici)<br />
Sistema serotoninergico<br />
Sistema colinergico
La sinapsi dopaminergica è un sito di azione per la modulazione delle psicosi
La concentrazione a<br />
cui diversi farmaci<br />
bloccano il recettore<br />
D2 della Dopamina<br />
correla con i loro<br />
dosaggi efficaci<br />
nell’ottenere l’azione<br />
antipsicotica nel<br />
paziente.<br />
Per gli antipsicotici<br />
atipici (amisulpride,<br />
aripiprazolo,<br />
clozapina, olanzapina,<br />
quetiapina, risperidone<br />
e zotepina), va tenuto<br />
conto anche delle loro<br />
interazioni con il<br />
sistema<br />
serotoninergico.<br />
Anche lo spiperone<br />
interagisce con<br />
recettori<br />
serotoninergici.
Integrazione dei segnali nervosi<br />
Ricostruiamo un motoneurone del midollo spinale:<br />
Riceve piu’ di 10000 sinapsi mediate da neurotrasmettitori diversi! Ma ha una singola<br />
uscita:<br />
i potenziali d’azione che possono variare in frequenza ma non in ampiezza
E’ quindi molto importante capire come<br />
l’attività delle varie sinapsi si integrino.<br />
Le proprieta’ passive sono importanti<br />
e quindi la<br />
localizzazione<br />
delle sinapsi ,<br />
che si<br />
raggiunge<br />
durante lo<br />
sviluppo, e’<br />
importante
La plasticità sinaptica
I circuiti nervosi possono modificarsi in risposta all’esperienza<br />
Questa capacità si definisce plasticità neurale<br />
La plasticità’ è presente in tutto il SNC, ma in particolar modo a livello<br />
della corteccia cerebrale<br />
Senza di essa<br />
•Il nostro cervello non si sarebbe sviluppato in maniera<br />
normale<br />
•Il nostro comportamento sarebbe stereotipato ed<br />
immodificabile dall’esperienza<br />
•Saremmo esseri senza memoria del passato
I cambiamenti dei circuiti neurali in risposta all’esperienza<br />
sono meno evidenti man mano che la plasticità corticale<br />
diminuisce con il procedere dello sviluppo<br />
Tuttavia, la plasticità corticale è presente nell’adulto, sia in<br />
risposta ad aumenti dell’attività afferente (apprendimenti) sia in<br />
risposta a riduzioni dell’attività afferente (lesioni)
In seguito ad una lesione al sistema nervoso centrale si<br />
verifica una riorganizzazione dei circuiti neurali,<br />
particolarmente evidente a livello corticale.<br />
Tale riorganizzazione può condurre, nel caso di lesioni che<br />
avvengono durante lo sviluppo, ad un recupero della<br />
funzione danneggiata.<br />
• Riorganizzazione perilesionale<br />
• Riorganizzazione con spostamento all’emisfero<br />
controlaterale<br />
Il caso di lesioni alle aree dell’emisfero sinistro e lo<br />
sviluppo del linguaggio:<br />
• Il “costo” della plasticità<br />
• La “non equipotenzialità” degli emisferi
Comprendere i meccanismi alla base della<br />
plasticità corticale, sia durante lo sviluppo<br />
che nell’adulto, potrebbe permettere di:<br />
correggere difetti insorti in seguito ad un<br />
processo di sviluppo deficitario o scorretto<br />
sfruttare al meglio o potenziare i tentativi<br />
spontanei di recupero che il sistema nervoso<br />
mette in atto dopo una lesione
Le “regole” della plasticità<br />
Raffinamento e maturazione dei circuiti neurali durante<br />
lo sviluppo<br />
Formazione di una traccia di memoria
Plasticità neurale: capacità dei circuiti nervosi di modificarsi,<br />
cambiando quindi la loro uscita, in risposta all’esperienza<br />
Cosa cambia nel circuito?<br />
L’efficacia delle connessioni sinaptiche si modifica in risposta ai<br />
cambiamenti nell’attività elettrica che l’esperienza induce nel<br />
circuito<br />
Ingresso<br />
presinaptico<br />
Risposta<br />
postsinaptica<br />
Esperienza<br />
(cambio di attività mantenuto nel tempo)<br />
Ingresso<br />
presinaptico<br />
Risposta<br />
postsinaptica<br />
o
Meccanismo Hebbiano di plasticità:<br />
rafforzamento delle sinapsi con attivita’<br />
sincrona<br />
Principio di Hebb: se un neurone presinaptico A attiva<br />
ripetutamente il neurone postsinaptico B la connessione sinaptica<br />
fra A e B si rafforza (1949)<br />
