UNIVERSITÉ PAUL CÉZANNE, AIX MARSEILLE III - IMEP
UNIVERSITÉ PAUL CÉZANNE, AIX MARSEILLE III - IMEP
UNIVERSITÉ PAUL CÉZANNE, AIX MARSEILLE III - IMEP

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
<strong>UNIVERSITÉ</strong> <strong>PAUL</strong> <strong>CÉZANNE</strong>, <strong>AIX</strong> <strong>MARSEILLE</strong> <strong>III</strong><br />
École Doctorale des Sciences de l’Environnement<br />
Culture du Lentinula edodes (Berk.) Pegler sur résidus oléicoles en fermentation<br />
en milieu solide : Transformation des polyphénols des margines<br />
THÈSE<br />
présentée par<br />
Hicham LAKHTAR<br />
pour obtenir le grade de<br />
DOCTEUR DE L’<strong>UNIVERSITÉ</strong> <strong>PAUL</strong> <strong>CÉZANNE</strong><br />
Faculté des Sciences et Techniques - Saint Jérôme<br />
(Discipline: Biologie des Populations et Ecologie)<br />
Soutenue le 7 Décembre 2009<br />
JURY:<br />
N° 2009<strong>AIX</strong>30048<br />
Pr. Jean LE PETIT Université Paul Cézanne, Marseille Président<br />
Dr. Maurice BENSOUSSAN Université de Bourgogne, Dijon Rapporteur<br />
Dr. Jean-Michel SAVOIE INRA, Bordeaux Rapporteur<br />
Pr. Gerardo SAUCEDO-CASTANEDA UAM, México, Mexique Examinateur<br />
Dr. Sevastianos ROUSSOS IRD, Marseille Directeur de Thèse<br />
Pr. Mustapha ISMAILI-ALAOUI IAV-Hassan II, Rabat, Maroc Co-directeur de Thèse
Remerciements<br />
Cette thèse a débuté en Novembre 2006 grâce au financement du Département<br />
Soutien et Formation des communautés scientifiques du Sud (DSF SF SF) SF de l’Institut de<br />
Recherche pour le Développement (IRD IRD IRD). IRD<br />
Je tiens tout d’abord à adresser mes remerciements les plus sincères au Dr. Sevastianos<br />
ROUSSOS de m’avoir accordé sa confiance depuis 4 ans déjà et de m’avoir permis de réaliser<br />
ce travail de recherche dans son laboratoire. À travers ses qualités professionnelles, en tant<br />
que directeur de recherche, il m’a transmis de précieuses connaissances en Fermentation en<br />
Milieu Solide, et en Mycologie ainsi qu’une rigueur scientifique qui me seront très utiles<br />
dans mon avenir professionnel. Je tiens également à le remercier à titre personnel d’avoir<br />
toujours été présent pour me faciliter la vie en France, malgré ses préoccupations.<br />
Je souhaite remercier également le Pr. Mustapha ISMAILI-ALAOUI d’avoir accepté<br />
de participer à la co-direction de la thèse et de m’avoir accueilli dans son laboratoire à l’IAV IAV<br />
durant mon séjour au Maroc. Je remercie également le Pr. Thierry TATONI, le directeur de<br />
l’<strong>IMEP</strong>, de m’avoir accueilli au sein de l’Institut Méditerranéen d’Écologie et de<br />
Paléoécologie (<strong>IMEP</strong> <strong>IMEP</strong> <strong>IMEP</strong>) <strong>IMEP</strong> de l’Université Paul Cézanne d’Aix-Marseille <strong>III</strong>.<br />
Je remercie les membres de jury en particulier le Dr. Maurice BENSOUSSAN, maitre<br />
de conférences à l’Université de Bourgogne-Dijon, et le Dr. Jean-Michel SAVOIE, chargé<br />
de recherche à l’INRA, pour le temps qu’ils ont bien voulu consacrer à lire et à juger ce<br />
travail en tant que rapporteurs . Egalement je remercie le Pr. Jean LE PETIT, professeur<br />
Emérite de l’Université Paul Cézanne qui m’a fait l’honneur de présider ce jury et<br />
Pr. Gerardo SAUCEDO-CASTANEDA, professeur à l’Université Autonome<br />
Métropolitaine d’Iztapalapa au Mexique qui m’ont fait profité de leur large expérience en<br />
Microbiologie et en Biotechnologie en me donnant de précieuses remarques pour améliorer<br />
mon rapport écrit.
Je remercie le Dr. Antonis PHILIPPOUSSIS, chercheur à la fondation grecque<br />
NAGREF d’Athènes, d’avoir accepté à nous fournir les souches de Lentinula edodes<br />
utilisées pour la réalisation de ce travail.<br />
Je remercie vivement mes collègues de l’IRD, qui ont collaboré à ce travail : Dr.<br />
Isabelle GAIME-PERRAUD et Dr. Hervé MACARIE, ainsi que pour leur soutien et leurs<br />
conseils. By the same occasion, I would like to thank my friend Roopesh<br />
KRISHNANKUTTY for his help for improving the English of articles, and the moments<br />
we spent together, in Marseille, will remain as sweet memories for ever.<br />
J’exprime ma profonde reconnaissance à toutes les personnes du laboratoire IRD:<br />
Martine MARTINEZ, Yoan LABROUSSE, Fanny BAROT, Claude CHARPPY<br />
ROUBAUD pour leur aide et leur soutien.
Avant propos<br />
Le travail de thèse a été effectué en partie à l’<strong>IMEP</strong> Equipe d’Ecologie Microbienne et<br />
Biotechnologies à l’université Paul Cézanne, Marseille, France et en partie à l’Institut<br />
Agronomique et Vétérinaire Hassan II, de Rabat, Maroc. Durant mon stage de 6 ème année<br />
d’ingénieur en Industrie Agroalimentaire- IAV-Hassan II, j’ai réalisé un séjour en France de 3<br />
mois à l’Unité Biotrans d’Avril à Juin 2006 dans le cadre d’un projet PRAD (Programme de<br />
Recherche Agronomique pour le Développement) sur la valorisation des grignons d’olive par<br />
la production d’enzymes fongiques. Cette collaboration nous a permis de préparer ce projet de<br />
recherche afin d’obtenir une bourse de la part de l’IRD pour réaliser ma thèse de doctorat sur :<br />
Culture du Lentinula edodes (Berk.) Pegler sur résidus oléicoles en fermentation en<br />
milieu solide : Transformation des polyphénols des margines.<br />
Cette thèse a été financée par le Département Soutien et Formation des communautés<br />
scientifiques du sud (DSF), Institut de Recherche pour le Développement (IRD), France, pour<br />
une durée de trois ans. Le Pr. Mustapha Ismaili-Alaoui, Professeur à l’IAV-Hassan II de<br />
Rabat et le Dr. Sevastianos Roussos, Directeur de Recherche à l’IRD, ont assuré la direction<br />
scientifique de ce travail de recherche scientifique. Dr. Isabelle Perraud-Gaime CR IRD et Dr.<br />
Anne Marie Farnet (MC Université de Provence), ainsi que Dr. Hervé Macarie CR IRD, Dr.<br />
Jean Michel Savoie, CR INRA-Bordeaux et Dr. Elisée Ferré MC UPCAM, ont participé aux<br />
réunions du Comité de pilotage.
Résumé<br />
L’objectif du travail a été d’étudier la valorisation de sous-produits oléicoles au<br />
Maroc par la culture en fermentation en milieu solide de champignons comestibles avec une<br />
double finalité: (i) la production de la biomasse fongique pour l’alimentation humaine et (ii)<br />
la détoxification des margines par la dégradation des composés phénoliques. Une enquête<br />
menée sur le terrain au Maroc a permis de faire le point sur la situation du secteur oléicole au<br />
Maroc. A partir de seize souches de L. edodes, une souche performante à la détoxification des<br />
margines a été sélectionnée pour mener la culture de L. edodes. La production de l’inoculum<br />
sur les grains de blé (spawn) a été optimisée. Un mélange des substrats oléicoles solides<br />
(SOS), composé de bois de taille d’olivier, de grignons d’olive et de margines, a été mis au<br />
point et les conditions de croissance de L. edodes sur ce mélange ont été optimisées. Le<br />
changement de la matière organique durant le développement de L. edodes sur le mélange<br />
dans des conditions optimales a été analysé par les techniques spectroscopiques de la RMN<br />
solide du 13 C couplées aux analyses biochimiques. Une technique de séchage solaire sous serre a<br />
été mise au point permettant la réduction en volume et le conditionnement des grignons d’olives<br />
mélangés aux margines. De ce travail, nous concluons la possibilité de valoriser l’ensemble des<br />
co-produits de l’industrie oléicole par la culture de L. edodes permettant, d’une part, la<br />
production d’une biomasse mycélienne importante et, d’autre part, la dégradation des<br />
composés phénoliques présents dans les margines. Ces résultats novateurs ouvrent des<br />
perspectives sérieuses pour la production de L. edodes sur le SOS.
Table de Matières<br />
1. INTRODUCTION ............................................................................................................. 1<br />
2. ANALYSE BIBLIOGRAPHIQUE.................................................................................. 5<br />
2.1. Le Secteur oléicole au Maroc .................................................................................... 5<br />
2.1.1. L’olivier ............................................................................................................... 5<br />
2.1.2. L’huile d’olive ..................................................................................................... 5<br />
2.1.3. Les unités de trituration d’olives au Maroc ..................................................... 6<br />
2.1.4. Les bilans quantitatifs des sous produits de l’industrie oléicole .................... 8<br />
2.1.5. Le bois de taille de l’olivier ................................................................................ 9<br />
2.1.6. Les grignons d’olive ........................................................................................... 9<br />
2.1.7. Les caractéristiques des margines d’olive ...................................................... 10<br />
2.1.7.1. pH, conductivité et matière sèche ................................................................ 11<br />
2.1.7.2. Charge minérale et métaux lourds .............................................................. 11<br />
2.1.7.3. Fraction organique ....................................................................................... 12<br />
2.1.7.4. Fraction phénolique ...................................................................................... 12<br />
2.1.7.5. Caractéristiques microbiologiques .............................................................. 13<br />
2.2. L’impact des margines d’olive sur l’environnement, traitements et valorisation .<br />
................................................................................................................................... 14<br />
2.2.1. L’impact sur les eaux : cas de l’Oued Sebou (Fès) ........................................ 14<br />
2.2.2. L’impact sur le sol ............................................................................................ 15<br />
2.2.3. Les traitements des margines .......................................................................... 16<br />
2.2.4. La valorisation des margines ........................................................................... 17<br />
2.2.4.1. Extraction des antioxydants (hydroxytyrosol) ........................................... 18
2.2.4.2. Culture de champignons supérieurs ........................................................... 18<br />
2.3. Le champignon comestible, Lentinula edodes (Berk.) Pegler .............................. 19<br />
2.3.1. Définition des champignons ............................................................................. 19<br />
2.3.2. La physiologie de croissance des champignons ............................................. 19<br />
2.3.3. Les techniques générales de culture de L. edodes .......................................... 20<br />
2.3.3.1. Culture sur bûches........................................................................................ 20<br />
2.3.3.2. Culture sur substrats synthétiques ............................................................. 21<br />
2.3.3.3. Culture sur substrats alternatifs ................................................................. 22<br />
2.3.4. Les phénoloxydases, les laccases ..................................................................... 23<br />
2.3.4.1. Structure des laccases ................................................................................... 23<br />
2.3.4.2. Mécanismes d’action des laccases ............................................................... 23<br />
2.3.4.3. Applications des laccases .............................................................................. 24<br />
2.4. Les fermentations en milieux solides ..................................................................... 24<br />
2.4.1. Définition des FMS ........................................................................................... 25<br />
2.4.2. Les principaux paramètres des FMS .............................................................. 25<br />
2.4.2.1. Microorganismes et l’inoculation du milieu solide .................................... 26<br />
2.4.2.2. Choix du support/substrat ........................................................................... 26<br />
2.4.2.3. Activité de l’eau et l’humidité ...................................................................... 26<br />
2.4.2.4. Aération des cultures en FMS ..................................................................... 27<br />
2.4.2.5. Contrôle de la température pour les FMS .................................................. 28<br />
2.4.2.6. Evolution du pH ............................................................................................ 28<br />
2.4.3. Les applications des FMS ................................................................................ 29<br />
2.4.4. Les avantages et les inconvénients de la FMS................................................ 30
2.5. Conclusion de l’analyse bibliographique ............................................................... 30<br />
3. MATERIEL ET METHODES ...................................................................................... 33<br />
3.1. Les substrats oléicoles de culture ........................................................................... 33<br />
3.1.1. La description de l'unité de trituration semi industrielle ............................. 33<br />
3.1.2. L’origine du substrat ....................................................................................... 34<br />
3.1.3. La préparation et le conditionnement du substrat ........................................ 35<br />
3.2. Les souches utilisées................................................................................................. 35<br />
3.3. Les milieux de culture ............................................................................................. 36<br />
3.3.1. Les milieux d'entretien ..................................................................................... 36<br />
3.3.2. Les milieux gélosés de criblage des souches ................................................... 36<br />
3.3.3. Les milieux à base de céréale pour la culture Spawn .................................... 36<br />
3.4. Les stratégies d'inoculation..................................................................................... 37<br />
3.4.1. L’inoculation par carrés de milieux gélosés ................................................... 37<br />
3.4.2. L’inoculation sur feuille de cellophane........................................................... 37<br />
3.4.3. L’inoculation par grain de céréales ................................................................ 38<br />
3.5. La mise en œuvre des FMS ..................................................................................... 39<br />
3.5.1. La préparation du substrat oléicole ................................................................ 39<br />
3.5.2. Les cultures en Erlenmeyer ............................................................................. 39<br />
3.5.3. Les cultures en colonnes de Raimbault .......................................................... 39<br />
3.5.4. La régulation des paramètres de FMS ........................................................... 40<br />
3.5.5. La description et le fonctionnement du PNEO .............................................. 40<br />
3.6. L’échantillonnage et le traitement des échantillons en FMS ............................... 42<br />
3.7. Les techniques d’analyse ......................................................................................... 42
3.7.1. La mesure de l’humidité .................................................................................. 42<br />
3.7.2. La mesure du pH .............................................................................................. 43<br />
3.7.3. La mesure de la croissance apicale ................................................................. 43<br />
3.7.4. L’estimation indirecte de la biomasse par analyse de l’ergostérol .............. 43<br />
3.7.4.1. Extraction de l’ergostérol ............................................................................ 43<br />
3.7.4.2. Dosage de l’ergostérol en HPLC ................................................................. 43<br />
3.7.5. Le dosage des sucres réducteurs ..................................................................... 44<br />
3.7.6. Le dosage des sucres totaux ............................................................................. 44<br />
3.7.7. Le dosage des phénols totaux .......................................................................... 44<br />
3.7.8. Le dosage des activités enzymatiques ............................................................. 45<br />
3.7.8.1. Activité Laccase ............................................................................................ 45<br />
3.7.8.2. Activité Mn peroxydase................................................................................ 45<br />
3.7.8.3. Activité Lignine peroxydase ........................................................................ 45<br />
3.7.8.4. Activité tannase ............................................................................................. 45<br />
3.7.8.5. Activité lipase ................................................................................................ 46<br />
3.7.9. L’analyse des monomères phénoliques par HPLC ....................................... 46<br />
3.7.10. Les analyses élémentaires : C et N .................................................................. 47<br />
3.7.11. La résonance magnétique nucléaire 13 C du solide ......................................... 47<br />
Chapitre I .................................................................................................................................. 51<br />
Criblage des souches de Lentinula edodes cultivées sur des margines d’olive en culture<br />
solide et liquide pour la biodégradation des polyphénols ................................................... 51<br />
Chapitre II ................................................................................................................................ 70<br />
Optimisation de la production de l’inoculum (spawn) de Lentinula edodes sur les grains<br />
de blé (Ebly®) ......................................................................................................................... 70
Chapitre <strong>III</strong> ............................................................................................................................... 87<br />
Culture de Lentinula edodes en fermentation en milieu solide sur substrat oléicole solide:<br />
Evaluation des facteurs de croissance .................................................................................. 87<br />
Chapitre IV ............................................................................................................................. 106<br />
Transformation de la matière organique et détoxification des substrats oléicoles solides<br />
par la culture de Lentinula edodes par fermentation en milieu solide............................. 106<br />
Chapitre V .............................................................................................................................. 125<br />
Disponibilité des substrats oléicoles : séchage solaire sous serre du mélange de grignons<br />
d’olive et de margines .......................................................................................................... 125<br />
Synthèse générale ................................................................................................................. 143<br />
Conclusions et perspectives ................................................................................................. 151<br />
Références bibliographiques ............................................................................................... 154
Liste des Tables<br />
Tableau 1. Bilan matière en grignons et margines obtenus par différents procédés de<br />
trituration des olives, 100 kg d’olives donnent en moyenne 20 kg d’huile d’olive 9<br />
Tableau 2. Composition chimique indicative des différents types de grignons 10<br />
Tableau 3: Valeurs minimales et maximales des minéraux et métaux lourds déterminées sur<br />
les margines issues des deux systèmes d'extraction de l’huile 11<br />
Tableau 4. Structure te teneur des monomères phénoliques rencontrés dans les margines 13<br />
Tableau 5. Procédés de traitement des margines 17<br />
Tableau 6 : Exemples de substrats utilisés et les applications en FMS 29<br />
Tableau 7. Comparaison de la fermentation en milieu liquide (FML) avec la fermentation en<br />
milieu solide (FMS) 30<br />
Tableau 8. Liste des espèces et origine des souches de champignons comestibles étudiées 35<br />
Tableau 9. Conditions d’analyses en HPLC pour la détection des monomères phénoliques 46<br />
Tableau 10. Conditions d’élution pour la détection des monomères phénoliques 47<br />
Tableau 11 . Attribution des régions RMN 13 C aux différents groupes fonctionnels trouvés<br />
dans la matière organique 48
Liste des Figures<br />
Figure 1. Evolution de la superficie de culture de l'olivier au Maroc 5<br />
Figure 2. Evolution de la production de l'huile d'olive au Maroc 6<br />
Figure 3. Répartition de la production mondiale de l’huile d’olive (%) pendant les 5 dernières<br />
années (2003 – 2008) 6<br />
Figure 4. Répartition et capacités de trituration des unités artisanales et industrielles au Maroc<br />
7<br />
Figure 5. Schéma de l’industrie oléicole équipée d’un procédé triphasique avec le système de<br />
presse 8<br />
Figure 6. Evolution de la DCO dans les eaux l’Oued Sebou avant, pendant et après la période<br />
d’activité des huileries 15<br />
Figure 7. Processus général de culture de L. edodes sur bûches 21<br />
Figure 8. Localisation de la région Tadla Azilal au Maroc 33<br />
Figure 9. Unité de trituration semi industrielle au Maroc 34<br />
Figure 10 . Olives de la variété picholine de maturité avancée 34<br />
Figure 11. Technique pour le suivi de la croissance apicale du mycélium et la mesure de<br />
biomasse des champignons en milieu gélosé 38<br />
Figure 12. Dispositif de FMS utilisé: 1, Entrée d’air ; 2, Bain marie thermostaté ; 3, Colonnes<br />
de Raimbault 40<br />
Figure 13 . Photographie du respiromètre PNEO et description de ses différents composants<br />
41<br />
Figure 14. Exemple de spectre CPMAS RMN 13 C d’une litière de Quercus ilex L. 48<br />
Figure 15 : Activité laccase produite par L. edodes Le119 sur des grains d’Ebly sans margines<br />
et avec des margines à 10% (v/v), incubés à 25°C avec aération forcée de 25 mL.min -1<br />
pendant 20 jours 71
1. INTRODUCTION<br />
Introduction<br />
Au Maroc, le secteur oléicole joue un rôle très important sur le plan socio-économique.<br />
Il contribue activement à la fixation des populations en milieu rural en offrant plus de<br />
15 millions de journées de travail (DPV, 2009). Avec une production nationale de<br />
75 000 tonnes d’huile d’olive, le Maroc occupe le 6 ème rang après l’Espagne, l’Italie, la<br />
Tunisie, la Turquie, et la Grèce. L’industrie oléicole engendre, en plus de l’huile comme<br />
produit principal, de grandes quantités de sous produits solides (grignons d’olive, feuilles et<br />
bois de taille), et liquides (margines). Les quantités annuelles des grignons ont été estimées à<br />
180 000 tonnes, tandis que les margines représentent un volume de 250 000 m 3 (Ben Sassi et<br />
al., 2006). Selon les estimations, 25 kg de feuilles et brindilles (diamètre inférieur à 3 cm)<br />
sont produites par an et par arbre (Nefzaoui, 1995).<br />
Avec la promotion des vertus bénéfiques de l’huile d’olive pour la santé, la demande<br />
mondiale ne cesse d’augmenter et par conséquent la production croît constamment pour<br />
répondre aux besoins des consommateurs (COI, 2005). Ainsi, la création des moulins pour la<br />
trituration des olives de plusieurs producteurs, forme des points de concentration des<br />
polluants non résorbables par le pouvoir épurateur des milieux. Or, pour les huileries, la<br />
nécessité de produire une huile de qualité, avec le minimum d’impacts négatifs sur<br />
l’environnement, est une condition essentielle pour qu’elles puissent rester concurrentielles<br />
sur le marché. De plus, l’image de pollueur peut nuire à l’image de marque de l’entreprise et<br />
peut l’exposer à des sanctions Pollueur- Payeur en fonction de la législation qui devient de<br />
plus en plus contraignante (Benyahia et Zein, 2003).<br />
Au Maroc, le bois de taille de l’olivier (rameaux, brindilles et feuilles) ne connaît pas<br />
d’utilité à part quelques utilisations comme combustible et aliment pour bétail (Nefzaoui,<br />
1995). Par ailleurs dans d’autres pays, leur composition chimique, en particulier des feuilles, a<br />
orienté de nombreuses études vers l’extraction et la bioconversion des polyphénols en<br />
antioxydants d’intérêt industriel (Bouaziz et al., 2008 ; Lee et al., 2009).<br />
Les moulins industriels de grande capacité posent de sérieux problèmes<br />
environnementaux. Les margines, provenant de ces moulins, ne subissent aucun traitement et<br />
sont souvent déversées dans les égouts d’assainissement, stockées dans des bassins<br />
d’évaporation et/ou épandues directement sur le sol sans contrôle. Il en résulte un impact<br />
négatif sur l’environnement qui se traduit par le colmatage des sols, la pollution des eaux<br />
superficielles et souterraines et le dégagement d’odeurs nauséabondes. L’exemple de la<br />
situation de l’Oued Sebou (Fès) est une illustration de ce fléau (Foutlane et al., 2002).<br />
1
Introduction<br />
La toxicité des margines est attribuée à la présence des acides gras libres à longue<br />
chaîne et de composés récalcitrants, difficiles à dégrader comme les composés phénoliques,<br />
en grande concentration (4-15 g.L -1 ), qui sont responsables d’effets phytotoxiques et<br />
antimicrobiens (Sayadi et al., 2000; El Hajjouji et al., 2007). Les grignons d’olive possèdent<br />
une filière de valorisation bien développée qui consiste à leur utilisation dans plusieurs<br />
domaines comme l’extraction d’huile de grignons destinée à la fabrication du savon (Serghini<br />
et al., 2007), l’alimentation de bétail (Haddadin et al., 2002), comme substrat pour la<br />
production d’enzymes (Cordova et al., 1998) et pour la culture de champignons comestibles<br />
Pleurotus sp. (Zervakis et al., 1996) et Agaricus bisporus (Altieri et al., 2009).<br />
