


Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
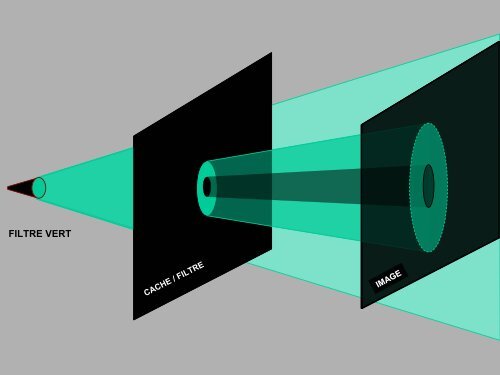
IMAGE IMAGE IMAGE<br />
CACHE CACHE / / FILTRE FILTRE<br />
FILTRE VERT
Dans un système linéaire, mesurer<br />
l’amplitude du signal de sortie du système<br />
pour une amplitude d’entrée constante<br />
(l’approche de l’ingénieur) est équivalent à<br />
mesurer l’amplitude entrante requise afin<br />
d’obtenir un signal de sortie constant<br />
(l’approche du psychophysicien).
LUMINANCE<br />
BANDE DE MACH<br />
ESPACE<br />
BRILLANCE
L’INHIBITION LATERALE<br />
Hartline, H.K., Wagne, H.G. & Ratliff, F.<br />
(1956). INHIBITION IN THE EYE OF<br />
LIMULUS. Journal of General Physiology 39,<br />
651-673.
BRILLANCE<br />
LUMINANCE<br />
One version of the Craik-O’Brien-Cornsweet effect.
An illusion by Vasarely, left, and a<br />
bandpass filtered version, right.
The simultaneous contrast effect.
Un ‘seul et même’ 1 CR pourrait donc<br />
expliquer la fonction de transfert de<br />
contraste humaine…<br />
__________<br />
1C.-à-d. une seule taile de CR.
FONCTION DE TRANSFERT DE<br />
MODULATION<br />
et<br />
REPONSE IMPULSIONNELLE
Inhibition centre-pourtour.
Dans un système linéaire et rétinotopique, la représentation d’un ensemble de points / image par<br />
un seul neurone est strictement identique à la représentation d’un point dans l’espace physique<br />
par l’ensemble des neurones qui le traitent.
CHAMP RECEPTEUR<br />
REPONSE IMPULSIONELLE
( ) = ΣE( l − k) • CR(k)<br />
S k<br />
l=+∞<br />
l=−∞<br />
( ) = ΣE() l • CR(l − k)<br />
S k<br />
l=+∞<br />
l=−∞
CONVOLUTION<br />
1 1 1 1 2 3 4 5 5 5 5<br />
-1 3 -1<br />
Σ<br />
Champ récepteur<br />
( ) E ∗h<br />
= ∫ +∞ −∞<br />
S X<br />
= E(X − x) • h(x) dx<br />
1 1 0 2 3 4 6 5 5<br />
E(X) = Entrée (fct. de X)<br />
S(X) = Sortie (fct. de X)<br />
CR = h(x) = Réponse Implle (fct. de x)<br />
Σ<br />
1 1 1 1 2 3 4 5 5 5 5<br />
-1 3 -1<br />
Réponse impulsionnelle<br />
3 -1<br />
-1 3 -1<br />
-1 3 -1<br />
-1 3 -1<br />
-2 6 -2<br />
-3 9 -3<br />
-4 12 -4<br />
-5 15 -5<br />
-5 15 -5<br />
E ∗h<br />
= E(X) • h(X − x)<br />
-5 15 -5<br />
-5 15<br />
( ) ∫ +∞ −∞<br />
S X<br />
= dx<br />
1 1 0 2 3 4 6 5 5
Campbell & Robson, 1968
Campbell & Robson, 1968
Campbell & Robson, 1968<br />
4 ⎡ 1 1<br />
⎤<br />
I(fx) carré = 1+ C<br />
⎢<br />
sin( π fx) + sin(3π fx) + sin(5π fx) + L<br />
π ⎣ 3 5<br />
⎥<br />
⎦<br />
Réponse / sensibilité prédite pour un réseau carré de fréquence fx et de<br />
contraste C :<br />
4 ⎡<br />
1 1<br />
⎤<br />
R(fx) carré = 1+ C<br />
⎢<br />
Rfx sin( π fx) + R3fx sin(3π fx) + R5fx<br />
sin(5π fx) + L<br />
3 5<br />
⎥<br />
sin sin sin<br />
π ⎣ ⎦
Campbell & Robson, 1968<br />
Mesuré<br />
Prédit<br />
4/π
Campbell & Robson, 1968
Il y aurait donc plusieurs tailles<br />
de CRs ou, ce qui revient au<br />
même, plusieurs filtres centrés<br />
sur plusieurs fréquences<br />
− + − − + −
1969<br />
1969
Wilson, H.R & Bergen, J.R. (1979). A<br />
four mechanisms model for spatial<br />
vision. Vision Research 19, 19-32.