Corollario: se A fallisce ripetutamente nell’attivare B, la<br />
connessione fra A e B si indebolisce
Neuroni presinaptici<br />
attivi sincronicamente<br />
Neuroni presinaptici<br />
attivi asincronamente<br />
Meccanismo Hebbiano di plasticità:<br />
rafforzamento delle sinapsi con attivita’ sincrona<br />
Attività del neurone<br />
postsinaptico
La verifica sperimentale dell’ipotesi di Hebb si ebbe con la scoperta, all’inizio degli<br />
anni ’70, del fenomeno del potenziamento a lungo termine (LTP).<br />
Esempio di potenziamento<br />
a lungo termine. Uno<br />
stimolo ad alta frequenza<br />
(treno di impulsi, indicato<br />
con il triangolo nero)<br />
produce un incremento<br />
transitorio della risposta<br />
sinaptica (LTP precoce).<br />
Se si eseguono 4<br />
stimolazioni ad alta<br />
frequenza si ha un<br />
potenziamento maggiore e<br />
permanente nel tempo<br />
(LTP tardivo). Tratto da<br />
Kandel et al., 2003.<br />
Monitorando le ampiezze degli EPSP ci si accorse che la risposta a stimoli test<br />
somministrati a bassa frequenza (uno ogni 15-60 secondi) aumentava se la sinapsi<br />
veniva sottoposta ad un breve episodio di stimolazione ad alta frequenza. Se<br />
venivano somministrate ripetute stimolazioni ad alta frequenza, il potenziamento era<br />
più ampio e permanente nel tempo. Questi fenomeni sono presenti sia in vitro che in<br />
vivo e sono stati osservati in molte aree cerebrali.
Esperimenti successivi hanno mostrato che LTP della<br />
trasmissione sinaptica in una via afferente veniva indotto anche<br />
con una stimolazione a bassa frequenza, a patto che essa fosse<br />
accoppiata ad una forte depolarizzazione della cellula<br />
postsinaptica indotta dallo sperimentatore.<br />
Questo esperimento verificava direttamente l’ipotesi di Hebb ed<br />
identificava tre importanti proprietà del LTP:<br />
la specificità del LTP per la via attivata, infatti la depolarizzazione<br />
della cellula postsinaptica per se non induceva LTP nelle vie che<br />
non erano state attive in coincidenza con la depolarizzazione;<br />
l’associatività, per cui se una via era debolmente attiva in modo<br />
però sincrono con una via fortemente attiva, andava incontro ad<br />
egualmente ad LTP in quanto si realizzava una coincidenza<br />
temporale tra il suo stato di attivazione e una forte attivazione<br />
postsinaptica;<br />
la cooperatività, per cui l’attivazione simultanea di numerose fibre<br />
presinaptiche è necessaria per elicitare LTP.
Ingresso<br />
presinaptico<br />
Risposta<br />
postsinaptica<br />
Ingresso<br />
presinaptico<br />
Risposta<br />
postsinaptica<br />
Ingresso<br />
presinaptico<br />
Risposta<br />
postsinaptica<br />
Meccanismi di plasticità sinaptica<br />
(non alternativi)<br />
Potenziamento<br />
sinaptico<br />
Nuovi siti<br />
sinaptici<br />
Crescita di nuove<br />
connessioni
Le modificazioni dell’efficacia sinaptica, per<br />
consolidarsi e mantenersi stabili nel tempo<br />
(modifiche a lungo termine) necessitano della<br />
trascrizione genica e della sintesi di nuove<br />
proteine
L’attività neuronale (=esperienza) modifica l’espressione genica<br />
I secondi<br />
messaggeri<br />
accoppiano gli<br />
ingressi<br />
neuronali<br />
all’espressione<br />
genica: le<br />
esperienze si<br />
traducono in<br />
alterazioni<br />
biochimiche e<br />
strutturali dei<br />
neuroni
Da qui 11 marzo 2008
L’accoppiamento attività neuronale – espressione<br />
genica è presente anche per la trasmissione sinaptica<br />
diretta ed è mediato dal calcio il quale può entrare nel<br />
neurone o attraverso i recettori per il glutammato di<br />
tipo NMDA oppure attraverso dei canali per il calcio<br />
voltaggio-dipendenti che si aprono quando il recettore<br />
non-NMDA depolarizza il neurone.<br />
Il calcio lega diversi enzimi intracellulari (proteine<br />
chinasi) che a loro volta possono traslocare al nucleo<br />
e modulare l’azione dei complessi proteici che<br />
determinano se un gene viene trascritto o meno.