De nombreuses études ont été menées avec le souci de trouver des solutions permettant<br />
le traitement des margines par des voies physiques, chimiques et/ou biologiques (Ranalli,<br />
1991 ; Achak et al., 2008 ; Hajjouji et al., 2008). Certes, l’épandage des margines dans les<br />
champs d’oliviers est une technique permettant un apport de minéraux (N, P) au sol et une<br />
élimination radicale des margines. Cependant, cette technique est une pratique controversée<br />
par la communauté scientifique (Mekki et al., 2007). Actuellement, avec les contraintes<br />
économiques, il n’existe pas encore une solution universelle pour le traitement des margines.<br />
Bien que la plupart des micro-organismes ne puisse se développer sur des substrats<br />
contenant des polyphénols, certains champignons filamenteux comme les Aspergillus et des<br />
champignons comestibles, comme Pleurotus sp. (Pleurotes) et Lentinula edodes (shiitake),<br />
sont capables de se développer et de produire des enzymes impliquées dans la dégradation ou<br />
la biotransformation de ces molécules en les utilisant comme sources de carbone (Tomati et<br />
al., 2004).<br />
Au niveau mondial, après l’A. bisporus (champignon de Paris), le deuxième<br />
champignon commercialisé est L. edodes avec une production de plus de deux millions de<br />
tonnes en 2002 (Royse et Sanchez-Vazquez, 2003). Depuis 25 ans (1985) le taux<br />
d’augmentation de la production mondiale de L. edodes est d’environ 25% (Chang, 1999). La<br />
plus grande partie de cette production commerciale (90% de la production mondiale) est<br />
réalisée dans le Sud Est Asiatique principalement en Chine (Royse, 2001). Le champignon a<br />
attiré l’attention de plusieurs chercheurs par ses aptitudes thérapeutiques qui sont attribuées<br />
principalement au lentinan (Tomati et al., 2004). Le lentinan est utilisé pour ses effets anti-<br />
tumoral, anti-inflammatoire, antiviral, antibactérien, antiparasite. Il permet la régulation de la<br />
pression artérielle, et il est utilisé également comme agent antidiabétique et hypoglycémiant<br />
(Tomati et al., 2004).<br />
2
Introduction<br />
L'utilisation de la fermentation en milieu solide (FMS) constitue une approche originale<br />
pour connaître la physiologie de croissance du mycélium des champignons comestibles<br />
(Martínez-Carrera et al., 1985; Roussos et al., 1997), étant donné que la plupart des études<br />
réalisées sur la culture de champignons comestibles concernent essentiellement la production<br />
de carpophores pour l'alimentation humaine (Martínez-Carrera et al., 1985; Pandey et al.,<br />
2000) sans tenir compte des potentialités offertes par la biomasse mycélienne pour<br />
l’enrichissement en protéine du résidu solide (meule) destiné à l’alimentation animale (sobal,<br />
2002).<br />
Le but de ce travail est de maîtriser la culture du mycélium de L. edodes sur un mélange<br />
des substrats oléicoles (grignons d’olive, bois de la taille et les margines d’olive) pour (i) la<br />
production de la biomasse mycélienne de L. edodes qui servira à la production du carpophore<br />
destiné à l’alimentation humaine, et (ii) la détoxification des margines. Le substrat épuisé<br />
pourrait être destiné à l’alimentation animale après sa détoxification.<br />
Tout au long de ce travail, nous suivrons l’évolution de la production de la biomasse<br />
mycélienne de L. edodes ainsi que la dégradation des phénols totaux des margines. Nous<br />
étudierons, dans un premier temps, la tolérance des souches de L.edodes aux margines afin de<br />
sélectionner une souche avec double compétence ;(i) production d’une grande biomasse<br />
mycélienne en milieu solide et (ii) réduction importante de la charge phénolique des<br />
margines. Puis nous procéderons à l’optimisation de la production de l’inoculum de<br />
L. edodes sur les grains de blé « spawn », ainsi que les conditions de culture de L. edodes sur<br />
les substrats oléicoles. Par la suite, une fermentation en milieu solide de L. edodes sur les<br />
substrats oléicoles dans des conditions optimales sera suivie par respirométrie. Cette<br />
expérience nous permettra de vérifier l’hypothèse initiale de ce travail:<br />
« Est il possible de valoriser les trois co-produits de l’industrie oléicole (bois de taille,<br />
grignons d’olives et margines) pour en faire un seul substrat oléicole solide, utilisé pour la<br />
culture de Lentinula edodes en FMS».<br />
L’étude bibliographique que nous présentons au début de ce travail, permettra de définir<br />
la problématique des substrats de l’industrie oléicole, les techniques de culture de L.edodes,<br />
ainsi qu’un état des connaissances des techniques de FMS. Au cours de cette étude, nous<br />
ferons également le point sur l’état actuel des recherches sur le traitement et valorisation des<br />
substrats de l’industrie oléicole.<br />
3
2. ANALYSE BIBLIOGRAPHIQUE<br />
2.1. Le Secteur oléicole au Maroc<br />
2.1.1. L’olivier<br />
Matériel et Méthodes<br />
L'olivier constitue la principale espèce fruitière plantée au Maroc; il occupe une surface<br />
de 650 000 ha (DPV, 2009) soit plus de 58% de la superficie arboricole totale (Fig. 1). Bien<br />
que la superficie des oliveraies connaisse une évolution progressive ces dernières années,<br />
l’évolution ne répond pas aux objectifs du plan national oléicole (PNO) visant de porter la<br />
superficie oléicole à un million d'hectares à l'horizon de 2010 (DPV, 2009).<br />
Figure 1. Evolution de la superficie de culture de l'olivier au Maroc (DPV, 2009).<br />
2.1.2. L’huile d’olive<br />
La production d'olives enregistrée en 2007/2008 est de l’ordre de 750.000 tonnes (DPV,<br />
2009). La variété Picholine marocaine représente 95% des variétés cultivées au Maroc, pour<br />
produire à la fois, les olives de table (4 ème place mondiale) et l’huile d’olive (HO). Malgré le<br />
caractère fluctuant de la production d’HO, son évolution depuis 2002 montre une tendance à<br />
la hausse qui est due essentiellement à une extension des superficies plutôt qu'à une<br />
amélioration des rendements (oscillant entre 0,7 et 1,7 T.Ha -1 d’après DPV, 2008), lesquels<br />
rendements restent faibles comparativement aux autres pays oléicoles (Grèce : 1,7 T.Ha -1 ,<br />
Espagne : 1,6 T.Ha -1 ) (COI, 2008). La production en 2003-2004 a connu une exception, avec<br />
une production d'huile d'olive de 112 000 tonnes (Fig. 2). Le Maroc occupe ainsi le sixième<br />
rang mondial parmi les producteurs du pourtour méditerranéen, qui génère près de 95% de la<br />
production mondiale en huile d’olive (Fig. 3).<br />
5
Matériel et Méthodes<br />
Figure 2. Evolution de la production de l'huile d'olive au Maroc (DPV, 2009).<br />
Huile d’olive (%)<br />
Figure 3. Répartition de la production mondiale de l’huile d’olive (%) pendant les 5<br />
dernières années (2003 – 2008) (COI, 2008).<br />
2.1.3. Les unités de trituration d’olives au Maroc<br />
Au Maroc, le secteur de trituration d’olive est caractérisé par la coexistence de moulins<br />
traditionnels (maâsras) et d’unités semi-industrielles et industrielles. Concernant le secteur<br />
traditionnel, on note l’existence de plus de 15 600 maâsras avec une capacité annuelle de<br />
trituration qui couvre près de 50% de la production nationale, alors que le secteur moderne et<br />
semi-moderne est représenté par 334 unités ayant une capacité minimale de trituration<br />
d’environ 10 tonnes/jour et susceptible d’être augmentée de façon substantielle (Fig. 4).<br />
6
Matériel et Méthodes<br />
Les maâsras fonctionnent majoritairement avec le procédé triphasique par le système de<br />
presse ou de centrifugation; et ce procédé génère deux types d’effluents solides (grignons<br />
d’olive) et liquide (margines d’olive). En plus de cette technologie, le Maroc a introduit le<br />
procédé à deux phases qui génère des grignons humides (50 et 60 %), et une faible production<br />
de margines.<br />
Figure 4. Répartition et capacités de trituration des unités artisanales et industrielles au<br />
Maroc (DPV, 2009).<br />
Nombre de maâsras<br />
Dans les maâsras fonctionnant avec le système de presse, les olives sont broyées et<br />
malaxées à l’aide de broyeurs comportant une ou deux meules qui tournent avec une énergie<br />
électrique. La pâte d’olive obtenue est répartie manuellement dans des sacs ou scourtins qui<br />
sont ensuite empilés sous des presses hydrauliques. Il en résulte, après pression, un résidu<br />
solide appelé grignon brut et un moût huileux constitué de l’eau de végétation (les margines)<br />
7
Matériel et Méthodes<br />
et de l’huile d’olive. La récupération de l’huile est obtenue après décantation accompagnée<br />
d’une séparation par une centrifugation (Fig. 5).<br />
Figure 5. Schéma de l’industrie oléicole équipée d’un procédé triphasique avec le<br />
système de presse (Ismaili-Alaoui et Heddoun, 2006).<br />
2.1.4. Les bilans quantitatifs des sous produits de l’industrie oléicole<br />
L’industrie oléicole engendre, en plus de l’huile d’olive comme produit principal, trois<br />
sous produits de type solide (produits de la taille des oliviers, les grignons d’olive) et de type<br />
liquide (margines d’olive) (Fig. 6).<br />
Après la récolte des olives, les oliviers subissent en général une taille sévère un an sur<br />
deux et une taille légère l’année suivante. Les quantités des produits de la taille ont été<br />
estimées à 25 kg de feuille et brindilles (diamètres inferieurs à 3 cm) produites par an et par<br />
arbre, selon Nefzaoui (1995).<br />
8
Matériel et Méthodes<br />
Par ailleurs, les quantités des grignons d’olive et des margines dépendent étroitement du<br />
procédé de trituration appliqué. Les bilans massiques en grignons et en margines pour chaque<br />
procédé de trituration appliqué au Maroc sont reportés dans le Tableau 1.<br />
Tableau 1. Bilan matière en grignons et margines obtenus par différents procédés de<br />
trituration des olives, 100 kg d’olives donnent en moyenne 20 kg d’huile d’olive.<br />
Procédé de Grignons d’olive Margines Références<br />
trituration<br />
d’olive<br />
Triphasique par 40 kg avec 30-40% 40 kg Amirante et al., 1993<br />
presse<br />
d’humidité<br />
Triphasique avec<br />
centrifugation<br />
55 kg avec 40-50%<br />
d’humidité<br />
biphasique 70 kg avec 60-70%<br />
d’humidité<br />
2.1.5. Le bois de taille de l’olivier<br />
>100 kg Amirante et al., 1993<br />
10 kg Di Giovacchino, 1988 ;<br />
Bing et al., 1994<br />
La composition chimique des feuilles et brindilles varie en fonction de nombreux<br />
facteurs; variété, conditions climatiques, époque de prélèvement, âge des plantations<br />
(Nefzaoui, 1995). La teneur en matières azotées totales des feuilles varie de 9 à 13%, alors<br />
que celle des rameaux ne dépasse guère 3 à 6 %. La teneur en lignine est de 18 à 20 %. Les<br />
résidus de la taille ont des applications nombreuses, entre autres l’utilisation dans<br />
l’alimentation animale. Ils peuvent être utilisés comme combustibles, servir à la fabrication de<br />
compost, ou constituer la matière première dans l’industrie du papier ou la fabrication des<br />
meubles (Nefzaoui, 1995). Récemment, une étude a été menée dans le but d’améliorer la<br />
digestibilité et la valeur nutritive de cette biomasse (feuilles d’olive et brindilles) par un<br />
traitement biologique (Fayed et al., 2009) consistant en une culture de moisissures. D’autres<br />
études ont porté sur la bioconversion des polyphénols des feuilles en antioxydants destinés<br />
aux industries agroalimentaire et pharmaceutique (Bouaziz et al., 2008 ; Lee et al., 2009). Par<br />
ailleurs, à cause de la disponibilité saisonnière des feuilles d’oliviers, d’autres auteurs ont mis<br />
l’accent sur le séchage des feuilles afin d’assurer une conservation et une disponibilité de<br />
cette biomasse toute l’année pour des éventuelles applications (Molina-Alcaide, 2008; Erbay<br />
et Icier 2009).<br />
2.1.6. Les grignons d’olive<br />
Les principaux déchets solides générés lors de l’élaboration de l’huile d’olive sont les<br />
grignons. Les grignons d’olives contiennent une quantité d’huile résiduelle qu’il n’est pas<br />
possible d’extraire par des moyens physiques et qui est obtenue dans les installations<br />
d’extraction d’huile de grignons par l’utilisation des solvants comme l’hexane. La<br />
9
Matériel et Méthodes<br />
composition des grignons dépend étroitement de la variété d’olives, le degré de maturation<br />
des olives et le système employé lors de l’extraction de l’huile d’olive. Le tableau 2 montre la<br />
composition chimique des grignons obtenus lors de l’élaboration de l’huile d’olive avec<br />
différents procédés et provenant d’origines variées.<br />
Contrairement aux autres tourteaux oléagineux, les grignons d’olive bruts sont pauvres<br />
en matières azotées et riches en cellulose brute. Ils restent relativement riches en matières<br />
grasses (huile d’olive). Le dénoyautage partiel réduit les teneurs en cellulose brute par<br />
l’élimination partielle des noyaux d’olives. Dans une autre étude, Chiofalo et al. (2004) ont<br />
constaté une nette différence entre le rendement gras des grignons de pressoir et les grignons<br />
des systèmes en continu. La différence est due fondamentalement à l’efficacité d’extraction<br />
des systèmes en continu par rapport aux systèmes traditionnels.<br />
Tableau 2. Composition chimique indicative des différents types de grignons (DPV, 2009).<br />
Type Matière<br />
* p.d. = partiellement dénoyauté<br />
2.1.7. Les caractéristiques des margines d’olive<br />
Les margines ont un aspect trouble, une coloration brun-rougeâtre à noire. Cette couleur<br />
est fonction de l’état de dégradation des composés phénoliques et des olives dont ils dérivent<br />
(Hamdi et Ellouz, 1993). Leur odeur rappelle celle de l’huile d’olive, mais elle peut devenir<br />
gênante lors des phénomènes de rancissement ou de fermentation anaérobie (Ranalli, 1991).<br />
Les composés fondamentaux des margines sont l’eau (83.2 %), les substances organiques<br />
(15 %) et les substances minérales (1.8 %) (Fiestas et Borja 1992). Les variations des<br />
caractéristiques des margines résultent du type du procédé d’extraction de l’huile, de la<br />
qualité et de la variété des olives et de la conduite des opérations d’extraction (Mouncif et al.,<br />
1993).<br />
sèche (%)<br />
% de la matière sèche<br />
Matière<br />
minérale<br />
Matière<br />
azotée totale<br />
Cellulose<br />
brute<br />
Matière<br />
grasse<br />
Grignon brut 75 - 80 3 - 5 5 - 10 35 - 50 8 - 15<br />
Grignon gras p. d.* 80 - 95 6 - 7 9 - 12 20 - 30 15 - 30<br />
Grignon épuisé 85 - 90 7 - 10 8 - 10 35 - 40 4 - 6<br />
Grignon épuisé p.d. 85 - 90 6 - 8 9 - 14 15 - 35 4 - 6<br />
Pulpe grasse 35 - 40 5 - 8 9 - 13 16 - 26 26 - 33<br />
10
2.1.7.1. pH, conductivité et matière sèche<br />
Matériel et Méthodes<br />
Les margines ont un pH acide avec des valeurs comprises entre 4.2 et 5.9 (Eroglu et al.,<br />
2008). Lors des traitements biologiques des margines, une correction du pH peut s'avérer<br />
nécessaire (El Hajjouji et al., 2008). La chaux vive a été souvent utilisée pour ajuster le pH<br />
des margines. En effet, Ranalli (1991) rapporte qu'il faut 1 kg de chaux par 1 m 3 d'effluent<br />
afin de faire augmenter le pH d'un point.<br />
La conductivité électrique est étroitement liée à la concentration des substances<br />
dissoutes et à leur nature. Dans le cas des margines, les valeurs de cette conductivité varient<br />
entre 18 et 50 ms.cm -1 (Di Serio et al., 2008). Cette mesure ne donne pas forcément une idée<br />
immédiate sur la charge minérale du milieu (Paredes et al., 2005). L'extrait sec des margines<br />
est extrêmement élevé et variable. Il est situé entre 15.5 et 266.5 kg.m -3 pour les margines de<br />
pression et entre 9.5 et 161.2 kg.m -3 pour les margines de centrifugation (Di Giovacchino et<br />
al, 1988).<br />
2.1.7.2. Charge minérale et métaux lourds<br />
Les margines sont très riches en cendres, leur teneur varie de 4 à 42 g.L -1 pour celles<br />
issues de l'extraction par pression et 0.4 à 12.5 g.L -1 pour celles de centrifugation (COI, 2008).<br />
Tableau 3: Valeurs minimales et maximales des minéraux et métaux lourds déterminées sur les<br />
margines issues des deux systèmes d'extraction de l’huile (COI, 2008).<br />
Eléments<br />
(mg.L -1 )<br />
Procédé d’extraction de l’huile<br />
Pression Centrifugation<br />
Sodium 38 - 285 18 - 124<br />
Potassium 1500 - 5000 630 - 2500<br />
Calcium 58 - 408 47 - 200<br />
Magnésium 90 - 336 60 - 180<br />
Fer 16.40 - 86.40 8.80 - 31.50<br />
Cuivre 1.10 - 4.75 1.16-3.42<br />
Zinc 1.60 - 6.50 1.42-4.48<br />
Manganèse 2.16 - 8.90 0.87 - 5.20<br />
Nickel 0.44 - 1.58 0.29 - 1.44<br />
Cobalt 0.18 - 0.96 0.12-0.48<br />
Plomb 0.40 - 1.85 0.35 - 0.72<br />
11
Matériel et Méthodes<br />
Il ressort du Tableau 3 que les margines sont très riches en potassium, ce qui a conduit<br />
plusieurs chercheurs à tester leur pouvoir fertilisant. La différence de concentration, en<br />
minéraux et métaux lourds des margines entre les deux procédés, peut être expliquée par le<br />
fait que, durant la trituration des olives par le procédé de centrifugation, on ajoute de l’eau<br />
chaude, ce qui entraîne une diminution de la concentration des éléments minéraux.<br />
2.1.7.3. Fraction organique<br />
Les margines d’olive ont un pouvoir polluant très important avec une demande<br />
biologique en oxygène (DBO) de 100 g.L -1 (Rosario et al., 1999) et une demande chimique en<br />
oxygène (DCO) de 200 g.L -1 (Rosario et al., 1999). Ces valeurs sont 200 à 400 fois<br />
supérieures à celles des eaux municipales (Cossu et al., 2001). La matière organique des<br />
effluents d’huileries d’olive est constituée par des polysaccharides (13-53%), des protéines (8-<br />
16%), des composés phénoliques (2-15%), des lipides (1-14%), des polyalcools (3-10%) et<br />
des acides organiques (3-10%) (Fiestas et Borja, 1992). Cette composition résulte de la<br />
destruction des tissus de l’olive au cours de la trituration et de l’extraction de l’huile (Cossu et<br />
al., 1993 ; Al Mallah et al., 2000; Centi et al., 2000; Chamkha et al., 2001).<br />
2.1.7.4. Fraction phénolique<br />
Les phénols totaux sont en quantité appréciable (5,5 à 12 g.L -1 de margines) dans les<br />
effluents des huileries d'olive (Fenice et al., 2003 ; Ben Sassi et al., 2006). Les monomères les<br />
plus rencontrés dans les margines sont présentés dans le Tableau 4. La formation des phénols<br />
libres dans les margines est due à l'hydrolyse enzymatique des glucosides et des esters de la<br />
pulpe des olives, au cours du processus d'élaboration de l'huile (Ranalli, 1991). Le pigment<br />
catéchol-mélaninique (flavonoïde) ne se retrouve pas dans les olives mais se forme pendant le<br />
broyage des olives à partir des ortho-diphénols dont la pulpe est riche, sous l'action des<br />
polyphénol-oxydases (enzymes inactives dans les drupes entières) qui en déterminent la<br />
quinonisation, puis la polymérisation. Les margines sont riches en hydroxytyrosol qui possède<br />
une activité antioxydante très importante recherchée dans le domaine agroalimentaire,<br />
cosmétique et pharmaceutique (Manna et al., 1999 ; D'Angelo et al., 2005 ; Hamden et al.,<br />
2009).<br />
12
Matériel et Méthodes<br />
Tableau 4. Structure te teneur des monomères phénoliques rencontrés dans les margines.<br />
Composés phénoliques Structure Teneurs Références<br />
Acide cafféique (mM)<br />
Acide p-coumarique<br />
(mM)<br />
Oleuropéine (mM)<br />
Hydroxytyrosol (mg.L -1 )<br />
Tyrosol (mg.L -1 )<br />
Acide vanillique (mg.L -1 )<br />
Acide ferrulique (mg.L -1 )<br />
0,32-1,36 D’Annibale et al.,<br />
2004 ; El Hajjouji et<br />
al., 2007<br />
0,19- 0,57 D’Annibale et al.,<br />
2004 ; El Hajjouji et<br />
al., 2007<br />
0,056 El Hajjouji et al.,<br />
2007<br />
37,9-143,34 Fki et al., 2005 ;<br />
Ergül et al., 2009<br />
8,51-9,43 Fki et al., 2005 ;<br />
Ergül et al., 2009<br />
20 Azabou et al., 2007<br />
95 D’Annibale et al.,<br />
2004 ; Fki et al., 2005<br />
La variation de la quantité des composés phénoliques dans les margines citée dans la<br />
littérature, dépend, en plus des conditions citées auparavant (procédé de trituration, variété<br />
d’olives, l’âge des margines), du procédé d’extraction et des techniques d’analyses qualitative<br />
et quantitative des monomères phénoliques (Hamdi 1996; De Marco et al., 2007).<br />
2.1.7.5. Caractéristiques microbiologiques<br />
Dans les margines d’olive, seuls quelques microorganismes arrivent à se développer. Ce<br />
sont essentiellement des levures et des moisissures. Dans la plupart des cas, il y a absence de<br />
microorganismes pathogènes et ils ne posent alors aucun problème de point de vue sanitaire.<br />
Le pouvoir antimicrobien des margines (Ramos-Cormenzana et al., 1996) est lié<br />
essentiellement à l’action exercée par les phénols monomériques et les pigments bruns ou<br />
13
Matériel et Méthodes<br />
catéchol-mélaninique (Hamdi et Ellouz 1993). Ces effluents agissent sur les bactéries en<br />
dénaturant les protéines cellulaires et en altérant les membranes (Ranalli, 1991). Ils peuvent<br />
inhiber également l’activité des bactéries symbiotiques fixatrices d’azote dans le tube digestif<br />
des ruminants en inhibant leur activité enzymatique (Hattenschwiler et Vitousek 2000).<br />
2.2. L’impact des margines d’olive sur l’environnement, traitements<br />
et valorisation<br />
Le rejet des margines d’olive sans traitement est un problème majeur dans les pays<br />
producteurs de l’huile d’olive du bassin méditerranéen. Au Maroc, la production annuelle de<br />
ces effluents est estimée à environ 250 000 m 3 (FODEP, 2004). Ces margines engendrent de<br />
sérieux dégâts environnementaux. L’absence de méthodes de traitements adaptées aux<br />
besoins des unités de trituration pousse les propriétaires à rejeter ces eaux dans la nature sans<br />
aucun contrôle et par la suite à surcharger le réseau d’égouts avec des substances toxiques. La<br />
concentration et la charge en matières organiques ne peut qu’entraîner un disfonctionnement<br />
des stations d’épuration.<br />
2.2.1. L’impact sur les eaux : cas de l’Oued Sebou (Fès)<br />
En plus des désagréments visuels et des mauvaises odeurs, la forte charge organique des<br />
margines détruit totalement la faune et la flore aquatique par absorption de tout ou partie de<br />
l'oxygène dissous dans les eaux. Le déversement de la totalité des margines dans l’oued<br />
Sebou provoque de graves pollutions (Foutlane et al., 2002). Les rejets lors des campagnes de<br />
trituration font augmenter la DBO des eaux du Sebou d'une manière dramatique, au point de<br />
faire chuter à zéro le taux d'oxygène dissous et ce sur plusieurs dizaines de kilomètres<br />
(Fig. 6). La capacité d'autoépuration de l'oued est ainsi annihilée. Conjuguée aux effets de la<br />
salinité des margines déversées dans l’Oued, toute vie aquatique s'en trouve totalement<br />
inhibée.<br />
Pendant la campagne oléicole au Maroc qui dure approximativement d'octobre à février<br />
(4 à 5 mois), la qualité des eaux de l'Oued Sebou en aval de la ville de Fès, se trouve en effet<br />
très dégradée. À titre d'exemple pendant la campagne 1991/92, ses eaux étaient tellement<br />
polluées que l'Office National des Eaux Potables (ONEP) s'est trouvé dans l'obligation<br />
d'arrêter trois stations de traitement de l'eau de l'oued Sebou pour la production de l'eau<br />
potable, et la ville de Fès était sur le point de manquer d'eau potable.<br />
14
Matériel et Méthodes<br />
Figure 6. Evolution de la DCO dans les eaux l’Oued Sebou avant, pendant et après<br />
la période d’activité des huileries (Foutlane et al., 2002).<br />
2.2.2. L’impact sur le sol<br />
La diminution progressive du contenu de substance organique dans les sols,<br />
subordonnée à l´agriculture intensive, est particulièrement préoccupante, étant donné la<br />
décomposition rapide de la substance organique. Les conséquences d’une telle diminution<br />
sont immédiatement identifiables par la dégradation des propriétés physiques des sols<br />
accompagnée de l’augmentation des risques d´érosion. L'utilisation agronomique de la<br />
biomasse résiduelle, comme les sous-produits de l´industrie oléicole, demeure un moyen pour<br />
réintégrer la perte de substance organique, pour recycler de manière correcte les éléments<br />
nutritifs et finalement pour la possibilité d'écouler ces refus au plus bas coût possible.<br />
15
Matériel et Méthodes<br />
Pour une évaluation quantitative des modifications physiques et chimiques des terrains<br />
traités avec les restes de trituration des olives, il est nécessaire de tenir compte de la<br />
considérable variabilité de la composition chimique de ces matériaux, tous caractérisés par un<br />
pH bas et une concentration élevée en sels et en substances organiques contenant des fractions<br />
difficilement biodégradables. En outre, la composition chimique de ces matériaux est même<br />
variable pendant leur stockage soit du fait de la sédimentation partielle de la fraction<br />
insoluble, soit de la transformation microbiologique de la substance organique et l'évaporation<br />
de la composante aqueuse.<br />
2.2.3. Les traitements des margines<br />
Dans la littérature, on rencontre de multiples solutions de traitement des margines<br />
d’olive. Les techniques de traitement des margines sont basées sur différents procédés<br />
physiques, chimiques et/ou microbiologiques. Bien qu’il n’existe pas encore de solution<br />
parfaite permettant d’éliminer la pollution des margines, certains procédés semblent être plus<br />
efficaces que d’autres. Les choix opérés par les décideurs dépendront ainsi des contraintes, du<br />
moment et des facteurs multiples et complexes d'ordres socio-économique, technique et<br />
politique. Le Tableau 5 résume les différents procédés de traitement des margines rencontrés<br />
dans la littérature.<br />
16
Tableau 5. Procédés de traitement des margines.<br />
Procédé Traitement Remarques Références<br />
Thermiques<br />
Physico-<br />
chimique<br />
Biologique<br />
Evaporation<br />
naturelle<br />
Procédé ralenti, grandes<br />
superficies,<br />
Evaporation forcée 40 fois plus importante que<br />
l’évaporation naturelle<br />
Incinération L'énergie nécessaire est<br />
Filtration/<br />
Ultrafiltration<br />
obtenue par combustion du<br />
grignon.<br />
Ozonation 80% de réduction des<br />
Matériel et Méthodes<br />
Ranalli, 1991<br />
Fiestas et Borja,1992<br />
Ranalli, 1991<br />
Réduction de 63% de DCO Pietro et al., 1988<br />
polyphénols<br />
Décoloration 15% de décoloration avec<br />
Traitement<br />
électrochimique<br />
coagulation-<br />
floculation<br />
(CaO) [4 à 35 g.L -1 ]<br />