REPRESENTATION ‘MULTIECHELLE’ DE CHAQUE POINT RETINIEN<br />
FOVEA<br />
Excentricité
Un « canal » d’Orientation…
Les « canaux » / filtres rétiniens dans le domaine<br />
chromatique (longueur d’onde) - normalisés
Les « canaux » / filtres<br />
rétiniens dans le domaine<br />
chromatique (longueur d’onde)<br />
– non-normalisés
Description of the ideal observer in a 2-feature trial in Experiment 2. First the ideal observer computes the<br />
ideal template, which is the difference between the target and the distractor. Then the ideal observer<br />
cross-correlates (takes the dot product)* this difference with the stimuli at each location. The location with<br />
the maximum dot product value is the stimulus that best matches the template, and is chosen as the<br />
target location.<br />
_______________________<br />
*x i y j [template] x i y j [stimulus]<br />
Shimozaki, S.S., Eckstein, M.P. & Abbey, C.K. (2002). Stimulus information contaminates summation<br />
tests of independent neural representations of features. Journal of Vision, 2, 354-370.
Schematic of the linear summation model. A 2-feature search from Experiment 2 is depicted with the<br />
target in the left locations. An independent response is generated for both spatial frequency and<br />
orientation at each location. These responses are weighted by the d’ for the particular feature and<br />
summed. The location with the maximal weighted linear response (xt-linear or xd-linear) is chosen as the<br />
target location.<br />
Shimozaki, S.S., Eckstein, M.P. & Abbey, C.K. (2002). Stimulus information contaminates summation<br />
tests of independent neural representations of features. Journal of Vision, 2, 354-370.
Feature 2<br />
Geometric description of linear summation. Each axis represents the sensitivity to each feature in d’ units.<br />
The sensitivity when both feature cues are available is equal to the Euclidean vector sum of the individual<br />
sensitivities. The orthogonal axes represent independence of the two features.<br />
Shimozaki, S.S., Eckstein, M.P. & Abbey, C.K. (2002). Stimulus information contaminates summation<br />
tests of independent neural representations of features. Journal of Vision, 2, 354-370.
Schematic of the probability summation model. A 2-feature search from Experiment 2 is depicted with the<br />
target in the left locations. An independent response is generated for both spatial frequency and<br />
orientation at each location, and the location with the maximal featural response is chosen as the target<br />
location.<br />
Shimozaki, S.S., Eckstein, M.P. & Abbey, C.K. (2002). Stimulus information contaminates summation<br />
tests of independent neural representations of features. Journal of Vision, 2, 354-370.
CANAUX, BRUIT<br />
et<br />
LA THÉORIE DE LA DÉTECTION DU SIGNAL
Le principe de l’UNIVARIANCE<br />
Réponse (s/s ou autre)<br />
Intensité ou « qualité » du Stimulus<br />
R<br />
Intensité<br />
«Qualité»(λ)<br />
V
RESPONSE NORMALIZATION<br />
Heeger, D.J. (1992) Normalization of cell responses in cat striate cortex. Visual Neuroscience, 9, 181-197.
LE MODÉLE STANDARD : LA THÉORIE DE LA DÉTECTION DU SIGNAL<br />
RESPONSE<br />
L<br />
RESPONSE<br />
L<br />
RESPONSE<br />
λ<br />
λ<br />
λ<br />
R<br />
«COULEUR»<br />
V<br />
R<br />
«COULEUR»<br />
V<br />
ΔR<br />
ΔR<br />
PROBAB.<br />
σ<br />
PROBAB.<br />
σ<br />
CONTINUUM SENSORIEL (R-V)<br />
CONTINUUM SENSORIEL (R-V)<br />
d’ n’est pas mesurable<br />
d’= ΔR σ
CANAUX & PROBABILITÉ DE RÉPONSE<br />
LE PLUS RÉPONDANT GAGNE<br />
RÉPONSE<br />
p Max<br />
p min<br />
REPONSE (e.g. s/s)<br />
R V<br />
p(s/s)<br />
p min<br />
x p max<br />
pmin x p max<br />
R V<br />
p[R(Y/G) largest]
CANAUX & PROBABILITÉ DE RÉPONSE<br />
« OFF-LOOKING » CHANNEL<br />
RÉPONSE<br />
REPONSE (e.g. s/s)<br />
R<br />
V<br />
p(s/s)<br />
REPONSE (e.g. s/s)<br />
R<br />
V<br />
p(s/s)