I passi che conducono ad una modifica duratura dei<br />
circuiti neurali in risposta all’esperienza<br />
coinvolgono diversi fattori<br />
Il recettore NMDA funge da detettore di attività correlata fra il terminale presinaptico ed il<br />
terminale postsinaptico (blocco NMDA=non si induce plasticità)<br />
Livelli di inibizione<br />
Protein chinasi (ERK, PKA)<br />
Fattori di trascrizione (CREB)<br />
Rimodellamento della cromatina<br />
Fattori neurotrofici (NGF, BDNF)<br />
Neuromodulatori (Acetilcolina, Noradrenalina)<br />
Componenti della matrice extracellulare e<br />
proteasi che li degradano
a<br />
Normal<br />
Open and<br />
closed eye<br />
Electrical<br />
activity<br />
Axonal<br />
terminals<br />
Cortical neuron<br />
otput<br />
Cortical neuron<br />
Dendritic spine<br />
b c<br />
Rapid,<br />
functional<br />
effects<br />
Classical view Current view<br />
MD<br />
Slow<br />
morphological<br />
effects<br />
Rapid<br />
functional<br />
AND<br />
morphological<br />
effects<br />
at cortical level<br />
Late rearrangement of<br />
LGN axonal arbors<br />
MD
O<br />
• Percorso tipico dei primi giorni<br />
del test, strategia di girare in torno<br />
al bordo della vasca cercando una<br />
via di fuga.<br />
N<br />
La memoria spaziale nei roditori<br />
MORRIS WATER MAZE.<br />
E<br />
S<br />
S<br />
START START<br />
O<br />
N<br />
• Percorso tipico degli ultimi giorni<br />
del test<br />
E
Latenza (sec)<br />
Piattaforma<br />
sommersa<br />
60<br />
50<br />
40<br />
30<br />
20<br />
10<br />
0<br />
La memoria spaziale nei roditori<br />
Apprendimento Probe test<br />
1 2 3 4 5 6 7 8 9<br />
Giorni di allenamento<br />
Tempo nel quadrante (sec)<br />
30<br />
25<br />
20<br />
15<br />
10<br />
5<br />
0<br />
NO NE SO SE
Per la memoria spaziale, si ritiene che cellule<br />
specializzate, dette “place cells” dell’ippocampo<br />
formino “campi di posizione” che corrispondono a<br />
precise locazioni dell’individuo nello spazio,<br />
determinando la memorizzazione di una mappa<br />
spaziale.
Collaterali di<br />
Schaffer<br />
CA3<br />
Via delle fibre<br />
muscoidi<br />
CA1<br />
Giro dentato<br />
Via perforante
Al sistema di tracciamento della<br />
posizione dell’animale<br />
Al sistema di registrazione<br />
dell’attività neurale
La memoria spaziale è deficitaria in soggetti deficitari del recettore NMDA<br />
nel campo CA1 dell’ippocampo<br />
Soggetto normale Soggetto senza recettori NMDA nel CA1<br />
Soggetti normali (a sinistra) imparano la posizione della piattaforma e, quando essa<br />
viene rimossa nel probe test, alla fine dell’apprendimento, la cercano attivamente,<br />
esplorando la posizione in cui si trovava, che era nel quadrante in alto a sinistra (la<br />
linea tratteggiata indica il percorso compiuto dall’animale). Soggetti con delezione<br />
del recettore NMDA nel campo CA1 invece non ricordano dove era la piattaforma e<br />
cercano su tutta la superficie della piscina.
LTP ippocampale è assente in soggetti deficitari del<br />
recettore NMDA nel campo CA1 dell’ippocampo<br />
EPSP amplitude/EPSP baseline<br />
4<br />
2<br />
1<br />
tempo<br />
Normale<br />
KO<br />
Linea base
Effetti di un deficit di recettori NMDA sulla formazione di una mappa<br />
spaziale stabile<br />
Esplorazione<br />
Soggetto normale<br />
Ri- esplorazione<br />
I campi recettivi di place cells diverse, sviluppati durante 10 minuti di esplorazione il giorno prima in<br />
un soggetto wild type, sono sempre presenti e stabili. Cellule con campi recettivi sovrapposti,<br />
indicate dalla freccia, scaricano sincronicamente. L’animale ricorda di aver già visitato questo<br />
ambiente e ne possiede una mappa spaziale.<br />
Soggetto con delezione dei recettori NMDA nell’ippocampo<br />
I campi recettivi di place cells diverse, sviluppati durante 10 minuti di esplorazione in soggetti con<br />
delezione dei recettori NMDA nel campo CA1 dell’ippocampo sono più grandi del normale. Il giorno<br />
dopo mostrano tracce di instabilità, ovvero si sono spostati. Cellule con campi recettivi sovrapposti,<br />
indicate dalla freccia, NON scaricano sincronicamente. E’ come se l’animale entrasse in un ambiente<br />
nuovo, mai visitato prima.
Arena di esplorazione<br />
2<br />
1<br />
3<br />
4<br />
Fase 2: sonno a onde lente<br />
Fase 1: esplorazione<br />
2<br />
3<br />
1<br />
2<br />
3<br />
1<br />
4<br />
4 Wilson e<br />
McNaughton<br />
1999)