Oxydation des composés<br />
phénoliques par électrolyse<br />
Andreozzi, 1988<br />
Flouri et al., 1997<br />
Longhi et al., 2001<br />
Scandia vision Jaouani et al., 2005 ;<br />
Achak et al., 2008<br />
électrocoagulation Seam Adelpa Inan et al., 2004 ;<br />
traitement<br />
anaérobie<br />
Prétraitement exigé pour<br />
augmenter le rendement<br />
Khoufi et al., 2008<br />
Hamdi, 1992 ; Azbar et<br />
al., 2008<br />
traitement aérobie Champignons Balice et al., 1988 ; El<br />
2.2.4. La valorisation des margines<br />
Hajjouji et al., 2008<br />
Compte tenu de leur pourcentage en matière organique (10 à 25%) et de leur<br />
composition chimique, les margines seules ne peuvent constituer un produit de valeur ajoutée<br />
intéressante. Enrichies, mélangées à d’autres résidus agricoles, concentrées, séchées et/ou<br />
purifiées, elles peuvent être valorisées et employées pour la production de certains<br />
composants de valeur ajoutée. Au cours des dernières décennies, des études de plus en plus<br />
17
Matériel et Méthodes<br />
nombreuses ont été conduites par plusieurs chercheurs avec la vision de développer<br />
différentes applications de valorisation des margines en vue de limiter leur effet polluant.<br />
2.2.4.1. Extraction des antioxydants (hydroxytyrosol)<br />
En vue de proposer des solutions adéquates, la société Biovalor a investi plus de 2<br />
millions de DH dans une unité sise dans la région de Fès pour l'extraction des antioxydants<br />
existant dans les margines, en particulier l’hydroxytyrosol. Elle est dotée d’un laboratoire<br />
d’analyse, une chaîne de froid pour le stockage des margines, et deux machines de traitement<br />
et de valorisation. Le procédé d’extraction des polyphénols de margines consiste à ajouter un<br />
polysaccharide aux margines, puis à atomiser le mélange pour obtenir une poudre contenant<br />
les composés phénoliques encapsulés à l’intérieur de la structure du polymère. Si cette<br />
opération est coûteuse, elle est néanmoins efficace et permet en même temps de traiter les<br />
margines en récupérant une eau utilisable dans l’agriculture.<br />
Reste à signaler que le traitement des margines de Fès, en grande quantité (10 tonnes<br />
par jour), nécessite au moins 70 millions de Dirham marocain. La rentabilité annuelle est<br />
estimée par Biovalor à 15% du total des investissements. Il serait, en outre, judicieux de<br />
souligner que, par rapport aux autres procédés proposés actuellement, celui-ci donne une<br />
solution efficace et valorisante à ces résidus.<br />
2.2.4.2. Culture de champignons supérieurs<br />
Les champignons, en particulier les basidiomycètes, sont connus pour produire des<br />
enzymes impliquées dans la dégradation de la lignine (Mishra et Leatham 1990; Elisashvili et<br />
al., 2008). Dans cette vision, plusieurs études ont été menées sur la dégradation des<br />
polyphénols des margines par les champignons, en particulier les basidiomycètes et des<br />
ascomycètes de genre d’Aspergillus (Garcia et al., 2000; D'Annibale et al., 2004; Olivieri et<br />
al., 2006; Zenjari et al., 2006; Di Serio et al., 2008). L’arsenal d’enzymes impliquées dans la<br />
décoloration, et la réduction des polyphénols des margines, diffère en fonction de l’espèce de<br />
champignon. Les enzymes les plus rencontrées dans la dégradation des polyphénols sont les<br />
laccases, Mn peroxydase et lignine peroxydase (Sayadi et Ellouz, 1993; D'Annibale et al.,<br />
2004; Silva et al., 2008).<br />
Par ailleurs, il y a peu d’études traitant l’utilisation des margines dans la culture des<br />
champignons comestibles. En effet, différents substrats lignocellulosiques mélangés avec des<br />
margines d’olive à différentes concentrations ont été testés pour la culture de Pleurotus sp.<br />
(Zervakis et al., 1996; Kalmis et Sargin 2004).<br />
18
Matériel et Méthodes<br />
2.3. Le champignon comestible, Lentinula edodes (Berk.) Pegler<br />
2.3.1. Définition des champignons<br />
Les champignons sont définis comme des organismes eucaryotes sporogènes non<br />
chlorophylliens, se reproduisant par voie sexuée ou asexuée. Ils sont à la base de nombreux<br />
procédés biotechnologiques allant de la production d’aliments fermentés comme les fromages<br />
à celle de molécules à haute valeur ajoutée comme les vitamines, les antibiotiques ou les<br />
enzymes (Cohen et al., 2002). Le développement des champignons se fait par croissance<br />
apicale du mycélium dans toutes les directions et de façon identique. Ce mode de croissance<br />
se traduit par la mise en place de colonies circulaires caractéristiques des champignons sur<br />
milieu gélosé (Roussos et Raimbault 1982; Gervais et Bensoussan, 1994).<br />
2.3.2. La physiologie de croissance des champignons<br />
Les champignons macromycètes (champignons supérieurs), par opposition aux<br />
moisissures (micromycètes), présentent un cycle de développement en deux phases<br />
(Alexopoulos et Mims, 1979) : i) la phase haploïde conduisant à la formation de mycélium<br />
capable de coloniser le milieu, ii) la phase dicaryotique conduisant à la formation du<br />
carpophore, communément appelé champignon où se trouvent les basides et les basidiospores.<br />
Pendant la croissance du mycélium, il y a une production de métabolites primaires et<br />
secondaires, de CO2 et une consommation d’O2. De façon générale, le métabolisme des<br />
champignons filamenteux est divisé en métabolisme primaire et secondaire. Le métabolisme<br />
primaire est le résultat de quelques réactions chimiques catalysées par les enzymes qui<br />
procurent aux cellules l’énergie nécessaire pour la production de nouvelles cellules et la<br />
formation de macromolécules indispensables (protéines, polymères structuraux et ADN). À<br />
partir de ce métabolisme, on peut obtenir un grand nombre de molécules d’intérêt industriel<br />
comme l’acide citrique, différents acides organiques, alcools et aussi de nombreuses<br />
d’enzymes comme les cellulases, lipases, etc. (Roussos et Raimbault 1982; Cordova et al.,<br />
1998). Ces métabolites primaires se produisent en général pendant la phase de croissance<br />
exponentielle du mycélium.<br />
Les champignons filamenteux comme Aspergillus, Penicillium, Rhizopus et<br />
Trichoderma sont les microorganismes les mieux adaptés pour la culture en milieu solide<br />
(Roussos, 1985; Oriol, 1987; Cordova et al., 1998). Les champignons saprophytes (Agaricus,<br />
Pleurotus, Lentinula) se développent très bien en milieu solide sur différents substrats<br />
19
Matériel et Méthodes<br />
agricoles (Philippoussis et al., 2001; Royse et Sanchez-Vazquez 2001; Philippoussis et al.,<br />
2003; Philippoussis et al., 2007). Ces microorganismes sont adaptés à certaines conditions de<br />
culture, température, pH, disponibilité en eau et oxygène, et peuvent se développer en culture<br />
solide sur différents substrats agricoles.<br />
Les champignons ectomycorriziens (Lactarius, Pisolithus, Suillus) ont été également<br />
cultivés en FMS, mais des recherches sont encore nécessaires afin de maîtriser leur<br />
domestication pour la production de carpophores (De Araujo et al., 2000). En raison des<br />
grandes potentialités présentées par le mycélium de certains champignons supérieurs, diverses<br />
études ont trouvé des applications de la phase végétative comme le Claviceps (Trejo-<br />
Hernandez, 1992) et les morilles qui développent des arômes caractéristiques (Kabbaj, 1997).<br />
2.3.3. Les techniques générales de culture de L. edodes<br />
La culture de L. edodes s’effectue selon deux méthodes différentes ; culture sur buches,<br />
qui est la culture traditionnelle, et culture sur des substrats synthétiques, qui est la méthode la<br />
plus répandue actuellement. Le principe de la culture, consiste à ensemencer un substrat<br />
donné avec le mycélium du L. edodes qui s’est développé auparavant sur un support à base de<br />
bois ou de grains de céréale (spawn). Par la suite, le substrat inoculé est incubé dans les<br />
conditions favorables de la souche utilisée, pour le développement du mycélium et pour la<br />
formation des carpophores. La technique générale de culture de L. edodes dans les colonnes<br />
de Raimbault est basée sur les travaux de Sobal (2002) et Kabbaj (1997).<br />
2.3.3.1. Culture sur bûches<br />
L’intérêt de cette technique réside dans l’utilisation des installations simples pour sa<br />
réalisation. Par ailleurs, il est recommandé d’appliquer cette technique traditionnelle dans des<br />
régions tempérées et humides avec une variation de température allant de 15 à 25°C (Royse,<br />
2001). Le processus de culture est résumé en 5 étapes ; préparation des buches, inoculation,<br />
incubation, induction et fructification (Figure 7).<br />
20
Figure 7. Processus général de culture de L. edodes sur bûches.<br />
2.3.3.2. Culture sur substrats synthétiques<br />
Matériel et Méthodes<br />
A cause de la limitation géographique du chêne, bois sur lequel se développe<br />
naturellement L. edodes, différents substrats agricoles synthétiques ont été testés pour la<br />
production de L. edodes (Philippoussis et al., 2000 ; 2001). Parmi les avantages de cette<br />
technique, on trouve la possibilité d’utiliser des sous-produits de l’agriculture et des industries<br />
agroalimentaires, la réduction du temps de culture, l’augmentation de l’efficience biologique<br />
et la possibilité d’avoir une production indépendante des conditions climatiques (Mata, 1997).<br />
Mais ce processus de culture nécessite des installations sophistiquées, une bonne maitrise de<br />
la technologie et de la biologie du champignon. Les étapes de la culture sont les<br />
suivantes (Mata, 1997) :<br />
a) Préparation du substrat. Le substrat pour la culture de L. edodes est obtenu à partir<br />
d’un mélange de produits lignocellulosiques, de suppléments nutritifs et d’eau. Le<br />
champignon L. edodes est capable de se développer sur une gamme diverse de substrats<br />
lignocellulosiques qui ne nécessitent pas de traitement de compostage au préalable. Toutefois,<br />
la sciure du bois a été utilisée comme substrat de culture de L. edodes.<br />
21
Matériel et Méthodes<br />
b) Traitement thermique. Pour éviter toute contamination du substrat par d’autres<br />
communautés microbiennes compétitrices, deux procédés de traitement sont utilisés:<br />
i) La stérilisation élimine tous les microorganismes présents dans le substrat. Elle est<br />
utilisée pour les substrats fortement supplémentés. Le substrat est conditionné dans des sacs<br />
plastiques et stérilisé en autoclave à 121°C pendant 1 heure.<br />
ii) La pasteurisation consiste à maintenir dans le substrat une température constante,<br />
autour de 65°C pendant au moins 24 h.<br />
c) Inoculation ou lardage. L’inoculation avec le blanc (spawn) est une étape très<br />
importante de la culture. Pour les substrats stérilisés, l’inoculation s’effectue dans un local<br />
stérile avec de l’air filtré. Par contre, pour un substrat pasteurisé, le substrat peut être inoculé<br />
juste dans un endroit propre. Le taux d’inoculation varie en fonction du procédé thermique<br />
utilisé, du substrat, et de la souche. En général un apport de 3 à 10% est pratiqué (Royse et<br />
Sanchez-Vazquez, 2001; Zhang et al., 2002; Royse et Sanchez-Vazquez, 2003).<br />
d) Incubation. Les conditions nécessaires pour le développement du mycélium sont la<br />
température, l’oxygénation et les périodes d’éclairage. Ce dernier est indispensable durant la<br />
période de formation des sporophores. La durée d’incubation varie entre 1 et 3 mois selon la<br />
souche utilisée et la nature du substrat (Philippoussis et al., 2001; Philippoussis et al., 2003;<br />
Royse et Sanchez-Vazquez 2007; Shen et al., 2008).<br />
e) Induction de fructification. Apres l’envahissement total du substrat, le mycélium<br />
forme de petites agrégations à l’origine des primordia. A ce stade, il faut enlever le sac<br />
plastique et placer le bloc dans une sale éclairée, humidifiée (85-90%) et à une température<br />
entre 15-18°C. L’application d’un choc thermique est pratiquée pour certaines souches.<br />
f) Production et récolte. Durant la fructification, les conditions d’induction sont<br />
maintenues. La maturité des carpophores est atteinte après 10-15 jours de leur apparition.<br />
2.3.3.3. Culture sur substrats alternatifs<br />
Dans la littérature, plusieurs formules de substrats ont été testées pour remplacer la<br />
culture de L. edodes sur les buches. La sciure de bois de chêne a été utilisée pour la culture de<br />
base de L. edodes (Levanon et al., 1993; Royse et Sanchez-Vazquez 2007). Par ailleurs des<br />
substituts du bois, riches en lignocellulose, permettant un meilleur rendement en L. edodes ont<br />
été testés (Zervakis et al., 1996; Curvetto et al., 2002a,b; Özçelik et Peksen 2007).<br />
La formulation des substrats dédiés à la culture de L. edodes nécessite de tenir compte<br />
de leur composition chimique. La composition des résidus lignocellulosiques varie selon leur<br />
nature et leur structure. Ainsi, la teneur en lignine est un facteur qui limite la croissance des<br />
22
Matériel et Méthodes<br />
champignons. La décomposition d’un résidu lignocellulosique comprend au moins deux<br />
étapes selon Chalaux (1993) ; tout d’abord, les sucres solubles et une partie de la cellulose et<br />
des hémicelluloses sont dégradés; ensuite, la dégradation des composés lignifiés est réalisée.<br />
Selon ce modèle, la vitesse de transformation des résidus lignocellulosiques est<br />
essentiellement conditionnée par la composition des substrats. Cependant, d’autres facteurs<br />
affectent la décomposition de ces matériaux, en l’occurrence la température, l’humidité, et la<br />
qualité du matériel.<br />
2.3.4. Les phénoloxydases, les laccases<br />
Les phénoloxydases (PO) jouent un rôle polyvalent dans la transformation des produits<br />
agricoles, puisqu’elles sont impliquées dans la minéralisation de composés phénoliques<br />
récalcitrants naturels, tels que la lignine, et dans la formation des substances humiques<br />
(Widsten et Kandelbauer, 2008). Les PO utilisant l’O2 dans leur cycle catalytique<br />
comprennent deux classes d’enzymes: les laccases et les tyrosinases (Durán et Esposito,<br />
2000).<br />
2.3.4.1. Structure des laccases<br />
Les laccases (E.C. 1.10.3.2 benzenediol: oxygen oxidoreductase) sont des<br />
métalloenzymes contenant des atomes de cuivre au niveau de leurs sites actifs respectifs; elles<br />
assurent simultanément l’oxydation de composés phénoliques et la réduction de l’O2 (Enguita<br />
et al., 2003). Les atomes de cuivre présents dans la structure des laccases sont de trois types.<br />
• Le cuivre de type 1 confère à la molécule sa couleur bleue et présente un pic<br />
d’absorption à 610 nm.<br />
• Le cuivre de type 2 n’a pas d’absorption dans le visible.<br />
• Les 2 atomes de cuivre de types 3 présentent une absorption à 330 nm.<br />
2.3.4.2. Mécanismes d’action des laccases<br />
Le mécanisme d’action général des laccases consiste en une réaction d’oxydation de<br />
composés aromatiques, faisant intervenir quatre électrons, couplée à la réduction de l’oxygène<br />
moléculaire en eau. Au niveau du site réactionnel, le cluster formé par les atomes de cuivre de<br />
type 2 et 3 est crucial pour la réduction de l’oxygène. L’oxydation des substrats aboutit à la<br />
formation d’un radical actif qui peut subir d’autres réactions non enzymatiques :<br />
i) Polymérisation des monomères : l’oxydation enzymatique des composés phénoliques<br />
conduit à la formation des intermédiaires qui sont des radicaux libres qui réagissent entre eux<br />
23
Matériel et Méthodes<br />
pour former des dimères, des oligomères ou des polymères couplés avec des liaisons<br />
covalentes C-C, C-O ou C-N.<br />
ii) dégradation des polymères : les laccases sont impliquées dans la dégradation de<br />
polymères naturels, tels que la lignine, les acides humiques (Claus et Filip 1998). En effet, la<br />
formation des radicaux produits conduit à la rupture de liaisons covalentes et par la suite à la<br />
libération des monomères. La dépolymérisation peut avoir lieu également indirectement, par<br />
l’intermédiaire des médiateurs, qui sont des composés organiques ou des métaux oxydés par<br />
l’action des laccases tels que l’alcool vératrylique ou les atomes de manganèse. Dans les<br />
procédés biotechnologiques, les médiateurs sont souvent utilisés pour accroître le potentiel<br />
d’oxydation des laccases (Claus et al., 2002; Farnet et al., 2004).<br />
2.3.4.3. Applications des laccases<br />
Les laccases sont des enzymes d’intérêt majeur dont la régulation est associée à la<br />
morphogenèse du corps fructifère chez les basidiomycètes et leur production est influencée<br />
par la qualité du substrat de culture (Lechner et Papinutti 2006). Elles semblent aussi être<br />
impliquées dans les processus de production de la biomasse mycélienne (Mansur et al., 2003)<br />
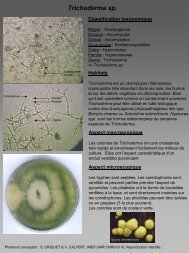
et d’inhibition du développement des moisissures comme les Trichoderma (Velazquez-<br />
Cedeño et al., 2008). Les laccases ont montré une grande capacité à l’oxydation d’un large<br />
spectre de molécules phénoliques (Durán et Esposito 2000). Elles sont utilisées pour<br />
délignifier des complexes lignocellulosiques et améliorer la production du bioéthanol obtenu<br />
à partir de ces complexes. Elles sont employées dans différents procédés de bioremédiation<br />
pour protéger l’environnement de dommages créés par des effluents industriels (Mayer et<br />
Staples 2002). Les laccases fongiques sont capables de décolorer l’eau de teinture des textiles<br />
(Eichlerova et al., 2006; Zouari-Mechichi et al., 2006). Après la récolte des champignons, le<br />
compost résiduel est une source potentielle de laccases et qui pourrait être valorisée par<br />
différents usages (Trejo-Hernandez et al., 2001). Les laccases ont été utilisées aussi pour le<br />
traitement des margines d’olive qui seraient utilisées comme amendement des sols<br />
(D'Annibale et al., 2000; 2004).<br />
2.4. Les fermentations en milieux solides<br />
Les fermentations en milieu solide (FMS) ont été pratiquées depuis très longtemps de<br />
façon artisanale en alimentation humaine traditionnelle. Elles concernent essentiellement les<br />
transformations de produits agricoles par les champignons inférieurs. En occident, les FMS<br />
sont devenues des techniques essentielles pour la fabrication de produits fermentés tels que le<br />
24
Matériel et Méthodes<br />
fromage. Par contre dans les pays asiatiques, ces fermentations sont utilisées essentiellement<br />
dans la fabrication des produits à base de protéines végétales comme celle issues du soja<br />
(Raimbault, 1980).<br />
2.4.1. Définition des FMS<br />
La FMS est définie comme étant la croissance de micro-organismes sur des substrats<br />
solides en absence totale ou presque d’eau libre (Raimbault, 1980, Pandey et al., 2000). Sa<br />
traduction en anglais est Solid State cultivation ou Solid Substrate Fermentation ou encore<br />
Solid State Fermentation. Hesseltine (1987) l’a définie comme étant, une fermentation dans<br />
laquelle le substrat n’est pas soluble dans une phase liquide.<br />
Selon d’autres chercheurs (Raimbault, 1980; Lonsane et al., 1985; Oriol, 1987; Trejo-<br />
Hernandez, 1992; Saucedo-Castañeda, 1995), la FMS est une culture microbienne qui se<br />
développe en surface et à l’intérieur d’une matrice poreuse et en absence de tout écoulement<br />
liquide. La matrice poreuse peut être constituée d’un substrat humide ou d’un support inerte<br />
capable d’absorber les nutriments qui se trouvent à l’état dissous. D’après ces définitions, les<br />
auteurs s’accordent à définir la FMS comme étant une fermentation ou une culture de micro-<br />
organismes sur un milieu ou substrat solide en absence d’eau libre. En fonction de la nature<br />
du support, on peut distinguer deux types de fermentations en milieu solide (Raimbault,<br />
1980 ; Roussos, 1985, Saucedo-Castañeda, 1995) :<br />
i) Culture solide sur une phase substrat-support qui est constituée d’un matériau<br />
assurant une double fonction : support et source de nutriments.<br />
ii) Culture solide avec une phase support constituée d’un matériau inerte, imprégné d’un<br />
milieu liquide nutritif.<br />
Dans la FMS, la capacité d’absorption d’une culture liquide est un facteur important à<br />
contrôler. Les matériaux utilisés sont divers et la plupart ont une forte capacité de rétention<br />
d’eau tels que la bagasse de canne à sucre, l’écorce de bois ou la mousse de polyuréthane<br />
(Contreras-Domínguez, 2007; Hassouni, 2007).<br />
2.4.2. Les principaux paramètres des FMS<br />
Le développement de microorganismes sur différents substrats ou supports solides<br />
dépend principalement des paramètres suivants ; la quantité d’inoculum, l’humidité et<br />
l’activité de l’eau, l’aération et le transfert d’oxygène, la régulation de la température et du<br />
pH. Pour mesurer la croissance du microorganisme en FMS, on peut faire appel à la<br />
respirométrie, aux analyses de la biomasse avec différentes méthodes indirectes<br />
25
Matériel et Méthodes<br />
(protéines, chitine, ergostérol, acides nucléiques) ou à la production de différents métabolites<br />
(Durand, 2003).<br />
2.4.2.1. Microorganismes et l’inoculation du milieu solide<br />
Les champignons filamenteux, contrairement aux organismes unicellulaires, sont les<br />
mieux adaptés à la FMS en raison de leur capacité à la colonisation des substrats par<br />
l’émission de filaments. Dans les procédés de FMS, on utilise deux types d’inoculation : la<br />
microflore endogène du substrat ou des ferments contrôlés (Raimbault et Alazard, 1980;<br />
Roussos et al., 1989). Dans les procédés de compostage et d’ensilage, on utilise la microflore<br />
naturelle, alors que dans les procédés de production d’enzymes et d’aliments fermentés, on<br />
utilise des ferments avec une ou plusieurs souches, cultures mixtes, (Perraud-Gaime, 1995).<br />
2.4.2.2. Choix du support/substrat<br />
Le support solide doit combler les besoins nutritifs du microorganisme, mais aussi<br />
assurer quelques impératifs physiques essentiels au bon déroulement de la FMS comme le<br />
transfert de chaleur et de masse, la granulométrie, la porosité, la capacité d’absorption de<br />
l’eau et la résistance à la compression et à l’agitation (Raimbault, 1980). La sélection du<br />
substrat dépend de son coût et de sa disponibilité. Sur le plan économique, il faut considérer le<br />
prix du milieu, le coût du fonctionnement de la fermentation et la valeur du produit final. Le<br />
plus économique est d’utiliser des substrats naturels comme les résidus de l’agriculture ou de<br />
l’agro-industrie. Grâce à la FMS, de nombreuses recherches ont été faites dans le but<br />
d’assurer une valorisation des sous-produits industriels pour différentes finalités telles que la<br />
détoxication, l’enrichissement en protéines microbiennes, ou la production de produits à forte<br />
valeur ajoutée (Bellon-Maurel et al., 2003).<br />
2.4.2.3. Activité de l’eau et l’humidité<br />
Le rôle de l’eau dans le processus de FMS est multiple. Elément dominant dans la<br />
composition de la biomasse, l’eau sert à véhiculer les enzymes et les composés nutritifs, et<br />
facilite les interchanges gazeux. La quantité maximale de liquide présente dans la phase solide<br />
est fonction de la capacité de rétention d’eau du substrat ou du support solide. Toutefois, cette<br />
quantité de liquide doit être suffisante à la croissance des microorganismes sans détruire la<br />
structure solide ou réduire la porosité du substrat ou du support (Oriol, 1987).<br />
Le développement microbien en FMS nécessite une humidité du substrat supérieure à<br />
50 %. Une humidité très élevée dans le substrat provoque une diminution de la porosité du<br />
substrat, une baisse de la diffusion d’oxygène et une augmentation du risque de la<br />
26
Matériel et Méthodes<br />
contamination. En FMS, du fait de son faible taux d’humidité, le substrat se comporte comme<br />
un milieu sélectif qui favorise le développement des champignons filamenteux puisque la<br />
croissance des bactéries et des levures nécessite des activités de l’eau plus élevées<br />
(0.85
2.4.2.5. Contrôle de la température pour les FMS<br />
Matériel et Méthodes<br />
Les champignons filamenteux utilisés dans les procédés de FMS sont souvent<br />
mésophiles et se développent à des températures optimales comprises entre 25 et 30°C<br />
(Roussos, 1985). Cependant, la production de chaleur métabolique au cours de la FMS cause<br />
une augmentation importante de la température dans les bioréacteurs et si cette chaleur n’est<br />
pas éliminée rapidement du milieu de culture, elle peut causer l’arrêt de la croissance des<br />
microorganismes. L’élévation de la température dans les réacteurs de FMS s’explique par la<br />
faible teneur en eau, l’absence d’agitation dans les fermenteurs statiques et la faible<br />
conductivité thermique des matériaux biologiques.<br />
L’augmentation excessive de la température est nocive pour la croissance des<br />
microorganismes, la production et la stabilité des produits d’intérêt. Il faut donc agir sur la<br />
culture afin de diminuer cette augmentation de la température. Différentes stratégies ont été<br />
proposées pour la régulation de la température. Le passage de l’air à travers le fermenteur<br />
peut réguler simultanément la température et l’humidité (Lonsane et al., 1992). Ainsi,<br />
l’aération et l’évaporation de l’eau sont utilisées constamment pour contrôler<br />
automatiquement l’humidité et la température dans les processus de FMS (Saucedo-Castañeda<br />
et al., 1995). D’autres moyens peuvent également être utilisés, comme les bains thermostatés,<br />
chambres thermorégulées ou doubles enveloppes (Bagnon, 1999). Une nouvelle stratégie a été<br />
proposée en utilisant des microorganismes thermophiles ou thermorésistants (Cordova et al.,<br />
1998). En conclusion, le problème de production excessif de chaleur au cours de la FMS ne se<br />
pose pas dans le cas des champignons supérieurs qui ont un métabolisme assez lent,<br />
faiblement exothermique et facilement contrôlable. De ce fait, très peu d’études ont été<br />
réalisées pour le transfert de chaleur au cours de la culture du mycélium des champignons<br />
supérieurs (Van Lier et al., 1994; Sobal, 2002).<br />
2.4.2.6. Evolution du pH<br />
L’activité métabolique des micro-organismes provoque une variation du pH du substrat<br />
de culture influençant par la suite la bonne croissance des micro-organismes. Les<br />
champignons filamenteux se développent bien dans des milieux acides et peuvent tolérer<br />
d’importants changements de pH (2,5 jusqu’à 7,5). Cependant le pH initial d’une culture doit<br />
être ajusté à son optimum par l’ajout d’une solution nutritive tamponnée. Le maintien du pH<br />
stable au cours de la fermentation reste une tâche très difficile à réaliser en particulier pour les<br />
FMS non agitées (Saucedo-Castañeda, 1995). L’utilisation des tampons permet de résoudre ce<br />
problème. Par ailleurs, Raimbault (1980) et Roussos (1985) ont démontré qu’un mélange de<br />
28
Matériel et Méthodes<br />
sulfate d’ammonium et d’urée confère au milieu de culture un pouvoir tampon et permet donc<br />
de maintenir le pH à des valeurs favorables à la croissance des micro-organismes.<br />
2.4.3. Les applications des FMS<br />
Les applications des FMS pour la valorisation des sous-produits agro-industriels sont<br />
nombreuses et variées (Tableau 6). Les domaines d’application des FMS sont chaque jour<br />
plus variés. Elle a été employée dans plusieurs domaines de recherche ; dépollution des sols,<br />
biodégradation de composés à risque, détoxication de résidus agro-industriels,<br />
biotransformation des cultures et de leurs résidus pour améliorer leurs qualités nutritionnelles<br />
(compenser les manques en vitamines, minéraux ou protéines).<br />
Tableau 6 : Exemples de substrats utilisés et les applications en FMS (Bellon-Maurel et al.,<br />
2003).<br />
Substrats Applications Substrats Applications<br />
L’application de la FMS dans les Bio-raffineries (biogaz, bioéthanol) a révélé l’intérêt<br />
de la FMS, qui apporte par la suite une valorisation aux déchets agricoles (Sukumaran et al.,<br />
2005 ; Singhania et al., 2007). Par ailleurs, la production de cellulase par la FMS sur des<br />
résidus agro-industriels pour la fabrication des biocarburants connaît une demande accrue.<br />
Ainsi, par l’emploi de la FMS, la production d’hydrogène, d’acides organiques, d'éthanol et<br />
de biodiesel a été concrétisée avec succès en utilisant soit des substrats solides soit des<br />
supports solides (Hama et al., 2007; Wu et al., 2007; Jo et al., 2008).<br />
29
2.4.4. Les avantages et les inconvénients de la FMS<br />
Matériel et Méthodes<br />
Le Tableau 7 contient une liste non exhaustive des avantages et des inconvénients de la<br />
fermentation en milieu solide par rapport à la fermentation en milieu liquide.<br />
Tableau 7. Comparaison de la fermentation en milieu liquide (FML) avec la fermentation en<br />
milieu solide (FMS) (D’après Sobal, 2002).<br />
Substrat<br />
FML<br />
FMS<br />
Conditions aseptiques Stérilisation par choc thermique Stérilisation ± obligatoire<br />
Eau Grands volumes consommés Consommation limitée<br />
Chaleur métabolique Contrôle facile de la température Transfert de chaleur faible<br />
Aération Limitée à l'oxygène soluble, haut<br />
niveau d'air requis<br />
Aération facile, surface<br />
d'échange air/substrat<br />
Contrôle du pH Facile Substrat solide tamponné<br />
Agitation mécanique Bonne homogénéisation Conditions statiques<br />
Inoculation Procédé continu Par spores ou mycélium<br />
Contamination Risque pour une seule souche de<br />
bactérie<br />
Risques si le champignon<br />
pousse lentement<br />
Energie Beaucoup d'énergie consommée Peu d'énergie consommée<br />
Volume de l'équipement Grand volume, grand coût Petit volume, petit coût<br />
Effluents, pollutions Grands volumes d'effluents pollués Pas d'effluents, pas de<br />
pollution<br />
Culture Pour tous les microorganismes Surtout les champignons<br />
2.5. Conclusion de l’analyse bibliographique<br />
(conditions naturelles)<br />
La synthèse bibliographique sur les principaux mots clés de la thèse, a permis en<br />
premier temps, de définir le secteur oléicole au Maroc qui se caractérise par la coexistence de<br />
plusieurs types d’unités de trituration dominées par les maâsras avec le procédé triphasique<br />
fonctionnant avec le système de presse (16 000 d’unités). En plus du bois de la taille (feuilles<br />
et les brindilles des oliviers), le procédé triphasique génère un co-produit solide (grignons) et<br />
un déchet liquide (margines). Les différents substrats oléicoles ont été étudiés<br />
individuellement afin de trouver des solutions de traitement et de valorisation.<br />
30
Matériel et Méthodes<br />
Les perspectives de certaines études ont mis l’accent sur l’utilisation des substrats de<br />
l’industrie oléicole pour la culture de champignons comestibles. C’est dans cette vision que<br />
notre projet de thèse s’inscrit, avec l’originalité de proposer un procédé qui permet la<br />
valorisation de la biomasse totale de l’industrie oléicole, incluant le bois de taille, par la<br />
culture de L. edodes en fermentation en milieu solide.<br />
En deuxième temps, les différentes études menées sur la culture de L. edodes ont mis en<br />
lumière les différentes capacités de ce champignon vis-à-vis de la dégradation des composés<br />
récalcitrants, par la synthèse d’un arsenal enzymatique varié et qui dépend de plusieurs<br />
paramètres de culture. Par ailleurs, d’autres études ont été focalisées sur la description et<br />
l’application des laccases dans différents domaines de traitement et de valorisation.<br />
En troisième temps, le procédé de fermentation en milieu solide a été discuté en mettant<br />
l’accent sur les différents paramètres influençant le procédé. Cette partie a soulevé<br />
l’importance de cette technique pour la culture du mycélium de champignons comestibles et<br />
médicinaux, en l’occurrence la culture de L. edodes, et qui est en accord avec la nature du<br />
substrat utilisé pour l’étude.<br />
Finalement, à partir de ces données bibliographiques, le chapitre suivant sur le Matériel<br />
et Méthodes a pu être établi, en particulier les techniques de culture du mycélium de<br />
basidiomycètes sur milieux gélosés munis de feuilles de cellophane permettant la séparation<br />
de la biomasse mycélienne du milieu gélosé épuisé. La technique de mesure de la biomasse<br />
mycélienne par l’analyse de l’ergostérol a permis d’estimer indirectement la quantité du<br />
mycélium présent dans le produit fermenté. Les résultats des mesures des polyphénols et des<br />
activités enzymatiques ont été comparés à ceux décrits dans la littérature.<br />
31
Matériel et Méthodes<br />
32
3. MATERIEL ET METHODES<br />
3.1. Les substrats oléicoles de culture<br />
Matériel et Méthodes<br />
Les échantillons ont été collectés dans une unité semi industrielle située dans la<br />
province de Béni Mellal situé dans la région de Tadla-Azilal (Fig. 8). Avec une superficie<br />
oléicole de 47.650 hectares, la région assure 20% de la production nationale d’olives. En<br />
2008, la production d’olive a atteint 112.550 tonnes d'après l'ORMVAT (Office Régional de<br />
la Mise en Valeur Agricole à Tadla). A titre d'exemple, la production d’huile d’olive en 2003-<br />
2004 a atteint 16.600 tonnes et en 2006-2007, 8550 tonnes. Le secteur de trituration est<br />
dominé par les maâsras traditionnelles et semi industrielle avec des capacités de trituration<br />
très variables. L'unité modèle choisie pour effectuer notre échantillonnage est un type des<br />
maâsras qui assurent près de 50% de la production nationale en huile d’olive.<br />
Figure 8. Localisation de la région Tadla Azilal au Maroc.<br />
3.1.1. La description de l'unité de trituration semi industrielle<br />
L'unité de trituration des olives au Maroc est d’une taille semi industrielle avec le<br />
procédé de presse (Fig. 9). Les olives sont broyées par des meules en granit agitées par un<br />
moteur électrique. Une fois la pâte d’olive obtenue homogène, elle est placée dans la presse<br />
pour l'extraction de l'huile d'olive mélangée avec l'eau végétale. La pression appliquée sur la<br />
pâte d’olive monte jusqu'à 400 kg.cm -2 . La séparation de l'huile des margines est faite par une<br />
décantation suivie d'une centrifugation. La capacité moyenne de trituration de ces unités est<br />
de l'ordre de 6 tonnes par jour.<br />
33
Figure 9. Unité de trituration semi industrielle au Maroc.<br />
3.1.2. L’origine du substrat<br />
Matériel et Méthodes<br />
Les échantillons de margines et de grignons d’olive ont été obtenus à partir d'un lot<br />
d'olives destinées à la trituration. Les olives sont de la variété picholine marocaine de maturité<br />
avancée (Fig.10). Le lot d'olives est de 600 kg avec une maturité avancée. Après l'extraction<br />
de l'huile d'olive par le procédé, on a obtenu 127 litres d'huile, 220 litres de margines et 253<br />
kg de grignons d'olive avec une humidité de 35%.<br />
Figure 10 . Olives de la variété picholine de maturité avancée.<br />
Toute la quantité de grignons d’olive a été placée dans des sacs de 30 kg, et les<br />
margines ont été reparties dans des bidons de 10 litres. Le bois de la taille des oliviers a été<br />
fourni par le propriétaire de l'unité.<br />
34
3.1.3. La préparation et le conditionnement du substrat<br />
Matériel et Méthodes<br />
Après 24 heures de récupération des échantillons, les margines qui sont restées intactes<br />
ont été homogénéisées, et réparties de nouveau dans les bidons de 10 litres. Les margines ont<br />
été stockées dans un congélateur à -20°C jusqu'à leur départ vers Marseille.<br />
Les grignons d’olive ont subi un séchage sous serre pour diminuer leur humidité de<br />
35% à 10%, pour assurer leur conservation. Les grignons d’olive ont été séchés sous serre à<br />
l'obscurité. Un contrôle de l'humidité a été fait chaque jour pour atteindre 10% d'humidité. Le<br />
conditionnement des grignons séchés a été fait dans des sacs en plastique.<br />
3.2. Les souches utilisées<br />
Les 16 souches de champignons supérieurs comestibles de L. edodes utilisées dans notre<br />
étude proviennent de la collection du Laboratoire de recherche « Athens Mushroom<br />
Research Laboratory, AMRL » dirigé par le Dr. Philippoussis en Grèce ainsi que de la<br />
collection du laboratoire de Mycologie et Sécurité des Aliments de l’INRA-Bordeaux dirigé<br />
par le Dr. Savoie. La liste complète des souches utilisées dans ce travail est reportée dans le<br />
Tableau 8.<br />
Tableau 8. Liste des espèces et origine des souches de champignons comestibles étudiées.<br />
Souches Origine Provenance<br />
Le 118 AMRL* Chine<br />
Le 119 AMRL Chine<br />
Le 120 AMRL Chine<br />
Le 121 AMRL Europe<br />
Le 122 AMRL Europe<br />
Le 123 AMRL Europe<br />
Le 124 AMRL Taïwan<br />
Le 125 Souche commerciale Osaka Japon<br />
Le 126 Souche commerciale Osaka Japon<br />
Le 127 Souche commerciale France Mycélium<br />
Le 128 Souche commerciale France Le Champion<br />
Le 129 Souche commerciale Belgique Mycelia<br />
Le 130 Souche commerciale USA, Lamber spawn<br />
Le 131 Souche commerciale Japon<br />
Le 132 Souche hybride INRA France<br />
Le 133 Homocaryon d’une souche commerciale Le Lion France<br />
35
3.3. Les milieux de culture<br />
3.3.1. Les milieux d'entretien<br />
Matériel et Méthodes<br />
Les souches ont été repiquées régulièrement et conservées sur milieu PDA (Sigma) et<br />
PDA avec 10% des margines. Trente neuf grammes de Patate Dextrose Agar (PDA) ont été<br />
dissous dans un litre d’eau distillée en absence ou présence de margines. Le milieu a été porté<br />
à ébullition et stérilisé à 120°C pendant 20 min et distribué en boîtes de Petri (9 cm) à raison<br />
de 20 mL par boîte. Les cultures ont été incubées à 25°C jusqu'à la colonisation complète de<br />
la surface gélosée et ensuite conservées à 4°C pendant 4-6 mois. Le mycélium de différents<br />
âges, obtenu dans ces conditions, a été utilisé pour inoculer le milieu Ebly® (grains de blé<br />
achetés auprès du supermarché Casino, France) et la culture en milieu liquide.<br />
3.3.2. Les milieux gélosés de criblage des souches<br />
Le même milieu d’entretien PDA (Sigma, France) qui a été utilisé pour conserver les<br />
souches et pour produire l’inoculum, a été également utilisé pour caractériser les souches en<br />
mesurant la croissance apicale, la morphologie de la colonie et la biomasse produite. De la<br />
même façon, le milieu PDA + 10% de margines a été préparé en ajoutant 39 g.L -1 de PDA à<br />
un mélange de 100 mL de margines et 900 mL d’eau et il a été aussi utilisé pour caractériser<br />
les souches. Le milieu ME (Extrait de Malt, 24 g.L -1 ) a été utilisé pour réaliser des cultures en<br />
milieu liquide.<br />
3.3.3. Les milieux à base de céréale pour la culture Spawn 1<br />
Le blé de marque Ebly® (Casino, France) a été utilisé comme substrat pour la<br />
production d’inoculum et la culture mycélienne de L. edodes en FMS. Il s’agit d’un produit<br />
standard commercialisé en grande surface. Deux types de Spawn ont été préparés, le Spawn<br />
blanc et le Spawn gris. La préparation du substrat a été réalisée de la manière suivante : dans<br />
un saladier on a introduit 250 mL d’eau, sans ou avec 10% de margines, portée à ébullition,<br />
une quantité de 250 g d’Ebly® a été ajoutée pour préparer le Spawn blanc et gris<br />
respectivement. On a laissé reposer le mélange pendant 5 min pour assurer l’absorption totale<br />
de l’eau. Ensuite, l’Ebly® humidifié a été réparti dans des récipients à fermeture hermétique<br />
et stérilisé à 120°C pendant 30 min.<br />
1 Dans notre étude, on appelle Spawn tout mycélium cultivé sur les grains de céréales.<br />
36
3.4. Les stratégies d'inoculation<br />
Matériel et Méthodes<br />
Pour initier la culture des champignons supérieurs au laboratoire, on utilise toujours du<br />
mycélium obtenu sur différents milieux de culture (Sobal, 2002). Cet inoculum peut provenir<br />
d’un milieu gélosé ou de grains de céréales colonisés pour une inoculation industrielle à<br />
grande échelle. La taille de l’inoculum a été exprimée soit en nombre de carrés de milieu<br />
gélosé déposés sur le milieu gélosé, soit en pourcentage (% ; poids/poids) pour le spawn<br />
introduit dans les colonnes de Raimbault contenant le substrat.<br />
3.4.1. L’inoculation par carrés de milieux gélosés<br />
À l’aide d’un emporte pièce et d’un pic stériles, un disque de gélose PDA a été prélevé<br />
dans la partie où le mycélium du champignon est le plus jeune. Ce morceau de mycélium a été<br />
ensuite utilisé pour ensemencer la gélose vierge d’une boîte de Petri ou pilulier pour assurer le<br />
repiquage et la conservation des souches. Les morceaux de gélose ont été aussi utilisés pour<br />
l’inoculation des grains de blé pour la préparation du spawn, et pour l’inoculation des<br />
Erlenmeyers pour la culture en milieu liquide. L’incubation a été réalisée dans une étuve à<br />
25±2°C pour les milieux solides, alors que les Erlenmeyers sont incubés sous agitation de<br />
100-120 rpm à la même température.<br />
3.4.2. L’inoculation sur feuille de cellophane<br />
Pour l’étude physiologique des souches, on a adopté la technique de papier cellophane<br />
(De Araujo et al., 2000). Cette technique permet la séparation du mycélium, pour déterminer<br />
la biomasse et sa composition chimique, du milieu de culture qui sert à déterminer les<br />
caractéristiques physiologiques du mycélium. Cette technique est décrite en détail ci-après.<br />
Préparation des feuilles de cellophane: les disques de film de cellophane d’un<br />
diamètre légèrement inférieur de celui de la boîte de Pétri ont été coupés en série. Ils ont été<br />
par la suite placés dans un bocal remplie d’eau, afin d’éviter le dessèchement des feuilles de<br />
cellophane lors de l’autoclavage. La stérilisation a été faite à 120°C pendant 20 min. Une fois<br />
les disques refroidis, ils ont été placés à la surface de la gélose, et laissés pendant 30 min pour<br />
assurer une bonne adhésion de la feuille de cellophane.<br />
Inoculation des cultures sur cellophane et incubation: Le mode d’inoculation de la<br />
surface gélosée a été le même que pour la production d’inoculum en utilisant du milieu PDA.<br />
Un carré d’inoculum gélosé contenant du mycélium est déposé au centre de la boîte de Petri<br />
sur un disque de cellophane préalablement placé stérilement sur la gélose. Les cultures ont été<br />
incubées à 25°C pendant 21 jours.<br />
37
Matériel et Méthodes<br />
Mesure du poids sec de la biomasse: Après une période d’incubation, les cultures ont<br />
été arrêtées pour estimer la biomasse produite, le mycélium ainsi que la feuille de cellophane<br />
ont été prélevés et introduits dans une coupelle, puis séchés à 105°C afin de déterminer le<br />
poids sec de la biomasse du champignon (Fig.11).<br />
Figure 11. Technique pour le suivi de la croissance apicale du mycélium et la mesure de<br />
biomasse des champignons en milieu gélosé (De Araujo et al., 2000).<br />
3.4.3. L’inoculation par grain de céréales<br />
Il s’agit d’utiliser une masse de grains de céréales colonisés par le mycélium du<br />
champignon pour ensemencer une masse supérieure de grains de céréales stériles. Afin de<br />
savoir à quel taux on réalise l’inoculation grain à grain, il est nécessaire de peser la quantité<br />
d’inoculum et de milieu (% poids/poids frais). Une fois pesés, les grains colonisés par le<br />
mycélium ont été soigneusement séparés individuellement et ajoutés stérilement au nouveau<br />
milieu ; on a agité doucement afin d’assurer une inoculation homogène, sachant qu’une<br />
38
Matériel et Méthodes<br />
agitation trop violente abîmerait le mycélium et diminuerait aussi sa vigueur. Pour nos<br />
expériences, les substrats ont été inoculés avec différentes quantités d'inoculum (5, 10, et 20<br />
% poids frais) à base d'Ebly® colonisé.<br />
suivante :<br />
3.5. La mise en œuvre des FMS<br />
3.5.1. La préparation du substrat oléicole<br />
La préparation des différents substrats utilisés dans l’étude a été réalisée de la façon<br />
• Le bois de la taille de l’olivier est composé des feuilles et des brindilles et<br />
branches de l’olivier; il a été broyé et tamisé selon la taille des particules désirées.<br />
• Les grignons d’olive ont été mécaniquement homogénéisés.<br />
• Les margines d’olive ont été décongelées juste avant leur utilisation, et<br />
conservées à 4°C pendant une semaine.<br />
Apres l’homogénéisation du bois de la taille avec les grignons d’olive selon différentes<br />
portions, le mélange a été humidifié avec les margines à différentes concentrations (25-50-75-<br />
100%). On a autoclavé, par la suite, l’ensemble des mélanges à 121°C pendant 30 min.<br />
L’humidité du substrat utilisé pour la FMS a été fixée à 50% (p/p).<br />
3.5.2. Les cultures en Erlenmeyer<br />
Des Erlenmeyers de 250 mL ont été remplis avec 45 g de substrat solide à une humidité<br />
de 50 %. Ces flacons ont été stérilisés par autoclavage pendant 30 min à 121°C. L’inoculation<br />
a été réalisée avec 5 g de spawn en mélangeant délicatement l’ensemble pour assurer une<br />
répartition homogène du spawn dans le substrat.<br />
3.5.3. Les cultures en colonnes de Raimbault<br />
Les cultures en FMS ont été réalisées en utilisant les colonnes décrites par Raimbault et<br />
Alazard (1980). Pour nos expériences, nous avons utilisé des colonnes de verre de deux<br />
dimensions différentes 20 et 40 cm de hauteur et de 2 cm de diamètre avec un volume utile de<br />
100 mL pour les substrats céréales et de 250 mL pour le substrat oléicole. Après l’autoclavage<br />
(121°C pendant 20 min) des colonnes préparées vides, elles ont été remplies avec le substrat<br />
solide inoculé. Les colonnes remplies ont été placées dans le dispositif FMS équipé d’un bain<br />
thermostaté à 25±2°C.<br />
39
3.5.4. La régulation des paramètres de FMS<br />
Matériel et Méthodes<br />
Le dispositif de FMS, mis au point par Raimbault (1980), permet le contrôle et le suivi<br />
de la fermentation de L. edodes sur le substrat oléicole (Fig. 12). L'aération dans les<br />
fermenteurs se fait grâce à une canalisation d'air comprimé à haute pression. A la sortie du<br />
manodétendeur, la pression de l'air a été maintenue à 0,2 bar. L'air a été ensuite humidifié<br />
deux fois à température ambiante à l'aide de bulleurs en verre frité et distribué en passant par<br />
un ballon à fond rond muni d'une entrée et de plusieurs sorties. L'aération de substrat dans les<br />
colonnes a été donc assurée par un flux d'air saturé en humidité et contrôlé par l'intermédiaire<br />
de vannes à pointeaux. Afin d’assurer la saturation et de la mise à température de l'air, un<br />
humidificateur individuel (petit bulleur) a été disposé sous chaque colonne FMS. Les<br />
effluents gazeux, collectés à la sortie de chaque colonne, ont été analysés à l’aide du<br />
respiromètre PNEO.<br />
Figure 12. Dispositif de FMS utilisé : 1, Entrée d’air ; 2, Bain marie thermostaté ; 3, Colonnes<br />
de Raimbault (Saucedo-Castañeda, 1995).<br />
3.5.5. La description et le fonctionnement du PNEO<br />
Un appareil prototype de respirométrie PNEO (Fig. 13) a été construit par l'équipe ITE<br />
du CNRS (Faculté des Sciences et Techniques de St Jérôme). Il a été mis au point au cours de<br />
cette étude pour suivre en ligne et de manière non destructive les principaux paramètres de<br />
FMS.<br />
40
Matériel et Méthodes<br />
Figure 13 . Photographie du respiromètre PNEO et description de ses différents<br />
composants, la partie supérieure étant composée des capteurs d'analyse de l'air sortant<br />
des colonnes et la partie inférieure étant la partie électronique avec la transmission des<br />
informations à l'ordinateur (ITE, 2009).<br />
PNEO est un appareil de respirométrie qui assure simultanément le suivi en ligne de<br />
quatre paramètres au cours de la FMS. Il permet la mesure de l'humidité relative de l'air<br />
passant à travers la colonne de fermentation entre 0 et 100%. Une mise à niveau du capteur<br />
est faite par passage d’un flux d'air sec entre chaque mesure pour éliminer la condensation qui<br />
pourrait se former sur le capteur.<br />
Il permet également la détermination de la température de l’air ,entre 0°C et 150°C, en<br />
sortie de colonne. Cependant la température mesurée est souvent sous-estimée par rapport à la<br />
température réelle, car l'air est refroidi dans le tuyau par l'air ambiant. Cela permet tout de<br />
même de savoir si la température reste stable au cours du temps.<br />
Le troisième paramètre qu’on peut mesurer avec le PNEO est le débit d'air en sortie de<br />
la colonne de fermentation. Le système permet l’ajustement du débit d’air. Le capteur peut<br />
mesurer des débits entre 0 et 200 mL.min -1 , ce qui laisse une marge puisque, le plus souvent,<br />
les débits utilisés sont compris entre 5 et 60 mL.min -1 . Le dernier paramètre mesuré est le CO2<br />
dégagé par le champignon. Le capteur infrarouge peut détecter des taux de CO2 compris entre<br />
41
Matériel et Méthodes<br />
0 et 10%. Ce paramètre permet de suivre en ligne la croissance du champignon.<br />
L’enregistrement des paramètres se fait grâce à un logiciel conçu par le même groupe qui<br />
permet l’exportation des données pour les calculs.<br />
3.6. L’échantillonnage et le traitement des échantillons en FMS<br />
A la différence des cultures liquides, la prise d’échantillons homogènes au cours des<br />
FMS est une chose difficile à obtenir. En effet, pour que les échantillons soient homogènes, il<br />
est nécessaire de sacrifier une colonne complète et de récupérer l’ensemble du produit<br />
fermenté et de l’homogénéiser avec un simple mélange manuel.<br />
Après chaque prélèvement, les produits fermentés ont été immédiatement homogénéisés<br />
et les échantillons frais ont été utilisés pour mesurer le poids sec et le pH. Les échantillons ont<br />
été congelés et lyophilisés dans un lyophilisateur (Heto Power Dry LL1500 Freeze Dryer,<br />
Thermo Electron Corporation, U.S.A.). Parallèlement, 10 g d’échantillons frais ont été<br />
congelés pour les analyses d’enzymes. Les échantillons lyophilisés ont subi un broyage dans<br />
un micro-broyeur (IKA Labortechnik) pour leur complète homogénéisation.<br />
3.7. Les techniques d’analyse<br />
3.7.1. La mesure de l’humidité<br />
En fermentation solide, il est indispensable de suivre l'évolution de la matière sèche du<br />
milieu de culture. En effet, l'eau produite lors de la dégradation des substrats carbonés, de<br />
même que la perte du poids sec due à la respiration (minéralisation des composés organiques)<br />
ainsi que le transfert de l'eau par le flux d'air saturé en eau conduisent à des variations<br />
importantes de l'humidité des produits au cours de la fermentation. Cette mesure de l'humidité<br />
permet d'exprimer la perte en poids après la fermentation. Le poids sec est évalué par<br />
différence de pesée.<br />
Environ 5 à 10 g d’échantillon de matière fraîche ont été pesés (P0) dans des coupelles<br />
en aluminium préalablement tarées (Pvide), à l’aide d’une balance de précision (Sartorius<br />
BP121S). L’échantillon a été mis à sécher à 105°C jusqu’à l’évaporation totale de l’eau dans<br />
le substrat (Psec). Les résultats sont exprimés en pourcentage de poids sec. La mesure a été<br />
effectuée en triplicata.<br />
Humidité (%) = [(P0 – Psec)/(P0 – Pvide)] x 100<br />
42
3.7.2. La mesure du pH<br />
Matériel et Méthodes<br />
Les mesures des valeurs du pH du substrat fermenté ont été réalisées dans une<br />
suspension (0,1 g.mL -1 d’eau distillée). La mesure a été réalisée à l'aide d'un pH-mètre digital<br />
Knick muni d'une électrode combinée. L'étalonnage de l'appareil a été obtenu avec des<br />
solutions tampon pH 4,0 - 7,0 et 10.0 (Schott-Geräte).<br />
3.7.3. La mesure de la croissance apicale<br />
La croissance apicale correspond à l’élongation du mycélium cultivé sur une surface<br />
solide. Cette élongation a été mesurée régulièrement à l’aide d’une règle graduée. La<br />
croissance apicale a été exprimée en millimètres rapportés au temps. Généralement, cette<br />
mesure a été réalisée pour des cultures en boîtes de Petri sur des milieux gélosés ou céréaliers.<br />
Vmoy = Vitesse de croissance apicale moyenne (mm.j -1 )<br />
Dmax = Diamètre de la colonie au moment de l’arrêt de la FMS (mm)<br />
C = Diamètre du disque de gélose (mm)<br />
T = Temps (h)<br />
Vmoy = [(Dmax- C)/2]/T<br />
3.7.4. L’estimation indirecte de la biomasse par analyse de l’ergostérol<br />
3.7.4.1. Extraction de l’ergostérol<br />
L’extraction de l’ergostérol a été faite à partir d’échantillons de produits fermentés: un<br />
échantillon de 50 mg d’échantillon lyophilisé a été mis en suspension dans 1 mL de méthanol<br />
pur (SIGMA) dans un tube Eppendorf de 1,5 mL en 5 exemplaires. Les échantillons ont été<br />
agités au vortex à 1500 rpm pendant 20 sec et laissés reposer pendant une nuit à 4°C à<br />
l’obscurité. Ces échantillons ont été ensuite agités au vortex à la même vitesse et centrifugés à<br />
14000 tr.min -1 dans une centrifugeuse SIGMA 3K15 à 4°C pendant 10 min. Les surnageants<br />
ont été récupérés pour les analyses de l’ergostérol en HPLC.<br />
3.7.4.2. Dosage de l’ergostérol en HPLC<br />
La plateforme HPLC-DAD Agilent technologies 1200 Series est composée d’un<br />
dégazeur (1200 Series Degasser, Agilent technologies), d’une pompe quaternaire (1200 Series<br />
Quaternary Pump, Agilent technologies), d’un échantillonneur automatique (1200 Series<br />
Automatic Liquid Sampler, Agilent technologies) et d’un détecteur à barrette de diodes (1200<br />
Series Diode-Array Detector, Agilent technologies). L’élution a été isocratique en utilisant le<br />
43
Matériel et Méthodes<br />
méthanol grade-HPLC comme phase mobile à un débit de 1 mL.min -1 . Un volume de 20-50<br />
µL de chaque échantillon a été injecté dans l’HPLC. Un lavage de la colonne a été effectué<br />
par l’injection du méthanol (qualité HPLC). Le pic de l’ergostérol a été déterminé par<br />
comparaison avec le temps de rétention du standard et en vérifiant le spectre d’absorption<br />
caractéristique de l’ergostérol. La DO pour l’ergostérol a été mesurée à 280 nm.<br />
3.7.5. Le dosage des sucres réducteurs<br />
Les sucres réducteurs ont été dosés par la méthode de Miller (1959). Les sucres<br />
réduisent l'acide 2,3-dinitrosalicylique (DNS) en acide 3-amino-5-nitrosalicylique, à chaud.<br />
Le produit de la réaction au milieu basique développe une coloration jaune orangé avec un pic<br />
d'absorption à 575 nm. L'échantillon à doser a été dilué convenablement de façon à obtenir<br />
une concentration en sucres réducteurs voisine de 1g.L -1 . A 1 mL de cette dilution, on a ajouté<br />
1 mL de réactif au DNS. Le mélange a été porté à ébullition pendant 5 min et refroidi<br />
rapidement. Après addition de 8 mL d'eau distillée, le contenu des tubes a été homogénéisé au<br />
vortex, et la densité optique a été lue à 575 nm. La courbe d’étalonnage a été réalisée avec une<br />
solution de glucose de 0 à 1g.L -1 .<br />
3.7.6. Le dosage des sucres totaux<br />
La réaction à l’anthrone constitue la base d’une méthode rapide pour la détermination<br />
des hexoses, acides hexouroniques et aldopentoses. Le complexe formé donne une couleur<br />
bleu-vert qui absorbe à 625 nm (Dubois et al.,1956). La présence de protéines contenant du<br />
tryptophane donne une couleur rouge à l’échantillon et peut donc interférer avec la réaction.<br />
Pour la préparation du réactif, 200 mg d’anthrone ont été dissous dans 100 mL d’acide<br />
sulfurique à 96 %. Ce réactif peut être conservé au frais pour 4 mois. Pour réaliser la courbe<br />
étalon, la solution mère a été préparée avec 5 g.L -1 de glucose (0,5 g.100 mL -1 ) et la courbe<br />
d’étalonnage a été établie pour des concentrations en glucose de 0 à 50 mg.L -1 .<br />
3.7.7. Le dosage des phénols totaux<br />
La détermination de la concentration des phénols totaux dans les différents échantillons<br />
a été réalisée par la technique de Folin Ciocalteu décrite par Bärlocher et Graça (2005). Le<br />
réactif de Folin Ciocalteu est réduit par le complexe phényl-cuivre qui donne une coloration<br />
bleue dont l’absorbance est maximale à 760 nm.<br />
La concentration des phénols totaux a été déterminée sur 0,1 mL d'échantillon<br />
(convenablement dilué dans de l'eau distillée) auquel sont rajoutés 5 mL du réactif C (2% de<br />
44
Matériel et Méthodes<br />
Na2CO3 dans NaOH, 0,1N). Après 5 min d’incubation à température ambiante, 0,5 mL de<br />
réactif de Folin Ciocalteu a été ajouté. Le milieu a été placé 2h à l'obscurité. L'absorbance a<br />
été lue à 760 nm contre un tube témoin. La gamme étalon a été réalisée dans les mêmes<br />
conditions en présence de l’acide cafféique (0 à 250 µg.mL -1 ).<br />
3.7.8. Le dosage des activités enzymatiques<br />
3.7.8.1. Activité Laccase<br />
L’extraction de laccase a été faite dans un tampon acétate (0,1M, pH 5,0) contenant<br />
5mM de CaCl2, 0,05% de tween et 5% de polyvinylpolypyrrolidone PVPP (Sampedro et al.,<br />
2007). Le mélange a été incubé pendant 1h sous agitation (100 rpm) à température ambiante.<br />
L’activité des laccases a été déterminée selon la méthode Criquet et al. (1999). Le<br />
milieu réactionnel a été constitué de 1mL d’extrait enzymatique et de 15 µL de syringaldazine<br />
(6 mg.mL -1 du méthanol). L’oxydation de syringaldazine en quinone a été déterminée en<br />
mesurant la densité optique à 525 nm (ε= 65 000 M -1 cm -1 ). L’activité des laccases a été<br />
exprimée en µmole de quinone formée par minute (U) et par gramme de matière sèche (U.g -1<br />
MS).<br />
3.7.8.2. Activité Mn peroxydase<br />
L’activité a été déterminée selon la méthode de Wariishi et al., (1992). Le milieu<br />
réactionnel a été composé de 1 mL de l’extrait enzymatique, 0.5 mM MnSO4 dans 50 mM du<br />
tampon malonate à pH 4,5, en présence de H2O2 (75 µM). La formation du complexe (Mn 3+ -<br />
Malonate) a été suivie à 270 nm (ε=11 590 M -1 cm -1 ).<br />
3.7.8.3. Activité Lignine peroxydase<br />
L’activité a été déterminée par le suivi de l’oxydation de l’alcool vératrylique à 310 nm<br />
(ε= 9 300 M -1 cm -1 ) en présence de H2O2. Le milieu réactionnel a été composé de 2,1 mL de<br />
tampon tartrate (0,1M, pH3), de 0,3 mL de H2O2 (50 µM), de 0,1 mL d’alcool vératrylique et<br />
0.5 mL d’extrait enzymatique.<br />
3.7.8.4. Activité tannase<br />
Nous avons utilisé la technique de Sharma et al., (2000) pour l’analyse des activités<br />
tannases produites par Pleurotus ostreatus cultivé sur coques de café en FMS. Cette technique<br />
utilise le méthylgallate comme substrat et sous l’action de la tannase, l’acide gallique libéré<br />
est colorée par la rhodanine. Cette nouvelle technique est très simple à réaliser et elle est trois<br />
fois plus sensible que les méthodes classiques. La gamme étalon a été réalisée avec de l’acide<br />
45
Matériel et Méthodes<br />
gallique (1 g.L -1 ). Une unité tannase correspond à la libération d’une µmole d’acide gallique<br />
par min à 35°C dans un tampon citrate 0,05 mM pH 5,0.<br />
3.7.8.5. Activité lipase<br />
Le milieu réactionnel est composé de 1 g du produit fermenté, 10 mM de laurate de p-<br />
nitrophényl (PNP), 2 mL d’heptane et de 2 mL d’eau. Il faut noter que la laurate de PNP est<br />
soluble dans l’heptane. Après l’incubation du mélange à l’obscurité à 30°C pendant 2 h avec<br />
agitation sur un secoueur, 4 mL d’eau ont été ajoutés et le tout a été filtré pour récupérer la<br />
fraction organique. Quatre mL de NaOH 0,1M ont été ajoutés à 200 µL du surnageant et la<br />
densité optique du mélange a été mesurée à 412 nm. La gamme d’étalonnage a été préparée<br />
avec différentes concentrations de PNP (Sigma, France).<br />
3.7.9. L’analyse des monomères phénoliques par HPLC<br />
L’extraction de la fraction phénolique a été effectuée avec une solution organique de<br />
méthanol : eau (60 :40). Un échantillon d’un gramme du substrat fermenté lyophilisé a été<br />
mélangé avec 10 mL de la solution organique, et le mélange a été homogénéisé et incubé<br />
pendant 1 h à 4°C. L’extraction a été répétée trois fois. Après la centrifugation du mélange,<br />
les surnageants ont été collectés et évaporés sous vide à 1/3 du volume. Une deuxième<br />
extraction a été faite par 10 mL l’acétate d’éthyle. Le surnageant a été évaporé sous vide à<br />
sec. Le résidu sec a été dissous dans 1 mL de méthanol pur (qualité HPLC) et injecté en<br />
HPLC.<br />
Les monomères phénoliques ont été déterminés par l’HPLC suivant la méthode<br />
modifiée de De Marco et al. (2007). Les conditions d’analyse sont décrites dans le Tableau 9.<br />
Tableau 9. Conditions d’analyses en HPLC pour la détection des monomères<br />
phénoliques.<br />
Débit 1 mL.min -1<br />
Volume d’injection 50 µL<br />
Température 25 ± 2 °C<br />
Solvant A Eau : Acide acétique, 97 :3 (v/v)<br />
Solvant B Acétonitrile : Méthanol, 80 : 20 (v/v)<br />
UV de détection 280 nm<br />
Spectre d’absorption 200-700 nm<br />
46
Matériel et Méthodes<br />
Tableau 10. Conditions d’élution pour la détection des monomères phénoliques.<br />
Condition Solvant A (%) Solvant B (%) Temps (min)<br />
3.7.10. Les analyses élémentaires : C et N<br />
Les teneurs en C et N, dans les échantillons de culture de L. edodes, ont été déterminées<br />
à l’aide d’un analyseur élémentaire (Flash EA 1212 Elemental Analyzer, France). Les<br />
échantillons ont été finement broyés, pesés et placés dans l’analyseur. L’analyseur a été<br />
calibré avec l’acide aspartique à différentes concentrations.<br />
3.7.11. La résonance magnétique nucléaire 13 C du solide<br />
Les spectres haute résolution ont été réalisés sur le spectromètre Bruker DSX 400 kHz<br />
du Spectropole (Université Paul Cézanne). Les spectres 13 C CP/MAS ont été enregistrés à la<br />
fréquence de 100,7 MHz avec découplage du proton. Environ 300 mg de substrat fermenté<br />
sont placés dans une sonde de 7 mm inclinée à l’angle magique (angle de 54,7° par rapport au<br />
champ magnétique statique) tournant à 6 kHz. Les paramètres d’acquisition utilisés pour un<br />
enregistrement optimal sont une impulsion de 90° pour le proton de 2,8 µs, un délai de<br />
répétition de 3 s, un nombre d’acquisition de 2000, et enfin un temps de contact de 2 ms. La<br />
déconvolution des spectres a été effectuée grâce au logiciel DmFit (Massiot et al., 2002). Le<br />
Tableau 11 et la Figure 14 présentent les principales régions obtenues en RMN du 13 C, et<br />
attribuées aux différents groupes fonctionnels trouvés dans la matière organique des litières et<br />
de compost.<br />
isocratique 80 20 10<br />
linéaire 60 40 15<br />
linéaire 45 65 20<br />
linéaire 25 75 30<br />
linéaire 0 100 45<br />
isocratique 0 100 50<br />
47
Matériel et Méthodes<br />
Tableau 11 . Attribution des régions RMN 13 C aux différents groupes fonctionnels trouvés<br />
dans la matière organique.<br />
Shift chimiques Désignation<br />
(ppm)<br />
Composés<br />
0-45 Alkyle C Lipides, acides aminés,<br />
45-60 N-Alkyle/méthoxyle C Acides aminés, (OCH3- lignine)<br />
60-110 O-Alkyle C Cellulose, hémicelluloses, autres polysaccharides<br />
110-145 Aryle C Polyphénols, Lignine (C aromatique)<br />
145-165 O-Aryle C Phénols, lignine (C phénolique)<br />
165-190 Carboxyle C acides organiques, amide et ester C<br />
Figure 14. Exemple de spectre CPMAS RMN 13 C d’une litière de Quercus ilex L.<br />
(Alarcon-Gutierrez, 2007).<br />
48
Résultats et discussion<br />
49
Résultats et discussion<br />
La partie « Résultats et Discussion » regroupe cinq chapitres préparés sous forme<br />
d’articles qui correspondent chacun à des étapes de mise au point technologique ou<br />
méthodologique afin de progresser vers la maîtrise de la valorisation des substrats oléicole par<br />
la culture de L. edodes. Chaque chapitre commence par une introduction en français qui<br />
justifie et relie entre les différents chapitres. Un résumé récapitule l’ensemble des résultats de<br />
chaque chapitre.<br />
Chapitre 1 :<br />
Criblage des souches de Lentinula edodes cultivées sur des margines d’olive en culture<br />
solide et liquide pour la biodégradation des polyphénols<br />
Lakhtar, H., Ismaili-Alaoui M., M., Philippoussis A., Perraud-Gaime I., Roussos S. (2009).<br />
Screening of strains of Lentinula edodes grown on model olive mill wastewater in solid and<br />
liquid state culture for polyphenol biodegradation. International Biodeterioration and<br />
Biodegradation (in press) (DOI: 10.1016/j.ibiod.2009.10.006).<br />
Chapitre 2:<br />
Optimisation de la production de l’inoculum (spawn) de Lentinula edodes sur les grains<br />
de blé (Ebly®).<br />
Lakhtar, H., Ismaili-Alaoui M., Roussos S., Non-esterified ergosterol analysis for Lentinula<br />
edodes spawn quantity estimation. A soumettre dans la revue Food Chemistry.<br />
Chapitre 3 :<br />
Culture de Lentinula edodes en fermentation en milieu solide sur substrat oleicole solide:<br />
Evaluation des facteurs de croissance.<br />
Lakhtar, H., Ismaili-Alaoui M., Labrousse Y., Roussos S., Solid state fermentation of<br />
Lentinula edodes on solid olive substrate: Evaluation of growth factors. A soumettre dans la<br />
revue Bioresource Technology.<br />
Chapitre 4 :<br />
Transformation de la matière organique et détoxification des substrats oléicoles solides<br />
par la culture de Lentinula edodes par fermentation en milieu solide.<br />
L’article de ce chapitre est en préparation.<br />
Chapitre 5 :<br />
Disponibilité des substrats oléicoles : séchage solaire sous serre du mélange de grignons<br />
d’olive et de margines<br />
Lakhtar, H., Macarie, H., Ismaili-Alaoui M., M., Perraud-Gaime I., Roussos, S., Solar drying<br />
in greenhouse of a mixture of olive cake and olive mill wastewater, a solution for olive mill<br />
wastewater disposal?. Accepté avec révision dans la revue Journal of Environmental<br />
Management.<br />
50
Résultats et discussion. Chapitre 1 : Criblage des souches de L. edodes<br />
Chapitre I<br />
Criblage des souches de Lentinula edodes cultivées sur des<br />
margines d’olive en culture solide et liquide pour la<br />
biodégradation des polyphénols<br />
Parmi les effluents les plus toxiques de l’industrie oléicole, on trouve les margines<br />
d’olive qui se caractérisent par un pH acide et une charge organique élevée associée à la<br />
présence i) des polyphénols à grande concentration et ii) des acides gras libres (Sayadi et al.,<br />
2000). En plus de la composition chimiques des margines, leurs volumes très importants<br />
produits durant la campagne oléicole causent d’énormes problèmes de gestion et<br />
d’environnement (Ben Sassi et al., 2006).<br />
La valorisation des margines par les champignons comestibles a attiré l’attention de<br />
plusieurs auteurs, par le fait que ces microorganismes sont capables de produire plusieurs<br />
enzymes ligninolytiques non spécifiques, telles que les laccases, les Mn peroxydases et les<br />
lignines peroxydases, impliquées dans la diminution de la toxicité des margines par la<br />
dégradation des polyphénols. Bien que les champignons comestibles (Pleurotus sp. et<br />
L. edodes) sont capables de se développer en présence des polyphénols, leur pouvoir à<br />
dégrader ces molécules diffère en fonction des souches de chaque genre (Sayadi et Ellouz,<br />
1993). Ainsi, une sélection de souches les plus performantes reste une étape importante dans<br />
le développement d’un procédé de valorisation des margines en vue d’éliminer leur toxicité.<br />
Dans ce premier chapitre sont présentés les résultats de l’étude de criblage des souches<br />
de L. edodes pour leur pouvoir de réduction des polyphénols de margines. Pour ce faire, une<br />
51
Résultats et discussion. Chapitre 1 : Criblage des souches de L. edodes<br />
collection de 16 souches de L. edodes provenant de différentes régions du monde a été établie<br />
et ces souches ont été évaluées pour leur tolérance aux margines. Le criblage des souches de<br />
la collection a été effectué en deux étapes : la première étape en culture en milieu solide<br />
gélosé et la deuxième étape en culture en milieu liquide en présence de margines. Plusieurs<br />
critères de sélection ont été choisis pour sélectionner une souche performante : tolérance aux<br />
margines, croissance apicale, biomasse mycélienne, décoloration des margines, dégradation<br />
des phénols totaux et production d’enzymes ligninolytiques.<br />
Ce chapitre a fait l’objet d’un article accepté pour publication dans la revue<br />
International Biodeterioration & Biodegradation (10.1016/j.ibiod.2009.10.006).<br />
52
Résultats et discussion. Chapitre 1 : Criblage des souches de L. edodes<br />
Screening of Lentinula edodes strains cultivated on model olive mill<br />
wastewater in solid and liquid state culture for polyphenols biodegradation<br />
Authors:<br />
Hicham Lakhtar 1 , Mustapha Ismaili-Alaoui 2 , A. Philippoussis 3 , Isabelle Perraud-<br />
Gaime 1 , Sévastianos Roussos 1*<br />
Author’s address:<br />
1 Institut Méditerranéen d'Ecologie et de Paléoécologie UMR CNRS/IRD 193; <strong>IMEP</strong> Case<br />
441; FST Saint Jérôme; Université Paul Cézanne; Av. Escadrille Normandie-Niemen; 13397<br />
Marseille cedex 20; France ;<br />
2 IAV –Hassan II, Laboratoire des Bioconversions ; BP. 6202-Instituts, 10101 Rabat, Maroc ;<br />
3 National Agricultural Research Foundation, IAMC, Laboratory of Edible and Medicinal<br />
Fungi, 61 Democratias St., 135 61 Ag. Anargyri, Athens, Greece<br />
*Corresponding author:<br />
Sévastianos Roussos: Institut Méditerranéen d'Ecologie et de Paléoécologie UMR CNRS/IRD<br />
193; <strong>IMEP</strong> Case 441; FST Saint Jérôme; Université Paul Cézanne; Av. Escadrille<br />
Normandie-Niemen; 13397 Marseille cedex 20; France ;<br />
E-mail: s.roussos@univ-cezanne.fr<br />
Tel: (+33) 04 91 28 91 03<br />
Fax: (+33) 04 91 28 86 68<br />
Scientific relevance of the study: Evaluation of shiitake cultivation (Lentinula<br />
edodes) as a biotechnological process (biodegradation and biotransformation) for<br />
simultaneous detoxification of phenol-rich olive mill wastewater (OMW) and the production<br />
of fungal enzymes and mycelial biomass. Application of an environmentally friendly fungal<br />
technology in the food (olive oil) chain production, medical research (L. edodes biomass) and<br />
bioremediation purposes (detoxification of OMW).<br />
53
Résultats et discussion. Chapitre 1 : Criblage des souches de L. edodes<br />
Criblage des souches de Lentinula edodes cultivées sur des margines d’olive<br />
en culture solide et liquide pour la biodégradation des polyphenols<br />
RESUME<br />
Au Maroc, l’industrie oléicole génère de grands volumes des margines d’olive dans une<br />
période courte (250 000 m 3 de margines pendant 4 mois, Novembre-Février). La présence des<br />
composés phénoliques confère une propriété phytotoxique ainsi qu’un effet antimicrobien.<br />
Plusieurs études ont été effectuées sur le traitement biologique et enzymatique des margines .<br />
Tandis que les perspectives d’utilisation des margines pour l’élaboration des produits à valeur<br />
ajoutée, telle que la culture de champignons comestibles, sont moins exploitées. Notre étude a<br />
pour objectif de sélectionner une souche de champignon comestible et médicinal capable de<br />
se développer en présence de margines produisant une grande quantité de biomasse<br />
mycélienne. La décoloration et la dégradation des phénols totaux des margines ont été<br />
également testées. Seize souches de Lentinula edodes ont été initialement évaluées pour leurs<br />
tolérance, croissance apicale et production de biomasse, sur milieux gélosés contenant les<br />
margines à différentes concentrations. Parmi les souches testées, la meilleur production de<br />
biomasse a été observée pour les souches (Le118, Le119, Le121 and Le122) et même en<br />
présence de 20% (v/v) de margines. Pour la seconde évaluation, la capacité des souches<br />
présélectionnées, pour décolorer et dégrader les phénols totaux de margines, a été testée en<br />
milieu liquide contenant des margines sans addition de nutriments. La souche de L. edodes<br />
Le119 a montré une décoloration des margines de 65% associée à l’élimination de 75% des<br />
phénols totaux mesurés par la méthode de Folin Ciocalteu. Cette souche a été selectionnée<br />
pour la culture en FMS de L. edodes sur les co-produits oléicoles. La reussite du procédé de<br />
FMS necessite la preparation d’un inoculum de très bonne qualité.<br />
Mots clé : Lentinula edodes, Margines d’olive, Culture en milieu solide, Culture en milieu<br />
liquide, Biomasse, Phénols totaux, Laccase, Détoxification.<br />
54
ABSTRACT<br />
Résultats et discussion. Chapitre 1 : Criblage des souches de L. edodes<br />
In Morocco, olive industry produces great amounts of Olive Mill Wastewater (OMW)<br />
in short time period (250 000 m 3 of liquid wastes in four months, November-February).<br />
Phenolic compounds are largely responsible for the phytotoxicity and antimicrobial effects of<br />
OMW. Several studies have been carried out focusing on the biological and enzymatic<br />
treatment of OMW. However, the use of OMW to produce value-added products, e.g.<br />
mushroom cultivation, are less explored. This research aimed to select shiitake mushroom,<br />
capable of growing on OMW, involving decolorization, removal of total phenol and high<br />
production of mycelial biomass. Sixteen strains of Lentinula edodes were evaluated for their<br />
tolerance to OMW, apical growth rate and biomass production on agar media. The highest<br />
biomass yields were recorded in four L. edodes strains (Le118, Le119, Le121 and Le122)<br />
grown in the presence of 20% (v/v) OMW. The ability of these pre-selected strains to<br />
decolorize and to remove total phenol from OMW was then tested by using the technique of<br />
liquid culturing, without any other nutriment supplement. L. edodes strain Le119 exhibited<br />
65% decolorization along with 75% elimination of total phenol as resulted from Folin<br />
Ciocalteu assays. The laccase activity was the main lignolytic activity observed, with one<br />
isoform of the enzyme got stained on native PAGE with p-phenylenediamine as the substrate<br />
at pH 5.0.<br />
Keywords: Lentinula edodes, Olive Mill Wastewater, Solid State Culture, Liquid State<br />
Culture, Biomass, Total Phenol, Laccase, Detoxification.<br />
55
1. Introduction<br />
Résultats et discussion. Chapitre 1 : Criblage des souches de L. edodes<br />
The production of olive oil is expanding in the Mediterranean region, which accounts<br />
for 95% of total worldwide production (Ismaili-Alaoui and Heddoun, 2006). Main producers<br />
of olive oil are Spain, Italy, Greece, Tunisia and Morocco (International Olive Oil Council,<br />
2008). The extraction of oil from olives generates large quantities of liquid by-products,<br />
which have been estimated in excess of 30 x 106 m3 per year (Crognale et al., 2006).<br />
Chemical characteristics of these liquid wastes depend on olive varieties, as well as the olive<br />
oil extraction system.<br />
In Morocco, about 50% of olive oil production comes from traditional mills called<br />
“Maâsra”, which use the conventional “three-phase system” (Ismaili-Alaoui and Heddoun,<br />
2006). A major drawback of this system is that huge amounts of olive mill wastewater<br />
(OMW) are discarded, containing a high concentration of polyphenols (Ben Sassi et al.,<br />
2006). OMW are now generating pollution problems in the Mediterranean region, particularly<br />
in Morocco, where suitable treatments for these effluents are poorly developed (Ben Sassi et<br />
al., 2006).<br />
Detrimental effects of OMW (Sayadi et al., 2000) on plant growth (Isidori et al.,<br />
2005), microbial activity, and soil properties have already been described (Saadi et al., 2007).<br />
The presence of phenolic compounds in high concentrations is considered the main factor<br />
from OMW causing harmful effects (Sayadi et al., 2000; El Hajjouji et al., 2007). Extensive<br />
research on white-rot basidiomycetous fungi has been conducted in order to select strains<br />
showing high detoxification capability (i.e., simultaneous removal of phenolic compounds<br />
and color) on OMW (Sayadi and Ellouz, 1993; D’Annibale et al., 2004). Other research<br />
efforts are focused on the use of fungal enzymes involved in the degradation of polyphénols<br />
(D’Annibale et al., 2000; Tsioulpas et al., 2002). The cultivation of edible mushrooms<br />
(Pleurotus or Lentinula) using OMW as a wetting agent has also been carried out (Zervakis et<br />
al., 1996; Kalmis and Sargin, 2004; Kalmis et al., 2008).<br />
Lentinual edodes (shiitake) is a white-rot mushroom 1 cultivated commercially, which<br />
has medicinal properties, as well as biodegradation and biotransformation abilities<br />
(Philippoussis et al., 2007; Israilides et al., 2008; Philippoussis, 2009). This species produces<br />
extracellular oxidizing enzymes capable of biodegrading lignin-related recalcitrant<br />
compounds, such as polyphenols (D’Annibale et al., 1998). The efficiency of polyphenol<br />
degradation by L. edodes is associated with several enzymes, including lignin peroxidase,<br />
56
Résultats et discussion. Chapitre 1 : Criblage des souches de L. edodes<br />
manganese peroxidase and laccase (D’Annibale et al., 2004; Ayed et al., 2005; Matos et al.,<br />
2007). However, the production of these enzymes depends on substrate composition,<br />
cultivation system and environmental factors affecting fungal growth (Alaoui et al., 2008;<br />
Elisashvili et al., 2008).<br />
In previous research work, it has been shown that mycelial growth rate, biomass yield<br />
and enzymatic activities are important factors for successful mushroom cultivation<br />
(Philippoussis et al., 2003; Silva et al., 2005). In this study, we selected strains of L. edodes<br />
capable not only of tolerating the phenolics present in OMW, but also of using them as a<br />
carbon source to yield high mycelial biomass. Decolorization and removal of phenolic<br />
compounds were also criteria of selection.<br />
2. Materials and methods<br />
2.1. Substrates<br />
OMW was obtained from an olive oil plant at Beni-Mellal (central Morocco), after the<br />
three-phase extraction. OMW was stored at -20°C until used. The composition of untreated<br />
OMW is such as: pH (4.9), suspended solids (13.84 %), total sugar (12,8 g l-1), reducing<br />
sugar (10.08 g l-1) and total phenol (12.54 g l-1).<br />
2.2. Microorganisms<br />
Sixteen strains of Lentinula edodes (Berk.) Pegler were obtained from different<br />
laboratories (Table 1). These strains were already selected for their ability to colonize<br />
agricultural residues. All strains were routinely grown on potato dextrose agar (PDA, Sigma,<br />
St Quentin Fallavier, France) plates. Inoculum was prepared by growing the mycelium on<br />
agar plates for 2 weeks.<br />
57
Résultats et discussion. Chapitre 1 : Criblage des souches de L. edodes<br />
Table 1: Origin and source of different strains of Lentinula edodes<br />
Strains Origin Source<br />
Le118 AMRL* China<br />
Le119 AMRL China<br />
Le120 AMRL China<br />
Le121 AMRL Europe<br />
Le122 AMRL Europe<br />
Le123 AMRL Europe<br />
Le124 AMRL Taïwan<br />
Le125 Commercial strain Osaka Japon<br />
Le126 Commercial strain Osaka Japon<br />
Le127 Commercial strain France Mycelium<br />
Le128 Commercial strain France Le Champion<br />
Le129 Commercial strain Belgique Mycelia<br />
Le130 Commercial strain USA, Lamber spawn<br />
Le131 Commercial strain Japon<br />
Le132 Hybrid strain INRA France<br />
Le133 Homocaryon of commercial strain Le Lion France<br />
* Athens Mushroom Research Laboratory (NAGREF)<br />
2.3. Solid State Culture<br />
Tolerance of shiitake mycelium was investigated on PDA containing different<br />
concentrations of OMW, ranging from 0-100%. The agar medium was sterilized at 121°C for<br />
20 min, cooled, and used to prepare PDA plates. These plates were inoculated with agar disks<br />
(10 mm diameter) from an actively growing colony, and then incubated at 25°C. Linear<br />
growth rates were determined as the average distance covered by the mycelium in two<br />
perpendicular directions. After complete colonization of Petri plates (90 mm diameter), the<br />
mycelial biomass was assessed using a sterile cellophane disc placed on the agar surface, as<br />
described by De Araujo et al. (2000). The mycelium was allowed to grow on the cellophane<br />
disc, and then lifted away to be dried at 105°C.<br />
58
2.4. Liquid State Culture<br />
Résultats et discussion. Chapitre 1 : Criblage des souches de L. edodes<br />
The inoculum of strains was prepared by growing the mushroom mycelium on PDA<br />
containing 2% (v/v) OMW. Erlenmeyer flasks containing 100 mL of OMW at 10% (v/v) were<br />
sterilized (121°C for 20 min), inoculated with three agar plugs (10 mm diameter), and<br />
incubated at 25±2°C for 30 days, under shaking (120 rpm) conditions. Controls were prepared<br />
by OMW without inoculum, and incubated under the same conditions. Experiments were<br />
performed in triplicate<br />
2.5. Analytical assays<br />
Total phenols were quantified by the method of Folin and Ciocalteu (Bärlocher and<br />
Graça, 2005). The blue reactive complex formed was determined spectrophotometrically at<br />
760 nm. The phenolic content of the samples were expressed as caffeic acid equivalent. After<br />
filtration of OMW from the liquid culture, decolorization of OMW was measured at 395 nm<br />
as reported by Sayadi and Ellouz (1993).<br />
2.6. Molecular weight distribution of polyphenols<br />
Gel filtration chromatography using Sephadex G-50 was carried out for the analysis of<br />
polyphenolic compounds present in raw and treated OMW (Sayadi and Ellouz, 1993). The<br />
column was previously equilibrated with NaOH 0.05 M, LiCl 0.025 M. The flow rate was<br />
adjusted to 0.3 mL.min -1 . The column was calibrated with polymeric standards (Blue Dextran,<br />
2 000 kDa; Carbonic anhydrase from Bovine, 29 kDa; Cytochrome C from Horse Heart,<br />
12.4 kDa; Aprotinin: 6.5 kDa and Syringic acid: 0.195 kDa).<br />
2.7. Chromatographic analyses<br />
The monomeric composition of the phenolic fraction in OMW was extracted with<br />
ethyl acetate as described by De Marco et al., (2007), and it was analysed by reversed phase<br />
HPLC using binary gradient elution. The analysis was performed on an Agilent HPLC system<br />
(USA) equipped with diode array detector. The chromatographic separation was achieved on<br />
5 µm dC18 (4.6x250 mm) reversed-phase column (Atlantis, Ireland). The solvent system used<br />
was a gradient of solvent A (water : acetic acid, 97:3 v/v) and solvent B (acetonitrile :<br />
methanol, 80:20 v/v). The gradient used was previously described by De Marco et al., (2007).<br />
59
Résultats et discussion. Chapitre 1 : Criblage des souches de L. edodes<br />
Peak detection was carried out at 280 nm. The monomeric phenols were identified by<br />
comparing the retention time of eluted peaks to that of standards.<br />
2.8. Enzyme assays<br />
Laccase (E.C. 1.10.3.2: benzenediol: oxygen oxidoreductase) activity was assayed<br />
spectrophotometrically at 30°C using syringaldazine as substrate and monitoring the<br />
formation of quinone at 525 nm (ɛ=65.000 M -1 cm -1 ) (Criquet et al., 1999). Manganese<br />
peroxidase (MnP) and Lignin peroxidase (LiP) activities were determined by the oxidation of<br />
veratrylic alcohol at 310 nm (ɛ= 93 000 M -1 cm -1 ) to veratrylic aldehyde. One international<br />
unit of enzyme activity (IU) is defined as the amount of enzyme that produces 1 µmol of<br />
product per min. Interferences in enzymatic assays due to OMW were eliminated by adding<br />
insoluble polyvinylpolypyrrolidone (PVPP), as described by Sampedro et al. (2007).<br />
2.9. Statistical treatment<br />
One way analysis of variance (ANOVA) was used to analysze the biomass production<br />
data and multiple pair-wise comparisons were performed by the Tukey test using Xlstat®<br />
software.<br />
3. Results<br />
Strain selection involved two steps. In solid state culture, apical growth and biomass<br />
yield were determined for all strains in order to select the best ones for OMW bioremediation.<br />
The second step was carried out in liquid state culture, in the presence of OMW. Kinetics of<br />
decolorization and the removal of total phenol from OMW were assessed during 30 days with<br />
respect to the ligninolytic system. The change of polymeric and monomeric phenol<br />
compounds was determined.<br />
60
Résultats et discussion. Chapitre 1 : Criblage des souches de L. edodes<br />
3.1. Tolerance of OMW by Shiitake strains<br />
All strains of L. edodes were capable of growing on PDA with 10% and 20% of OMW<br />
(v/v), while no mycelial growth was observed above 40% of OMW. The period of initial<br />
growth varied among strains. By contrast, the period for complete colonization of the Petri<br />
plate by L. edodes strains varied from 10-30 days. Apical growth rates were significantly<br />
different among strains. They were reduced by 50% as the concentration of OMW increased<br />
in PDA (Fig. 1).<br />
Apical growth ( mm day -1 )<br />
12,00<br />
10,00<br />
8,00<br />
6,00<br />
4,00<br />
2,00<br />
0,00<br />
118 119 120 121 122 123 124 125 126 127 128 129 130 131 132 133<br />
L. edodes strains<br />
Figure 1: Growth rate (mm day -1 ) of sixteen L. edodes strains on PDA with 10% and 20% of<br />
OMW in solid culture at 25°C until complete colonization of the Petri plate (diam. 90mm).<br />
The apical growth of all strains was higher on PDA without OMW (data not shown).<br />
The highest growth rate was recorded for Le118 growing on agar medium containing 10%<br />
and 20% of OMW (10.72 and 4.97 mm day -1 , respectively), while the lowest growth rate was<br />
shown by the strain Le126 (1.38 and 0.46 mm day -1 , respectively).<br />
61
3.2. Biomass production<br />
Résultats et discussion. Chapitre 1 : Criblage des souches de L. edodes<br />
Dried mycelial biomass produced on PDA containing OMW was different among the<br />
strains of L. edodes (Table 2). The strain Le119 showed the highest biomass production (7.02<br />
mg day -1 l -1 ), followed by Le121, Le122, and Le118 (5.43 mg day -1 l -1 , 5.26 mg day -1 l -1 , and<br />
4.75 mg day -1 l -1 , respectively). The lowest biomass yield was recorded in the strain Le 133<br />
(2.29 mg day -1 l -1 ). In the case of strain Le119, the addition of 10% of OMW to PDA resulted<br />
in a threefold increase of biomass production. However, the increase of OMW concentration<br />
to 20% reduced the biomass yield of all strains. On the basis of mycelial growth<br />
characteristics (apical growth rate and biomass production) on PDA containing with 10% and<br />
20% of OMW, four strains (Le118, Le119, Le121, Le122) were selected for further<br />
evaluation.<br />
Table 2: Mycelial biomass production (mg day -1 l -1 ) during solid culture of sixteen L.<br />
edodes strains on PDA with 10% and 20% of OMW in 25°C.<br />
L. edodes PDA with 10%<br />
PDA with 20%<br />
strains of OMW<br />
of OMW<br />
Le119 7,02 a 1 4,49 a<br />
Le121 5,43 b 4,40 a<br />
Le122 5,26 b,c 3,96 b<br />
Le118 4,75 c,d 3,99 b<br />
Le124 4,33 d,e 0,36 g<br />
Le127 4,30 d,e 3,47 c<br />
Le129 4,23 d,e 2,49 d<br />
Le125 4,18 d,e 2,43 d<br />
Le123 4,15 d,e 0,44 g<br />
Le128 4,07 e 3,96 b<br />
Le131 4,05 e 0,35 g<br />
Le130 4,02 e 3,54 c<br />
Le126 3,28 f 3,67 b<br />
Le132 3,13 f 1,42 f<br />
Le120 2,90 f 0,00 h<br />
Le133 2,29 g 1,85 e<br />
1<br />
Mean values of each parameter within the same column not sharing common letters<br />
are significantly different (p
3.3. Decolorization of OMW<br />
Résultats et discussion. Chapitre 1 : Criblage des souches de L. edodes<br />
The decolorization of OMW by four selected strains of L. edodes is shown in Figure 2.<br />
The maximum amount of color removal was recorded in the strain Le119 (65%), while the<br />
strain Le118 exhibited low OMW decolorization during the initial stage of incubation but<br />
increased absorbance values with time. The other two strains were found to be moderate<br />
decolorizers of OMW. At the end of incubation period, a progressive darkening of the<br />
biomass was observed, which could be due to the adsorption of phenolic polymers by the<br />
mycelium. Accumulation of colored compounds into the mycelium has been estimated in<br />
about 10 % of the initial total color.<br />
Absorbance at 395 nm<br />
2,5<br />
2<br />
1,5<br />
1<br />
0,5<br />
0<br />
0 5 10 15 20 25 30<br />
Time (day)<br />
Figure 2: Profiles of color in OMW incubated, without inoculation (control) ( ) and with<br />
four L. edodes strains Le118 ( ), Le119 ( ), Le121 ( ) and Le122 ( ), at 25°C.<br />
63
Résultats et discussion. Chapitre 1 : Criblage des souches de L. edodes<br />
3.4. Degradation of total phenol<br />
The kinetics of total phenol degradation by L. edodes on OMW is shown in Figure 3.<br />
Degradation appeared to be correlated with decolorization (r 2 = 0.867, P
Résultats et discussion. Chapitre 1 : Criblage des souches de L. edodes<br />
3.5. Laccase enzyme production<br />
Laccase production was the main enzymatic activity observed during experiments,<br />
which involved degradation of the phenolic fraction. Figure 4 shows significant difference in<br />
laccase activity among the four strains studied of L. edodes.<br />
Laccase activity (UI ml -1 )<br />
2<br />
1,6<br />
1,2<br />
0,8<br />
0,4<br />
0<br />
0 5 10 15 20 25 30<br />
Figure 4: Time courses of laccase activity of four L. edodes strains Le118 ( ), Le119 ( ),<br />
Le121 ( ) and Le122 ( ) grown on OMW liquid culture without supplement, incubated at<br />
25°C.<br />
The onset of laccase occurred after 5 days of incubation, while the maximum enzyme<br />
activity was reached by the strain Le119 after 15 days (1.6 ± 0.21 UI ml -1 ). The strain Le118<br />
produced low and steady titer of laccase (less than 0.2 UI ml -1 ). The strain Le122 exhibited<br />
the maximum enzyme activity (0.65 UI ml -1 ) after 15 days, while the strain Le121 towards the<br />
end of the incubation period (20-30 days).<br />
Time (day)<br />
65
Résultats et discussion. Chapitre 1 : Criblage des souches de L. edodes<br />
3.6. Molecular weight distribution of OMW<br />
The evolution of molecular weight distribution from extractable polymeric fraction of<br />
OMW during the incubation of selected strains of L. edodes is shown in Figure 5.<br />
OD 280nm<br />
Fractions (Volume)<br />
Figure 5: Evolution of the molecular weight distribution of aromatic polymer present in<br />
OMW without inoculation ( ) (control) and after 30 days of incubation at 25°C with four L.<br />
edodes strains ( ) Le118, ( ) Le119, ( ) Le121 and ( ) Le122. A: Carbonic anhydrase<br />
from Bovine, 29kDa; B: Cytochrome C from Horse Heart, 12.4 kDa; C: aprotinin, 6.5 kDa).<br />
The untreated OMW showed three groups of aromatic compounds. The first peak had<br />
a molecular weight (MW) higher than 30 kDa, the second one less than 12.5 kDa, and the<br />
third one apparently less than 5.5 kDa. This clearly indicated that, after 30 days of incubation<br />
of strains, the MW distribution of polyphenol was affected. The reduction of the first fraction<br />
(MW >30 kDa) was recorded in the strains Le119, Le121 and Le122, while the same fraction<br />
was highly produced by the strain Le118.The strains Le118, Le119, and Le121 produced<br />
lower depolymerization of high molecular weight aromatics than the strain Le122.<br />
66
Résultats et discussion. Chapitre 1 : Criblage des souches de L. edodes<br />
3.7. Phenolic monomer compound<br />
Ethyl acetate OMW extracts were analyzed with reversed-phase HPLC in order to<br />
assess degradation of individual monomeric aromatic components (Table 3).<br />
Chromatographic analyses showed that the strain Le119 was capable of completely removing<br />
hydroxytyrol, caffeic acid, and para-coumaric acid. The removal of other phenolic<br />
compounds was of 67.12% and 89.81% for vannilic acid and tyrosol, respectively. The other<br />
three strains (Le118, Le121, Le122) completely removed at least one compound, and partially<br />
removed 22% of oleuropein (strain Le122) and 93.25% of para-coumaric acid (Le118). Non-<br />
phenolic compound, such as trans-cinnamic acid, was not degraded.<br />
Table 3: Monomeric aromatic compound removal (%) with four L. edodes strains (Le118,<br />
Le119, Le121 and Le122), in liquid culture of OMW at 25°C after 30 days of incubation.<br />
OMW component Le118 Le119 Le121 Le122<br />
3,4-Dihydroxyphenylethanol<br />
(hydroxytyrosol)<br />
67 100 57 73.51<br />
4-Hydroxyphenylethanol (tyrosol) 88.83 89.81 79.04 77<br />
(+)-Catechin 73.35 68.98 100 65<br />
Vannilic acid 0 67.12 20.29 11.97<br />
Caffeic acid 100 100 100 100<br />
Para-Coumaric acid 93.25 100 53.07 100<br />
Trans-Cinnamic acid 0 0 0 0<br />
Oleuropein 74.07 77.85 84.14 22<br />
4. Discussion<br />
The use of cellophane membrane allowed the growth of all strains studied of<br />
L. edodes, as well as the color change of OMW agar medium after incubation (PDA<br />
containing 10% and 20% OMW). The initial color was black turning to clear brown after<br />
incubation, excepting the strain Le118 which exhibited dark color at the end of colonization.<br />
This change of color is an indicator of the action of fungal enzymes on the hydrolysis of<br />
phenols during mycelial growth (Sobal et al., 2003). The presence of OMW in the culture<br />
medium affected mycelial growth of all strains. Three selected strains (Le119, Le121, Le122)<br />
showed mycelial colonies of high density, while the strain Le118 had low density. The use of<br />
PDA allowed better assessment of decolorization efficiency in all strains. In fact, it has been<br />
67
Résultats et discussion. Chapitre 1 : Criblage des souches de L. edodes<br />
shown that the metabolism for decolorization requires the presence of readily available carbon<br />
source, such as glucose (D’Annibale et al., 1998). Enzymatic determination using ABTS<br />
(2,2'-azino-bis-(3-ethylbenzothiazoline-6-sulphonic acid) test (Collins et al., 1998), showed<br />
that most selected strains, except Le118, were capable of producing laccase on PDA without<br />
OMW (data not shown).<br />
Comparative assessment, based on apical growth rate and biomass yield on OMW<br />
agar medium, showed that the strains Le118, Le119, Le121 and Le122 were the most<br />
efficient, and accordingly selected for further evaluation of decolorization and the removal of<br />
total phenol from OMW. The inhibitory effects of OMW on fungi have already been reported<br />
(Fountoulakis et al., 2002; Alaoui et al., 2008). Considering this fact, dilutions of OMW were<br />
prepared to allow mycelial growth of L. edodes. In this study, the use of mycelium previously<br />
grown on PDA containing 2% of OMW resulted in enhanced tolerance of strains to inhibitory<br />
compounds from liquid OMW. This positive effect of pre-adaptation to OMW, previously<br />
reported for several white-rot fungi (Leontievsky et al., 2002; D’Annibale et al., 2004), has<br />
been attributed to the triggering effect of phenolic compounds for inducing the production of<br />
ligninolytic enzymes (Fenice et al., 2003 ; Farnet et al., 2004).<br />
Laccase was detected as the main lignin-degrading enzyme in strains studied, which<br />
was similar to previous reports (D’Annibale et al., 2004; Olivieri et al., 2006). It has also been<br />
reported that LiP and MnP are capable of decolorizing and removing phenolics from OMW<br />
(Sayadi and Ellouz 1993, 1995). The molecular weight of phenolic compound from OMW<br />
was modified by all strains at different levels. The strains Le119 and Le121 were able to<br />
reduce all fractions obtained from OMW extract. This was in agreement with previous results<br />
from Jouani et al. (2003) and D’Annibale et al. (2004). However, Casa et al. (2003) reported<br />
the appearance of a novel phenolic fraction in OMW after treatment with laccase.<br />
The difference of distribution patterns from aromatic compounds was also observed in<br />
OMW incubated with the fungus P. chrysosporium in liquid culture, which was associated to<br />
the production of LiP, and with purified LiP (Sayadi and Ellouz, 1993, 1995). This<br />
interpretation is based on the assumption that other auxiliary degradative mechanisms<br />
involving enzymes and small molecular weight agents, such as hydrogen peroxide and oxalic<br />
acid (D’Annibale et al., 1998), are implicated in the breakdown of aromatic substances, which<br />
could serve as substrates brought about by white-rot fungi (Durán and Esposito, 2000). The<br />
68
Résultats et discussion. Chapitre 1 : Criblage des souches de L. edodes<br />
fact that low molecular weight aromatics remained could be explained by the limitation of<br />
incubation time or the enzymatic system (Sayadi and Ellouz 1993).<br />
In relation to the effect of mycelial culture on OMW phenolic compounds, the strain<br />
Le119 was found to be more efficient than the other strains for completely removing<br />
hydroxytyrosol, caffeic acid and para-coumaric acid. The monomeric phenol compounds<br />
have been shown to be particularly phytotoxic, and this toxicity increases with the synergistic<br />
effect of monomers (Boukhoubza et al., 2007). Therefore, Isidori et al. (2005) showed that the<br />
most toxic compounds were catechol (EC50s ranging from 0.40 mmol l -1 for Sorghum bicolor<br />
to 1.09 mmol l -1 for Cucumis sativus) and hydroxytyrosol (EC50s ranging from 0.47 mmol l -1<br />
for S. bicolor to 1.55 mmol l -1 for C. sativus). The non degradation of trans-cinnamic could be<br />
due to the lack of appropriate substituent in the aromatic ring to enable laccase action<br />
(D’Annibale et al., 2004).<br />
Considering all screening parameters studied (Table 4), the strain Le119 of Lentinula<br />
edodes showed the highest performance for treatment of OMW. Accordingly, it has been<br />
selected for further experimental work in solid state cultures, using a mixture of olive<br />
biomass, including olive tree wood and olive cake, as substrate, where OMW acts as the<br />
wetting/moistening agent.<br />
Table 4: Comparison of performance of four L. edodes strains (Le118, Le119, Le121 and<br />
Le122) tested in liquid culture of OMW at 25°C.<br />
L. edodes<br />
strains<br />
Decolorization (%)<br />
30 th day<br />
Total phenols<br />
removal (%)<br />
30 th day<br />
Laccase activity<br />
15 th day (UI.ml -1 )<br />
Monomeric<br />
phenols<br />
removal (%)*<br />
Le119 65 75 1.50 75<br />
Le121 50 59 0.69 61<br />
Le122 42 38 0.65 56<br />
Le118 - 21 0.12 62<br />
* average of degradation of the eight monomeric compounds tested.<br />
Acknowledgements<br />
Hicham Lakhtar is thankful to DSF-IRD France for awarding the PhD fellowship. The<br />
authors wish to thank Dr. J. M. Savoie who kindly provided some of the L. edodes strains.<br />
69
Résultats et discussion. Chapitre 2: Optimisation de la production du spawn<br />
Chapitre II<br />
Optimisation de la production de l’inoculum (spawn) de<br />
Lentinula edodes sur les grains de blé (Ebly®)<br />
Dans le chapitre précédent, nous avons montré que la souche L. edodes Le119 était la<br />
plus performante parmi celles de la collection; nous l’avons retenue pour la suite du travail.<br />
Ce chapitre a pour but d’étudier la production de l’inoculum (spawn) de la souche Le119 sur<br />
les grains de céréales. Nous avons étudié la physiologie de croissance de la souche Le119 sur<br />
les grains de blé de marque Ebly ® pour la production d’inoculum en grande quantité et<br />
acclimaté pour la culture sur margines. Car pour initier la culture de L. edodes en FMS et<br />
éviter la contamination du milieu par d’autres microorganismes antagonistes, il faut disposer<br />
d’un inoculum performant (Mamiro et Royse, 2008). En effet, lors des premiers essais de<br />
culture de la souche Le119 sur des mélanges de substrats oléicoles (bois de la taille, grignons<br />
d’olive et margines), nous avons rencontré des problèmes de contamination des cultures par<br />
des moisissures antagonistes (Aspergillus niger, Trichoderma harzianum). Pour surmonter ce<br />
problème, nous avons modifié la technique de préparation du spawn, par l’ajout des margines<br />
à 10% (v/v) lors de la préparation de l’Ebly ® .<br />
Cette technique de culture du mycélium sur Ebly ® imbibé de margines a permis<br />
d’obtenir d’une part l’induction de la production de laccases par le mycélium et d’autre part la<br />
production importante de mycélium sur les grains de l’Ebly ® . Dans un premier temps, nous<br />
présentons l’induction du mycélium développé sur l’Ebly ® pour la production de laccases et,<br />
dans un deuxième temps, l’estimation de la biomasse mycélienne sur les grains de l’Ebly ® par<br />
la quantification de l’ergosterol sera présentée sous forme d’article.<br />
70
Résultats et discussion. Chapitre 2: Optimisation de la production du spawn<br />
I. Induction de l’activité laccase<br />
Dans la figure 15 sont présentés les résultats comparés de la production des activités<br />
laccases produites par L. edodes Le119 cultivé sur deux milieux de culture en présence ou en<br />
absence de margines.<br />
Activité laccase (UI.g -1 SPS)<br />
Figure 15 : Activité laccase produite par L. edodes Le119 sur des grains d’Ebly sans margines<br />
( ) et avec des margines à 10% (v/v) ( ), incubés à 25°C avec aération forcée de 25 mL.min -1<br />
Temps (jour)<br />
pendant 20 jours.<br />
On observe que, dans le milieu Ebly ® sans margines, l’activité laccase produite reste<br />
très faible, alors que sur le milieu d’Ebly avec des margines à 10% l’activité laccase produite<br />
est importante (1,7 UI.g -1 SPS après 12 jours d’incubation à 25°C). La présence de margines<br />
dans le milieu de culture induit fortement la production de laccases par L. edodes Le119. Ces<br />
résultats sont en accord avec les résultats rapportés par plusieurs auteurs (Munoz et al., 1997;<br />
Shicheng et al., 2003; Farnet et al., 2004), qui montrent que l’addition au milieu de culture<br />
des molécules phénoliques, telles que l’acide vanillique et l’acide férulique, induit une<br />
importance augmentation de l’activité laccase par les basidiomycètes. D’autres composés,<br />
telles que les atomes de cuivre et de cadmium ont été également utilisés comme inducteurs<br />
pour l’augmentation de la production des laccases (Velazquez Cedeño, 2005).<br />
Par ailleurs, plusieurs études ont montré que les laccases sont impliquées dans le<br />
mécanisme de défense de L.edodes face aux antagonistes tels que les moisissures de genre<br />
Trichoderma sp. en culture sur paille de blé (Mata et Savoie, 1998 ; Savoie et Mata, 1999,<br />
Savoie et Mata, 2003). En effet, les souches de L. edodes forment des barrières mélanisées<br />
71
Résultats et discussion. Chapitre 2: Optimisation de la production du spawn<br />
liées à une forte production de laccase lors de confrontations avec des Trichoderma sp.<br />
(Savoie et al., 2001). Ces résultats confirment bien l’intérêt d’addition des margines durant la<br />
préparation de l’Ebly ® pour l’élaboration du spawn, afin d’éliminer la contamination des<br />
cultures de la souche Le119 sur les substrats oléicoles (Bois de taille, grignons d’olive,<br />
margines) et d’accélérer le démarrage des points d’inoculation.<br />
II. Analyse de l’ergostérol non estérifié pour l’estimation quantitative de l’inoculum<br />
(blanc) de Lentinula edodes<br />
Contrairement à la fermentation en milieu liquide, la détermination directe de la<br />
biomasse mycélienne durant la FMS est très difficile à réaliser à cause des filaments qui<br />
restent enlacés au substrat de culture (Raimbault et Alazard, 1980). Différentes méthodes<br />
indirectes ont été proposées par plusieurs auteurs, mais ces méthodes ne sont pas très<br />
efficaces (Matcham et al., 1985 ; Bellon-Maurel et al., 2003 ; Couri et al., 2006). Dans notre<br />
travail, nous avons choisi la méthode de la quantification de l’ergostérol non estérifié pour<br />
l’estimation indirecte de la biomasse fongique de L.edodes Le119 développé sur les grains de<br />
l’Ebly ® . Cette méthode reste tributaire d’une bonne proportionnalité entre l’élément dosé et la<br />
quantité de biomasse (Sobal, 2002). Egalement la stabilité de la teneur en ergosterol des<br />
parois du mycélium en fonction de l’âge de la culture, des conditions environnementales et de<br />
la composition du milieu de culture doivent être vérifiés. Le dosage de l’ergosterol sur les<br />
différents échantillons prélevés réalisés tout au long des FMS doit permettre de suivre<br />
l’évolution de la biomasse mycélienne contenue dans les produits fermentés, mais aussi<br />
d’estimer le pourcentage de biomasse fongique présente dans le produit fermenté par rapport<br />
au substrat initial.<br />
Le travail sur l’analyse de l’ergostérol a fait l’objet d’un article à soumettre à la revue<br />
Food Chemistry.<br />
72
Résultats et discussion. Chapitre 2: Optimisation de la production du spawn<br />
Biomass estimation and physiological state evaluation of Lentinula edodes<br />
spawn by non-esterified ergosterol analysis<br />
Authors:<br />
Hicham Lakhtar 1 , Mustapha Ismaili-Alaoui 2 , S. Roussos 1*<br />
Author’s address:<br />
1 Institut Méditerranéen d'Ecologie et de Paléoécologie UMR CNRS/IRD 193; <strong>IMEP</strong> Case<br />
441; FST Saint Jérôme; Université Paul Cézanne; Av. Escadrille Normandie-Niemen; 13397<br />
Marseille cedex 20; France ;<br />
2 IAV –Hassan II, Laboratoire des Bioconversions ; BP. 6202-Instituts, 10101 Rabat, Maroc ;<br />
*Corresponding author:<br />
Sevastianos Roussos: Institut Méditerranéen d'Ecologie et de Paléoécologie UMR CNRS/IRD<br />
193; <strong>IMEP</strong> Case 441; FST Saint Jérôme; Université Paul Cézanne; Av. Escadrille<br />
Normandie-Niemen; 13397 Marseille cedex 20; France ;<br />
E-mail: s.roussos@univ-cezanne.fr<br />
Tel: (+33) 04 91 28 91 03<br />
Fax: (+33) 04 91 28 86 68<br />
73
Résultats et discussion. Chapitre 2: Optimisation de la production du spawn<br />
Analyse de l’ergosterol non estérifié pour l’stimation de la biomasse et<br />
l’evaluation de l’état physiologique de l’inoculum (blanc) de Lentinula<br />
edodes<br />
RESUME<br />
A cause de la demande accrue, la production des champignons comestibles et<br />
médicinaux connaît actuellement une expansion importante. De ce fait, la culture des<br />
champignons est devenue de plus en plus populaire à travers le monde en raison de la maîtrise<br />
au niveau industriel de leur culture sur différents substrats lignocellulosiques avec des<br />
rendements élevés. La réussite de la culture de champignons dépend essentiellement de la<br />
qualité, de l'état physiologique et de la taille de l’inoculum utilisé. Cependant, la<br />
quantification de spawn (mycélium produit sur les grains de céréales) est très difficile. Dans<br />
ce travail, la détermination de la biomasse mycélienne de L. edodes sur les grains d’Ebly ® au<br />
cours de la phase végétative est décrite pour la première fois. L’ergostérol, composé de la<br />
paroi du mycélium de L.edodes Le119, a été mesuré. L’extraction non alcaline de l’ergosterol<br />
a été réalisée dans l’éthanol à 4° C durant une nuit. L'ergostérol extrait du mycélium a été<br />
quantifié par l’HPLC en comparaison avec l’ergostérol pur comme standard. L’effet du<br />
substrat et de l’âge du mycélium des cultures a été évalué et aucune différence statistique n'a<br />
été observée pour L. edodes Le119. L’addition des margines d’olive sur les grains d’Ebly à<br />
raison de 10% (v/v) a montré l’amélioration du rendement de la biomasse de la souche de L.<br />
edodes. En outre, la respirométrie (production de CO2) de L. edodes Le119 cultivées en FMS<br />
sur des grains d’Ebly ® durant la phase végétative a été corrélée avec la teneur de l'ergostérol<br />
(r 2 = 0,80) dans le produit fermenté.<br />
Mots clé: Lentinula edodes, Ergostérol, Inoculum (spawn), Biomasse fongique, HPLC,<br />
Respirométrie, Fermentation en milieu solide, Facteur de conversion.<br />
74
ABSTRACT<br />
Résultats et discussion. Chapitre 2: Optimisation de la production du spawn<br />
Edible and Medicinal Mushroom production is nowadays under huge expansion owing<br />
to extensive demand in world Market. Thereby, mushroom culture is becoming more popular<br />
throughout the world because of the improvement of mushroom culture utilizing various<br />
lignocellulosic substrates with good yield of production. However, mushroom culture<br />
successful depends primarily to quality, physiological state and size of spawn used. While the<br />
quantification of spawn (mycelium produced on cereal grain) is very difficult to obtain. In the<br />
present work, the determination of mycelial biomass of L. edodes spawn on wheat grain<br />
Ebly® during spawn running is reported for the first time. Ergosterol of Le119 L. edodes<br />
strain mycelium was measured. Non-Alkaline extractions were performed with ethanol at 4°C<br />
for overnight. The extracted ergosterol was quantified by RP-HPLC against ergosterol pure as<br />
standard. Effect of different substrates and ages of L. edodes mycelium were evaluated. No<br />
statistical differences were exhibited for the Lentinula edodes Le119 strain. Presence of<br />
supplement on wheat grain as olive mill wastewater in 10% (v/v) exhibited enhancement of<br />
mycelial biomass yield of L. edodes strain. Furthermore, the respirometry (CO2 production) of<br />
L. edodes cultivated in solid state fermentation on wheat grain during spawn running was<br />
correlated to ergosterol content of fermented products (r 2 = 0.80).<br />
Keywords: L. edode, Ergosterol, Spawn, Fungal Biomass, HPLC, Respirometry, Solid State<br />
Fermentation, Conversion Factor.<br />
75
1. Introduction<br />
Résultats et discussion. Chapitre 2: Optimisation de la production du spawn<br />
Ergosterol is principal sterol present in fungi biomass (Czub and Baginski, 2006) with<br />
its two forms, as free ergosterol and esterified ergosterol. The relative abundances of free to<br />
esterified ergosterol are different among the various species of fungi (Czub and Baginski<br />
2006; Yuan et al., 2007). Therefore, the determination of ergosterol consists in measuring of<br />
living fungal biomass, because of the assumption of its instability after death of biomass<br />
(Mille-Lindblom et al., 2004). This ergosterol was also subject for certain investigations in<br />
order to study its conversion to Vitamin D2 (Jasinghe and Perera, 2005; 2006) which has been<br />
suggested for therapeutic applications in the treatment of several diseases including<br />
hyperproliferative diseases, secondary hyperparathyroidism, post-transplant survival, and<br />
various malignancies (Morgan, 2001).<br />
Culture of L. edodes attracks attention of several authors for its proved medicinal<br />
properties and antimicrobial effect (Kitzberger et al., 2007; Israilides et al., 2008; Rao et al.,<br />
2009) and biodegradation as well as biotransformation abilities of phenolic compound as<br />
lignin by producing large spectrum of enzymes involved in transformation of these<br />
recalcitrant compounds (Philippoussis al., 2007). While the kinetics of growth of the<br />
mycelium is related to nutritional and environmental factors, several authors (Mamiro and<br />
Royse 2008; Kuforiji and Fasidi 2009; Royse and Chalupa 2009) demonstrated that the high<br />
quality crop of mushroom depends primarily on the quality of the spawn, which in turn<br />
depends on the purity and size of biomass and physiological state of the culture. However,<br />
direct quantification of biomass during spawn running is very difficult or even impossible,<br />
owing to the problem of separating the fungal mycelium from the substrate, where the fungal<br />
hyphae penetrate into and bind tightly to the substrate (Bellon-Maurel et al., 2003; Singhania<br />
et al., 2009).<br />
However, several techniques of indirect fungal biomass in Solid state fermentation<br />
(SSF) were reported depending to measuring the biological activity (e.g respiration O2 intake<br />
and/or CO2 production) or measuring the content of certain cell components. Recently digital<br />
image processing has been developed as a tool for measuring of biomass of Aspergillus niger<br />
strains in SSF (Couri et al., 2006). The growth of this strain was quantified as the total area<br />
occupied by the hyphae, while this technique was poorly correlated with glucosamine test.<br />
The drawback of this technique consists in setting up of the focus and illumination parameters<br />
which become more difficult when complex substrates are used (Couri et al., 2006).<br />
76
Résultats et discussion. Chapitre 2: Optimisation de la production du spawn<br />
Therefore, to our knowledge, there is no report on estimation of quantity and quality of<br />
biomass during the production of medicinal Lentinula edodes spawn on wheat grain. In the<br />
present study, we report for the first time ergosterol-biomass conversion factor in spawn of L.<br />
edodes. Non-alkaline protocol of extraction of ergosterol is developed for estimation of both<br />
quantity and quality ways of L. edodes biomass during spawn running. Effect of substrate of<br />
culture and age of mycelium were evaluated in model system to determine the quantity of<br />
spawn. The correlation between ergosterol levels and CO2 production was determined to<br />
estimate the quality of L. edodes spawn.<br />
2. Materiel and methods<br />
2.1. Microorganism<br />
Lentinula edodes Le119 was previously selected among collection of sixteen strains<br />
for its capability of colonization of olive wastes. The preserved stock culture of strain was<br />
maintained in sterilized Potato Dextrose Agar (PDA) (39 g.L -1 , Sigma, France) with 10% of<br />
Olive mill wastewater (OMW) (Lakhtar et al., 2009) with periodic transfer of one month.<br />
2.2. Culture Media<br />
2.2.1. Liquid state cultivation<br />
Liquid cultures were grown on Malt Extract medium (ME) (30 g.L -1 , Sigma, France)<br />
and on diluted OMW (10% of v/v) without any supplements. The culture media (100 mL)<br />
were sterilized (121°C, 20 min) and then inoculated with three plugs (1 cm diameter) of<br />
earlier uniformly colonized PDA of one week. Medium was incubated at 25°C under shaking<br />
(120 rpm). Triplicate flasks were harvested for determination of mycelial biomass dry weight<br />
and ergosterol content. Mycelium was harvested from the liquid medium by filtration<br />
(Watman filter N°1) and frozen dried.<br />
2.2.2. Solid state Cultivation on model system<br />
Three different culture media were used for solid culture: PDA, PDA supplemented<br />
with 10% of OMW and Ebly Agar (EbA) consisting of 24 g of wheat grain (Ebly®, Casino,<br />
France), which were milled to powder, and 15 g of agar in 1 mL of distillated water. These<br />
media were used to determine ergosterol content in mycelial biomass of L. edodes Le119<br />
strain. Biomass and ergosterol content was estimated using sterilized cellophane disc placed<br />
on the agar surface from Petri dishes (diam. 90 mm) (De Araujo et al., 2002). After<br />
colonization, the mycelium was separated from the substrate and cellophane membrane. It<br />
77
Résultats et discussion. Chapitre 2: Optimisation de la production du spawn<br />
was then cut out and frozen dried. Each part of mycelial colony with different ages was tested<br />
to quantify ergosterol content (Fig. 1).<br />
1 2 3 4<br />
Figure 1. Mycelium growth of L.edodes Le119 on solid medium with cellophane membrane<br />
incubated at 25°C, 1: Inoculum (6 mm), 2: with mycelium 20 days old, 3: with mycelium 12<br />
days old, 4: with mycelium 5 days old.<br />
2.3. Solid state fermentation of L. edodes on wheat grain<br />
Tow spawn kinds of L. edodes Le119 strain were prepared. White spawn (WS) was<br />
prepared as described by Sobal (2002) with modification. Quantity of 200 g of wheat grain of<br />
Ebly® were humidified with distilled water to 50% (w/w). While the brown spawn (BS), was<br />
prepared with addition of OMW; 200 g of wheat was wetted with 10% of OMW to get 50%<br />
(w/w) of humidity, heated with microwave for 5 minutes to allow absorption of OMW by<br />
Ebly®. The substrates were autoclaved for 30 min at 121°C and allowed to cool for 24h. Pre-<br />
weighed “Raimbault columns” (De Araujo et al., 1997) were filled with sterilized Ebly®<br />
alternated by inoculum (three plugs of 5 mm) cut from PDA previously colonized with Le119<br />
strain of 1 week. Forced aeration of the substrate was regulated to 25 mL.min -1 (which<br />
allowed five time of air renewal in column per hour). The CO2 production was recorded using<br />
Infra-red detector which was calibrated with air containing 1 and 5% of CO2.<br />
2.4. Extraction of non esterified ergosterol<br />
Pure mycelium and spawn (Ebly® fermented) samples were frozen-dried to constant<br />
weight and milled to a uniform fine powder using grinder (IKA Labortechnik, Germany).<br />
Ergosterol was extracted from 50 mg samples of pure mycelium and spawn by<br />
homogenization in 1 mL of methanol. The mixtures were vortexed for 1 minute and<br />
subsequently shaken vigorously and then incubated at 4°C for overnight in the dark.<br />
Supernatants were recovered by centrifugation for 10 minutes at 14 000 rpm. The supernatant<br />
was then filtered through 0.2 µm syringe filter (Iso-Disc TM Filters, USA).<br />
78
Résultats et discussion. Chapitre 2: Optimisation de la production du spawn<br />
2.5. Recovery of ergosterol from spawn spiked with ergosterol and mycelium.<br />
Assessment of the recovery efficiency of ergosterol was carried out by artificially<br />
addition of pure ergosterol (from Sigma) or mycelium which its ergosterol content was<br />
already determined [3.55 mg (ergosterol).g -1 (mycelium)]. Recovery of pure ergosterol was<br />
determined after mixing 0.5 mL of methanol containing 100 µg dissolved ergosterol with 10 g<br />
(dry weight basis) of Le119 spawn. After mixing, sub-samples were extracted using the<br />
described procedures and ergosterol was determined by HPLC. For recovery of fungal<br />
ergosterol, portion of 100 mg of pure mycelium were mixed with 10 g (dry weight) of spawn.<br />
After mixing, sub-samples were extracted using the described procedures and the amount of<br />
ergosterol was determined.<br />
2.6. HPLC analysis<br />
Final separation of ergosterol from extract matrix was performed by reversed-phase<br />
high-performance liquid chromatography (RP-HPLC). The chromatographic systems used<br />
were an Agilent Series 2000. Volume injection was 20 µl of sample. The chromatographic<br />
separation was achieved on 5 µm dC18 (4.6x250 mm) reversed-phase column (Atlantis,<br />
Ireland) and detection was set at 282 nm. Elution of ergosterol at column temperature of 40°C<br />
was performed with isocratic run with 100% methanol at a flow rate of 1 mL.min -1 . Peak<br />
identity was checked on the basis of retention times of pure ergosterol. Correlation between<br />
dry biomass and ergosterol content was verified by measuring the peak height and area. UV<br />
spectrum of ergosterol extracted from spawn was recorded using the same apparatus from 265<br />
to 300 nm. All experiments were carried out in triplicate.<br />
2.7. Statistical Analysis<br />
One way analyses of variance were tested for effects of medium composition and<br />
culture age on the ergosterol concentration in mycelium. The significance level was set at P =<br />
0.05. The correlation coefficient was used to determine the relationship two properties<br />
(biomass and ergosterol; ergosterol and CO2 production). The analysis were performed using<br />
Xlstat® software.<br />
79
3. Results<br />
Résultats et discussion. Chapitre 2: Optimisation de la production du spawn<br />
The present study was aimed to evaluate ergosterol from mycelium of a basidiomycete<br />
(Lentinula edodes), for indirect estimation of quantity of biomass culturing in solid state<br />
fermentation for production of spawn on Ebly®. The effects of medium of culture and age of<br />
mycelium were evaluated in liquid state and in agar plate covered with cellophane membrane<br />
as model system for solid state culture. This system allows the growth of mycelium and<br />
prevents penetration of the mycelium into the substrate of the culture.<br />
3.1. Growth curves in liquid state culture.<br />
The growth curves of L. edodes cultivated on different media in liquid state culture are<br />
shown in Fig. 2. The highest ratio of biomass was obtained on ME (102 ±6.0 mg.L -1 ).<br />
However, the ergosterol content (3.6 mg.g -1 dw) of L. edodes in both media (ME and 10% of<br />
OMW) was not statistically different during cultivation time.<br />
Dry weight of mycelium (mg)<br />
150<br />
130<br />
110<br />
90<br />
70<br />
50<br />
30<br />
10<br />
-10<br />
0 7 14<br />
Incubation Time (day)<br />
21 28 0<br />
Figure 2. Time course of biomass production (solid lines) and ergosterol content (mg.g -1 dw)<br />
(dashed lines) on ME ( ) and OMW ( ) at 25°C.<br />
4,5<br />
4<br />
3,5<br />
3<br />
2,5<br />
2<br />
1,5<br />
1<br />
0,5<br />
Ergosterol content mg.g -1<br />
80
Résultats et discussion. Chapitre 2: Optimisation de la production du spawn<br />
3.2. Conversion factor of Lentinula edodes in solid state culture:<br />
In solid state culture, the biomass produced in PDA, PDA supplemented with 10% of<br />
OMW and EbA is shown in Table1. Maintain of mycelium on agar slants in the presence of<br />
OMW was successful to produce high biomass yield compared to that produced in PDA (268<br />
and 75 mg respectively at 20 days of incubation), while the biomass produced in EbA was<br />
moderate (105 mg at 20 days of incubation). EbA was used to determine conversion factor<br />
(ergosterol content in mycelium) in medium that contain the substrate used for production of<br />
spawn. The agar media used were analyzed and were found not to contain ergosterol (Data<br />
not shown). Differences among media were not significant when all data were analyzed<br />
together. In 20-day old mycelium, ergosterol content exhibited the highest variability in all<br />
culture media (Table1).<br />
Table1. Effect of age and substrate on mycelial ergosterol contents<br />
Medium Culture age Biomass Ergosterol content<br />
(days) (mg)<br />
(mg.g -1 of dry weight)<br />
PDA 5 20 3,46 ± 0,28<br />
PDA +10 %<br />
of OMW<br />
12 45 3,72 ± 0,32<br />
20 75 3,17 ± 0,74<br />
5 75 3,97 ± 0,21<br />
12 170 3,82 ± 0,54<br />
20 268 3,32 ± 1,02<br />
EbA 5 43 3,64 ± 0,45<br />
12 65 3,71 ± 0,23<br />
20 105 3,32 ± 0,95<br />
Straight line relationship between fungal dry biomass and ergosterol content was<br />
observed and the statistical analysis proved a high correlation (r 2 =0.99) (Fig. 3). Under the<br />
HPLC conditions tested, the calibration curve was established by plotting the peak area and<br />
peak height against ergosterol standard.<br />
81
Ergosterol (mg)<br />
Résultats et discussion. Chapitre 2: Optimisation de la production du spawn<br />
0,2<br />
0,16<br />
0,12<br />
0,08<br />
0,04<br />
0<br />
y = 0,003x - 0,000<br />
R² = 0,999<br />
0 10 20 30 40 50<br />
Figure 3. Relationship between fungal dry weight and ergeosterol content, for L. edodes<br />
Le119 cultivated in solid state culture with cellophane system, incubated during 20 days at<br />
25°C.<br />
3.3. Recovery of spiked ergosterol:<br />
The recovery of methanol-dissolved ergosterol from spawn was estimated to 75-80%<br />
for both in brown and white spawn respectively, and the recovered value did not differ<br />
significantly (Table 2). However, 90% of recovery of fungal ergosterol was found for Le119<br />
strain.<br />
Biomass dry weight (mg)<br />
Table 2. Recovery of Sigma pure ergosterol and fungal ergosterol (homogenized<br />
mycelium of L. edodes Le119) added to Brown and White spawn<br />
Samples Recovery of pure<br />
ergosterol µg.g -1<br />
Brown Spawn 7 .52 3.17a 1<br />
White spawn 8.04 3.21a<br />
Added amount 2 10.0 3.55b<br />
Recovery of fungal<br />
ergosterol<br />
1 Different letters indicate significant differences.<br />
2 Added amount of ergosterol for Le119 was based on direct measurements of ergosterol in<br />
the mycelium using described protocol for extraction.<br />
82
Résultats et discussion. Chapitre 2: Optimisation de la production du spawn<br />
3.4. Growth curves of spawn<br />
The growth curves of L. edodes Le 119 shiitake strains on different preparation of<br />
Ebly® (white and brown spawn) presented by ergosterol content and CO2 production by<br />
mycelium are shown in Figure 4. On Ebly® supplemented with 10% OMW, the biomass<br />
produced, which was represented and determined by measuring ergosterol contained in<br />
spawn, was higher than on wheat without supplementation [0.6 and 1.8 mg (biomass).g -1 dw<br />
(spawn) respectively after 20 days of incubation]. These results indicated that presence of<br />
OMW is more efficient for spawn production of Le119 strain of L. edodes. At the end of the<br />
growth period, high variation of ergosterol was observed for both media.<br />
CO2 production (mL .h -1 .g -1 dw)<br />
Time (day)<br />
Figure 4. Time course of biomass production (mg.g -1 substrate dw) (solid lines) and CO2<br />
production (dashed lines) of Le119 strain on wheat with ( , ) and without OMW ( , ) at<br />
25°C.<br />
The time courses of CO2 production were similar for all two media. Strong CO2<br />
production was observed after 6-8 days until 14-16. After this period, the amount of CO2<br />
produced was slightly increased. The mycelial biomass produced on both media was<br />
estimated using conversion factor of 281 (f = 1/3.56 x 1 000, average of factor conversion<br />
found by culturing of mycelium on EbA). Estimating ergosterol in spawn preparation, close<br />
correlation coefficient between ergosterol content in spawn and CO2 production was found for<br />
the both media WS (r 2 = 0.965 and 0.956 for BS and WS respectively) (Fig. 5).<br />
Ergosterol content (mg.g -1 )<br />
83
Ergosterol content (mg.g -1 )<br />
Résultats et discussion. Chapitre 2: Optimisation de la production du spawn<br />
Figure 5. Correlation between ergosterol content and CO2 production by L. edodes Le119 in<br />
WS ( ) and BS ( ) incubated at 25°C during 20 days.<br />
4. Discussion<br />
CO2 production (mL .h -1 .g -1 dw)<br />
Estimation of fungal biomass in solid state fermentation was subject of several studies<br />
(Ooijkaas et al., 1998; Scotti et al., 2001; Niemenmaa et al., 2008). Therefore, respiration<br />
measurement (O2 consumption or/and CO2 production) and biochemical analysis of cell<br />
components were evaluated to find a good indicator for biomass estimation. Thus, the<br />
combination of different indicators was carried out in order to offer a significant improvement<br />
of correlation between real biomass and which was estimated (Matcham et al., 1985; Ooijkaas<br />
et al., 1998). Whereas, higher the number of indicators, the more difficult to interpret the<br />
results and the conversion of growth become totally impracticable.<br />
In our study, ergosterol was tested for estimation of L. edodes biomass cultivated in<br />
solid state fermentation to produce spawn of L. edodes which is the main factor of the<br />
attainment for high quality crop of mushroom (Mamiro and Royse 2008; Kuforiji and Fasidi<br />
2009; Royse and Chalupa 2009). Ergosterol content showed close correlation to mycelial<br />
growth (r= 0.987). Similar result was obtained for Agaricus bisporus (Matcham et al., 1985)<br />
which was cultivated on different media in solid and liquid state culture, while it was in<br />
disagreement with Coniothyrium minitans produced on PDA (Ooijkaas et al., 1998).The<br />
explanation could be that the spore production of C.minitans affects the ergosterol content<br />
which in return was depending to the nature of substrate. Thus the determination of ergosterol<br />
could present a good result for estimation of basidiomycete mycelium.<br />
84
Résultats et discussion. Chapitre 2: Optimisation de la production du spawn<br />
The substrates used for L. edodes growth in both liquid and solid state culture exhibited<br />
constant ergosterol content in mycelium. Mean value of ergosterol content in pure mycelium<br />
3.57± 0.56 (mg.g-1 of dry weight) was within the range found in other experiments with other<br />
microorganism (Pasanen et al., 1999; Barajas-Aceves 2000; Jasinghe and Perera 2005). While<br />
some of fungal species exhibited large differences in ergosterol content between the isolates,<br />
as can be seen from the relatively large standard deviation (Klamer and Bååth 2004; Mille-<br />
Lindblom et al., 2004; De Ridder-Duine et al., 2006).<br />
It would appear that culturing of mycelium on EbA could allow for operating nearly to<br />
natural condition so as to gain insight into the magnitude of variation and hence the potential<br />
error when calculating fungal biomass from ergosterol concentration measured in spawn<br />
fermentation (Gessner and Chauvet 1993). Determination of ergosterol allowed for reducing<br />
the use of hazardous organic solvents which involved in saponification step of ergosteryl ester<br />
(De Ridder-Duine et al., 2006), and this thereby permeates for measuring of living fungal<br />
biomass (Mille-Lindblom et al., 2004) based on the assumption that the ergosterol have been<br />
considered as kind of lipids storage (Klamer and Bååth 2004). Furthermore, the recovery of<br />
ergosterol with this simple protocol was exhibited in the most of case very high as could be<br />
reported by several authors (Gong et al., 2001; De Ridder-Duine et al., 2006; Yuan et al.,<br />
2007).<br />
The identification of ergosterol with HPLC was achieved by comparing its retention<br />
time and spectrum against ergosterol standard (Fig. 6). The ergosterol spectrum exhibited<br />
characteristic pattern, because it has a conjugated pair of double bonds at carbons 5-6 and 7-8,<br />
it absorbs UV light strongly between 240 and 300 nm. The interference of sterol containing in<br />
wheat, as reported by Seitz et al. (1977), was eliminated by verification of purity of ergosterol<br />
peak detected by HPLC, and verification of spectrum of peak corresponding to ergosterol.<br />
The conversion of ergosterol, which was considered as precursor, to vitamin D2<br />
(Ergocalciferol) by UV irradiation was reported by several authors (Jasinghe and Perera 2005;<br />
Jasinghe and Perera 2006; Teichmann et al., 2007). Certain author investigated conversion<br />
kinetics of ergosterol to vitamin D2 in natural products such as in L. edodes mushrooms so as<br />
to modeling of conversion in order to predict the yield of vitamin D2 after the irradiation<br />
process.<br />
85
D.O (mAU)<br />
Résultats et discussion. Chapitre 2: Optimisation de la production du spawn<br />
Figure 6. UV spectrum (260- 300 nm) of ergosterol extracted from mycelium of L. edodes<br />
Le119 strain grown on Ebly ® in solid state fermentation.<br />
5. Conclusion<br />
The results presented in this paper showed that the non-esterified ergosterol<br />
measurement of L. edodes can be considered as a reliable indirect method for estimating<br />
fungal biomass present in spawn. In fact, non-esterified ergosterol content is constant<br />
whatever the age of the mycelium, culture conditions and medium composition. In the other<br />
hand, this study demonstrated that the established correlations between Ergosterol and CO2<br />
rate production in solid state fermentation permit a good estimation of the physiological state<br />
of mycelial biomass. By using the linear regression equation obtained in solid state culture,<br />
quantities of non-esterified ergosterol measured could be easily converted into fungal dry<br />
matter quantities per gram of dry solid medium (1 g of mycelial biomass contain 3.57 mg of<br />
ergosterol). To optimize a solid state fermentation process for biomass production, the use of<br />
such method cannot be avoided.<br />
Wavelength (nm)<br />
86
Résultats et discussion. Chapitre 3: Evaluation des facteurs de croissance<br />
Chapitre <strong>III</strong><br />
Culture de Lentinula edodes en fermentation en milieu solide sur<br />
substrat oléicole solide: Evaluation des facteurs de croissance<br />
Dans les chapitres précédents, nous avons optimisé la préparation de l’inoculum<br />
(spawn) de L. edodes Le119 sur les grains de l’Ebly® par l’addition des margines à 10%.<br />
L’ajout des margines a permis de produire un inoculum avec une biomasse mycélienne active,<br />
abondante et induite pour la production des laccases, permettant ainsi d’éliminer la<br />
contamination des cultures par des moisissures indésirables. Après cette étape cruciale qui<br />
conditionne la réussite de la culture de L. edodes sur les substrats oléicoles, nous avons<br />
cherché à optimiser les conditions de culture de L. edodes Le119 sur un mélange optimisé, de<br />
bois de taille, grignons d’olive et de margines, qui va constituer tout le long de ce travail le<br />
Substrat Oléicole Solide (SOS).<br />
Avant toute démarche d’optimisation des conditions de culture d’un microorganisme,<br />
la première étape consiste en la détermination des facteurs susceptibles d’avoir une influence<br />
sur la culture. Plusieurs méthodologies ont été rapportées dans la littérature, entre autres la<br />
méthode ‘Une variable à la fois’ et la méthode de plan d’expérience (Mathieu et Phan-Tan-<br />
Luu, 1997). La méthode ‘Une variable à la fois’ consiste à fixer tous les facteurs sauf un, pour<br />
connaître son effet sur la réponse. Elle est coûteuse en nombre d'essais, et inefficace : elle ne<br />
permet pas d'optimiser le processus ni de trouver un modèle prédictif s'il existe des<br />
interactions entre les facteurs. Dans un plan d'expérience au contraire, toutes les données sont<br />
87
Résultats et discussion. Chapitre 3: Evaluation des facteurs de croissance<br />
utilisées simultanément pour calculer le poids de chaque effet, d'où une précision accrue et un<br />
faible nombre d'essais.<br />
Plusieurs matrices d’expériences ont été utilisées selon l’objectif de l’étude et selon la<br />
qualité d’information recherchée (Mathieu et Phan-Tan-Luu, 1997). Dans notre étude, nous<br />
avons sélectionné une matrice factorielle fractionnaire asymétrique, ayant 5 facteurs (pH<br />
initial du substrat, débit d’aération, âge de l’inoculum, taux de l’inoculum et taille des<br />
particules) avec deux niveaux et un facteur (solution minérale) avec 5 niveaux sur 16<br />
expériences (2 5 5 1 //16).<br />
Dans un premier temps, l’approche expérimentale a consisté en la détermination d’un<br />
mélange optimal de bois de taille, grignons d’olive et de margines permettant une bonne<br />
production de la biomasse mycélienne de L. edodes Le119. Dans un deuxième temps, on a<br />
évalué l’influence des six facteurs, définis auparavant, sur la biomasse mycélienne produite<br />
sur le mélange optimisé et sur la dégradation des phénols totaux du mélange en fermentation<br />
en milieu solide dans des colonnes de Raimbault.<br />
Technology.<br />
Ce chapitre a fait l’objet d’un article à soumettre dans la revue Bioresource<br />
88
Authors:<br />
Résultats et discussion. Chapitre 3: Evaluation des facteurs de croissance<br />
Solid state fermentation of Lentinula edodes on solid olive substrate:<br />
Evaluation of growth factors.<br />
Hicham Lakhtar 1 , Mustapha Ismaili-Alaoui 2 , Sévastianos Roussos 1*<br />
Author’s address:<br />
1 Institut Méditerranéen d'Ecologie et de Paléoécologie UMR CNRS/IRD 193; <strong>IMEP</strong> Case<br />
441; FST Saint Jérôme; Université Paul Cézanne; Av. Escadrille Normandie-Niemen; 13397<br />
Marseille cedex 20; France ;<br />
2 IAV –Hassan II, Laboratoire des Bioconversions ; BP. 6202-Instituts, 10101 Rabat, Maroc ;<br />
*Corresponding author:<br />
Sevastianos Roussos: Institut Méditerranéen d'Ecologie et de Paléoécologie UMR CNRS/IRD<br />
193; <strong>IMEP</strong> Case 441; FST Saint Jérôme; Université Paul Cézanne; Av. Escadrille<br />
Normandie-Niemen; 13397 Marseille cedex 20; France ;<br />
E-mail: s.roussos@univ-cezanne.fr<br />
Tel: (+33) 04 91 28 91 03<br />
Fax: (+33) 04 91 28 86 68<br />
89
Résultats et discussion. Chapitre 3: Evaluation des facteurs de croissance<br />
Culture de Lentinula edodes en fermentation en milieu solide sur substrat<br />
oleicole solide: Evaluation des facteurs de croissance<br />
RESUME<br />
L’objectif de l’étude a été d’optimiser la culture de Lentinula edodes Le119 en<br />
fermentation en milieu solide sur les substrats oléicoles composés de bois de taille, des<br />
grignons d’olive et des margines constituant ainsi un substrat oleicole solide destiné à la<br />
culture du mycélium. Tout d’abord, la formule optimale de mélange de substrats oléicoles<br />
pour la fermentation en milieu solide de la souche de L. edodes Le119 a été déterminée. Par la<br />
suite, les facteurs de croissance (pH initial, aération, âge d’inoculum, taux d’inoculum, taille<br />
de particules, et solution minérale) ont été évalués suivant la méthodologie de plan de criblage<br />
sous la forme d’un plan d’expérience factoriel factionnaire 2 5 5 1 //16 expériences. Les résultats<br />
de la première série d’expériences ont montré que la quantité de chaque composant du<br />
mélange est la suivante ; 20% de bois, 30% de grignons d’olive et 50% de margines. Ce<br />
mélange permet une croissance abondante du mycélium et une production de la biomasse<br />
mycélienne de 16,24 mg.g -1 substrat poids sec (SPS). Les résultats de la deuxième série<br />
d’expériences ont montré que l’addition de la solution minérale exerce un effet négatif sur la<br />
production de la biomasse mycélienne ainsi que la dégradation des phénols totaux, tandis que<br />
le taux d’inoculum, utilisée pour démarrer les cultures, exerce un effet positif sur les deux<br />
réponses. La croissance mycélienne de L. edodes Le119 a montré une bonne corrélation avec<br />
la réduction des phénols totaux (r 2 = 0,81, P< 0.001). Les conditions optimisées de culture de<br />
L. edodes Le119 sur SOS avec 10% (p/p) d’inoculum et sans addition de solution minérale a<br />
abouti à la production de la biomasse mycélienne de 30,44 mg.g -1 SPS, à l’absence de<br />
contamination de culture et à 48% de réduction en phénols totaux initialement présents dans<br />
les margines.<br />
Mots clé : Bois de la taille, Grignons d’olive, Margines, Fermentation en milieu solide,<br />
Planification expérimentale, Optimisation, Dégradation des phénols.<br />
90
ABSTRACT<br />
Résultats et discussion. Chapitre 3: Evaluation des facteurs de croissance<br />
The aim of the present work was to optimize the conditions of solid state fermentation<br />
of Lentinula edodes on solid olive substrate (SOS) including olive twigs and leaves (OTL),<br />
olive cake (OC) where olive mill wastewater (OMW) acts as the wetting/moistening agent. To<br />
do so, the optimal formulation of substrate for L. edodes Le119 strain culture was determined.<br />
In the other hand, the growth factors (Initial pH, aeration, inoculum age, inoculum size<br />
particle size and mineral solution) were evaluated following the experimental screening<br />
design methodology using 2 5 5 1 //16 experiments. The results of a first series of experiments<br />
showed the optimal formulation of substrate for Le119 strain was OTL (20%) + OC (30%) +<br />
OMW (50%) allowing mycelial biomass production of 16,24 mg.g -1 of substrate dry weight.<br />
For a second series of experiments, the results showed that the nutriment added to the<br />
substrate exercised a negative effect on mycelial growth of L. edodes Le119 strain and total<br />
phenol removal of substrate, while the spawn size exercised a positive effect on both<br />
responses. The mycelial growth appeared to be in correlation with reduction of total phenols<br />
(r 2 = 0.81, P< 0.001). Optimized conditions of Le119 culture on SOS with 10% of spawn and<br />
without solution mineral led to production of mycelial biomass of 30.44 mg.g -1 of substrate<br />
dry weight, absence of culture contamination and to a 48% drop in total phenols initially<br />
present in OMW.<br />
Keywords: Olive twigs and leaves, Olive cake, Olive mill wastewater, Solid state<br />
fermentation, Experimental design methodology, Optimization. Phenol degradation.<br />
91
1. Introduction<br />
Résultats et discussion. Chapitre 3: Evaluation des facteurs de croissance<br />
The importance of olive oil mill industry in Mediterranean countries is well known, as<br />
well as the serious problems that olive mill factories have in disposing their by-products<br />
(Cayuela et al., 2006, Altieri and Esposito 2008). Commonly, olive mills produce residual<br />
solids, the olive cake (OC) which make up a fibrous lignocellulosic waste and generally used<br />
as fuel (Ait Baddi et al., 2009); and a black liquid effluent, the olive mill wastewater or<br />
(OMW), which is a strong pollutant mainly due to its high phenolic content and organic load<br />
(Mekki et al., 2007). In addition, it has been estimated that each olive tree could produces 25<br />
kg leaves and twigs per year (Fayed et al., 2009).<br />
In the past, several investigations were carried out to select microorganisms (notably<br />
fungi) capable of eliminating the inhibitory effects of those substances and of converting the<br />
initial waste either directly into a useful end-product or by making it susceptible to further<br />
physicochemical and biological treatments (Jaouani et al., 2003; Lakhtar et al., 2009). Among<br />
Basidiomycotina in particular, white-rot fungi belonging to the genus Pleurotus and Lentinula<br />
were selectively tested for their ability to decompose the lignin and are among the most<br />
efficient producers of lignocellulosic enzymes (Elisashvili et al., 2008). In addition, they can<br />
be cultivated on a large lignocellulosic substrates transforming them into food and feed<br />
(Zhang et al., 1995).<br />
Lentinula edodes (Berk.) Pegler (shiitake) is the second most popular edible<br />
mushroom in the world because of its flavor, taste and quality ((Tsai et al., 2009)). In<br />
addition, L. edodes is one of the best known and the best characterized mushrooms used for<br />
medicinal purposes (Hadar and Dosoretz 1991; Hirasawa et al., 1999; Hatvani 2001; Gu and<br />
Belury 2005; Israilides et al., 2008; Shen et al., 2009). Owing to L. edodes characteristic,<br />
different substrate formulations have been developed in different countries, depending on<br />
their readily available raw material. Agricultural wastes such as oak, cereal straw, corn cobs,<br />
sugarcane bagasse are used alone or in combination with other wastes in L. edodes cultivation<br />
(Sunchez et Royse 2001; Philippoussis et al., 2003; Kalmis et Sargin 2004; Özçelik and<br />
Peksen 2007; Philippoussis et al., 2007). However, for the first time, the use of mixture of<br />
olive cake, olive twigs and leaves supplemented with OMW is reported.<br />
To determine the parameters affecting biomass production of L. edodes and<br />
polyphenol removal from OMW, with the intention to optimize the conditions of solid state<br />
fermentation, the set of factors influencing the growth of L. edodes on olives residues was<br />
92
Résultats et discussion. Chapitre 3: Evaluation des facteurs de croissance<br />
studied. Asymmetrical screening design was established and processing the results with the<br />
Nemrodw software (New Efficient Methodology for Research using Optimal Design) enabled<br />
us to quantify the direct effects of the various factors on the responses chosen and to identify<br />
those that will require fine-tuning during the optimization phase.<br />
2. Materials and methods<br />
2.1. Microorganism and spawn preparation<br />
Single strain of L.edodes (Le119) was used in the experiments. The strain was<br />
maintained at 4 °C on potato dextrose agar (PDA, Sigma, France) supplemented with 10% of<br />
OMW (Lakhtar et al., 2009). Wheat grains (Ebly®, Casino, France) were used as substrate<br />
and growth support for spawn preparation. The grains were cooked for 5 min in a water<br />
solution containing 10% (v/v) of OMW at a ratio of 1:1 (Ebly®: Solution; w: v). Once<br />
cooked, the wheat grains were packaged (250 g of wheat grains in jar of 250 ml) and<br />
sterilized in an autoclave at 121 °C for 30 min. After sterilization, the grains were inoculated<br />
with 5 agar disks of 5 mm diameter containing mycelium and then placed in “Raimbault<br />
columns” (De Araujo et al., 1997) and incubated at 25±2 °C in the absence of light for a<br />
periode of 10 and 25 days.<br />
2.2. Preparation of substrates<br />
The substrates were composed of different parts; OC, olive twigs and leaves (OTL)<br />
and OMW as wetting agent. OC was dried in greenhouse retaining 10% of humidity (w/w).<br />
OC and OMW used throughout the study were taken from a three phase system used in the<br />
conventional press type olive oil mill plant (Beni-Mellal, Morocco). The OC and OTL were<br />
dried in solar greenhouse with 10% (w/w) of moisture retained. OMW was stored at -20°C<br />
until it was used. OMW dilutions were performed in distillate water, unless otherwise stated.<br />
The OTL were ground and sieved to desirable particle size. The individual ingredients of the<br />
substrate were mixed thoroughly by mechanical means to achieve a homogenous mixture.<br />
2.3. Formulation of substrate for L. edodes culture<br />
In order to determine the optimal formulation of substrates allowed for high mycelia<br />
biomass, different mixtures of OC and OTL (max. particle size: 5 mm) were soaked with 50%<br />
(v/v) of OMW to obtain 50% of moisture (Table 1). Sterilization was carried out at 121°C for<br />
30 min. After cooling, 5g of grains of wheat spawn (15 days age) added to 45 g of sterile<br />
substrate were gently mixed and placed in sterile 200 mL Erlenmeyer flask. The Erlenmeyer<br />
flasks were then incubated at 25±2 °C for 1 month. Mycelial biomass was estimated using<br />
93
Résultats et discussion. Chapitre 3: Evaluation des facteurs de croissance<br />
ergosterol analysis as reported previously. At least three Erlenmeyers flasks per treatment<br />
were evaluated.<br />
Table 1. Mixing ratio of olive twigs and leaves (OTL), olive cake (OC) and olive mill<br />
wastewater (OMW) used as substrate for L.edodes growth.<br />
Treatment No Ingredient<br />
OTL (%) Olive cake (%) OMW (%)<br />
1 0 50 50<br />
2 20 30 50<br />
3 30 20 50<br />
4 50 0 50<br />
2.4. Experimental methodology<br />
2.4.1. Asymmetrical screening design<br />
For systems with a great number of variables different approaches of experimental<br />
factorial designs can be applied to achieve a screening of critical variables and to estimate<br />
their main effects on the responses. Inspiration was drawn from previous works (Baçaoui et<br />
al., 2001; Alinsafi et al., 2005; El Hajjouji et al., 2008). In this study, a 2 5 5 1 //16 experimental<br />
design was processed using Nemrodw software (Mathieu et al., 2000). It allowed the<br />
investigation of six factors (A-F) in sixteen experiments, five factors A-E each at two levels<br />
and one factor F at five levels. Tables 2 and 3 listed the values given to each factor; the choice<br />
was based on our preliminary experiments. Table 4 showed the 2 5 5 1 //16 experimental design.<br />
The results treatment was processed using Nemrodw software.<br />
From the 16 runs we can compute, using the least square method (Baçaoui et al., 2001;<br />
El Hajjouji et al., 2008), the ‘‘weight’’ of each factor level. For each factor, the weight of<br />
each level is related to the upper level weight, which becomes the ‘‘reference state’’ among<br />
each factor (Mathieu and Phan-Tan-Luu, 1997). The weight describes the factor effects on the<br />
response when changing factor levels with respect to the reference state. The obtained results<br />
are generally presented as histograms which graphically illustrate the variable differential<br />
weights (Lewis et al., 1999).<br />
94
Résultats et discussion. Chapitre 3: Evaluation des facteurs de croissance<br />
Table 2. Particle size and Composition of mineral solution used in experimental conditions<br />
Particle size<br />
0.8
Résultats et discussion. Chapitre 3: Evaluation des facteurs de croissance<br />
Table 4. Experimental conditions of the screening design with 6 factors (A-F).<br />
Run No A B C D E F<br />
1 5 20 25 20 T2 SO<br />
2 6 30 10 10 T1 SO<br />
3 5 20 25 20 T1 MS1<br />
4 6 30 10 10 T2 MS1<br />
5 5 20 10 10 T2 MS3<br />
6 6 30 25 20 T1 MS3<br />
7 5 20 10 10 T1 MS2<br />
8 6 30 25 20 T2 MS2<br />
9 5 30 25 10 T2 MS4<br />
10 6 20 10 20 T1 MS4<br />
11 5 30 25 10 T1 MS3<br />
12 6 20 10 20 T2 MS3<br />
13 5 30 10 20 T2 MS1<br />
14 6 20 25 10 T1 MS1<br />
15 5 30 10 20 T1 SO<br />
16 6 20 25 10 T2 SO<br />
2.5. Methods of analysis<br />
2.5.1. Standard chemical analysis<br />
The pH value of the sample of substrates was assayed following dilution of 1 g in<br />
10 mL of distilled water, and by using a digital pH meter. Mycelial biomass was measured by<br />
determination of non esterified ergosterol. A sample of 50 mg of frozen-dried sample was<br />
used to extract non esterified ergosterol which was determined with HPLC. Pure ergosterol<br />
(Sigma, France) was used as standard. The total phenol was determined as reported by<br />
Lakhtar et al. (2009). Five replicates per treatment were used.<br />
2.5.2. Elemental analysis<br />
The instrument used for carbon and nitrogen concentrations was a Flash EA 1212<br />
Elemental Analyzer (France). About 1 mg (0.8 - 1.2 mg) of the sample in a capsule made of<br />
silver (assay of C and N) was burned at 920-1000°C. The quantity of each element is<br />
expressed in percent mass. Each determination was performed in duplicate. The instrument<br />
was calibrated with aspartic acid standard.<br />
96
Résultats et discussion. Chapitre 3: Evaluation des facteurs de croissance<br />
2.5.3. 13 C-nuclear magnetic resonance ( 13 C-NMR) analysis<br />
Freeze-dried samples were used directly in NMR experiments without special<br />
preparation. Owing to their size, the samples were gently ground (IKA Labortechnik,<br />
Germany) in order to fill the rotor. Solid-state 13 C CPMAS NMR spectra were obtained on a<br />
Bruker Avance-400 MHz spectrometer (Bruker, Bremen, Germany) operating at a 13 C<br />
resonance frequency of 100.7 MHz. Samples were placed in a 7-mm zirconium rotor and spun<br />
at the magic-angle at 6 kHz. All measurements were made at room temperature. The 13 C<br />
chemical shifts were referred to tetramethylsilane and calibrated with the glycine carbonyl<br />
signal, set at 172.5 ppm. Deconvolution of the NMR spectra was performed using the DmFit<br />
software (Massiot et al., 2002). This software adjusts the spectra to obtain the line width and<br />
the peak positions (in ppm) and to integrate each peak so as to obtain the percentage of each<br />
contribution. The deconvolution and determination of different measurements were performed<br />
as reported previously.<br />
3. Results<br />
3.1. Physico-chemical characterisation of substrates<br />
The physical and chemical characteristics of the three substrates are presented in Table<br />
5. The pH of all the three substrates was between 5.45 and 4.9 for OTL and OMW<br />
respectively. While nitrogen content was the same for all substrates (0.82±0.1 %), the C/N<br />
ratio was different for all substrate and OTL presented the lowest value (54.07±2.12) followed<br />
by OC and OMW (60.11± 3.24 and 63.2±0.65 respectively). Statistical analysis (P=0.05)<br />
showed no significant difference in carbohydrate and lignin content were observed for OMW<br />
and OC but significant difference for OTL which presented the highest carbohydrate content<br />
(68.64±2.68 %) and lowest lignin content (16.87±1.86 %).<br />
97
Résultats et discussion. Chapitre 3: Evaluation des facteurs de croissance<br />
Table 5. Chemical composition of olive twigs and leaves (OTL), olive cake (OC) and<br />
olive mill wastewater (OMW) used as substrate in different culture media for L.edodes<br />
growth.<br />
Physicochemical<br />
properties<br />
Substrate<br />
OTL OC OMW (dry matter)<br />
Moisture content (%) 10.0±0.41 10±0.35 90±1.2<br />
pH 5.45±0.15 5.1±0.2 4.9±0.4<br />
C (%) 42.72±2.01 51.12±0.18 41.6±1.5<br />
N (%) 0.81±0.15 0.85±0.09 0.8±0.01<br />
C/N 54.07±2.12 60.11± 3.24 63.2±0.65<br />
Carbohydrate (%) 68.64±2.68 53.00±2.32 49.72±1.62<br />
Lignin (%) 16.87±1.86 25.81±1.31 22.25±3.65<br />
Carbohydrate:lignin ratio 4.07±0.02 2.05±0.01 2.23±0.35<br />
Fat (%) - 10±1.20 10±2.21<br />
3.2. Formulation of solid olive substrate<br />
Lentinula edodes strains Le119, selected from previous screening experiments in<br />
respect to their efficacy in decolorizing OMW (Lakhtar et al., 2009), was initially grown on<br />
PDA supplemented with 10% of OMW. The effect of various olive substrate mixtures on<br />
mycelial biomass of L. edodes Le119 strain is shown in Table 6. Treatments two and three,<br />
containing a mixture of OTL and OC, had mycelial biomass of 16.24 and 14.58 mg.g -1 of<br />
substrate dry weight (dw), respectively. The treatment containing OTL wetted with OMW<br />
were considered group d containing the lowest mycelial yield (5.98 mg.g -1 of substrate dw).<br />
The best formulation of SOS contains OTL (20 %), OC (30 %) and OMW (50 %). This<br />
mixture will used to follow this work.<br />
Table 6. Mycelial Biomass of L. edodes (Le119 strain) on different mixtures of substrate.<br />
Trt. No Ingredient Mycelial biomass<br />
OTL (%) Olive cake (%) OMW (%) (mg.g -1 )*<br />
1 0 50 50 9.91 c**<br />
2 20 30 50 16.24 a<br />
3 30 20 50 14.58 b<br />
4 50 0 50 5,98 d<br />
* Biomass produced after 1 month of incubation.<br />
** Means followed by the different letter are significantly different (P= 0.05 after LSD).<br />
98
3.3. Screening design<br />
Résultats et discussion. Chapitre 3: Evaluation des facteurs de croissance<br />
Sixteen experiments have been carried out according to the preparation method<br />
described above and the conditions fixed by the experimental design. Although the main<br />
purpose was to achieve the maximization of mycelial biomass production and the<br />
maximization of total phenol removal. The obtained values related to the mycelia biomass<br />
(mg of mycelium/g of substrate) and total phenol removal (%) are as shown in Table7.<br />
Table 7. Experimental conditions of the screening design with 6 factors (A-F) and the<br />
corresponding experimental responses.<br />
Run<br />
no<br />
A B C D E F Biomass<br />
Yield (mg.g -1 )<br />
Total phenol<br />
removal (%)<br />
1 5 20 25 20 T2 SO 48.72 82.35<br />
2 6 30 10 10 T1 SO 20.30 41.28<br />
3 5 20 25 20 T1 MS1 34.50 61.65<br />
4 6 30 10 10 T2 MS1 16.24 42.00<br />
5 5 20 10 10 T2 MS3 12.18 24.12<br />
6 6 30 25 20 T1 MS3 33.50 51.91<br />
7 5 20 10 10 T1 MS2 29.44 36.23<br />
8 6 30 25 20 T2 MS2 38.56 63.75<br />
9 5 30 25 10 T2 MS4 12.18 15.48<br />
10 6 20 10 20 T1 MS4 22.32 29.49<br />
11 5 30 25 10 T1 MS3 14.20 31.51<br />
12 6 20 10 20 T2 MS3 28.42 47.20<br />
13 5 30 10 20 T2 MS1 34.50 45.45<br />
14 6 20 25 10 T1 MS1 18.26 40.71<br />
15 5 30 10 20 T1 SO 44.66 82.68<br />
16 6 20 25 10 T2 SO 30.44 47.98<br />
3.4. Variable effects on mycelial biomass response<br />
Coefficient values (the weight associated to each factor level) and statistical analyses,<br />
using t-test are reported in Table 8a. These results are illustrated by the two histograms shown<br />
in Fig.1. Fig. 1a represents the weight of each factor taking its highest level as reference. Fig.<br />
1b represents the differential effects of each factor when considering two different levels<br />
taken two by two (Mathieu and Phan-Than-Luu 1997; Lewis et al., 1999).<br />
99
Résultats et discussion. Chapitre 3: Evaluation des facteurs de croissance<br />
From Figure 1 and Table 8a was observed that the significant parameter effects were<br />
mineral solution (F) and inoculum size (D), while the rest of the parameters had no significant<br />
effects. Figure 1a showed that inoculum size exhibited significant positive effects on the<br />
response: the increase of these variables resulted in the increase of mycelial biomass which<br />
was correlated to the total phenol removal. The mineral solution, on the other hand, exhibited<br />
a negative effect on the response. Finally, the use of OMW without supplements showed an<br />
interesting positive effect on the mycelial biomass (Fig. 1a).<br />
3.5. Variable effects on total phenol removal response<br />
Coefficient values and related statistical analyses are reported in Table 8b. These results<br />
are illustrated by the two histograms shown in Figure 2. The significant parameter effects<br />
were found, at all levels studied, as mineral solution (F) and inoculum size (D). Initial pH of<br />
substrate, inoculum age, particle size and aeration seemed to have no significant effects. The<br />
results showed a correlation between the parameters effecting the mycelial biomass and total<br />
phenol removal.<br />
3.6. Substrate composition<br />
Measurements of pH and of different experiments after the colonization process by<br />
L. edodes Le119 were different. A steady decrease of pH in all media without supplement<br />
tested was observed; the initial pH values (5.0–6.0) fell at the end of incubation to 4.39 and<br />
4.66 for experiment 1 and 16 respectively. The initial pH of substrates affected slightly the<br />
final pH after the colonization. On the other hand, the supplemented media showed an<br />
increase of pH after incubation (from 5 to 5.6 for experiment 9).<br />
100
Résultats et discussion. Chapitre 3: Evaluation des facteurs de croissance<br />
Table 8. Estimates and statistics on the coefficients response for each factor.<br />
Factor Coefficient F.Inflation Standard texp. Significance<br />
Name<br />
deviation<br />
(% )<br />
(a)Mycelial Biomass response<br />
Cst 5.31 3.31 1.60 12.3<br />
A1 2.87 1.00 1.83 1.57 13.2<br />
B1 1.37 1.00 1.83 0.75 46.2<br />
C1 2.62 1.00 1.83 1.43 16.7<br />
D1 16.12 1.00 1.83 8.78 < 0.01 ***<br />
E1 0.37 1.00 1.83 0.20 84.0<br />
F1 18.50 2.25 3.18 5.81 < 0.01 ***<br />
F2 8.50 2.25 3.18 2.67 1.93*<br />
F3 4.75 2.25 3.18 1.49 15.0<br />
F4 17.00 1.75 3.67 4.63 0.0130<br />
(b) total phenol removal response<br />
Cst 8,31 3.529 1.57 13.6<br />
A1 3,37 1.00 1.957 1.15 26.8<br />
B1 -1,52 1.00 1.957 -0.52 61.2<br />
C1 4,26 1.00 1.957 1.45 16.6<br />
D1 24,25 1.00 1.957 8.26 < 0.01 ***<br />
E1 -2,48 1.00 1.957 -0.85 41.0<br />
F1 38,66 2.25 3.390 7.60 < 0.01 ***<br />
F2 25,21 2.25 3.390 4.96 0.0143 ***<br />
F3 16,44 2.25 3.390 3.23 0.521 **<br />
F4 28,74 1.75 3.915 4.90 0.0162 ***<br />
101
Résultats et discussion. Chapitre 3: Evaluation des facteurs de croissance<br />
Figure 1. Graphical study of the effects of the factors A-F on the mycelial biomass response,<br />
(a) Graphical study of the total effects, (b) Differences of the weights of the different levels.<br />
102
Résultats et discussion. Chapitre 3: Evaluation des facteurs de croissance<br />
Figure 2. Graphical study of the effects of the factors A-F on total phenol removal, (a)<br />
Graphical study of the total effects, (b) Differences of the weights of the different levels.<br />
4. Discussion<br />
The present study was aimed at the screening 6 factors of L. edodes growth on OTL<br />
and OC supplemented with OMW. A pure culture of L. edodes, which was previously<br />
selected for its capability to remove phenolic compounds from OMW (Lakhtar et al., 2009),<br />
was employed to both convert the substrate into mycelial biomass and remediate the<br />
phytotoxic effects of OMW through the growth of L. edodes mycelium.<br />
Mycelial biomass revealed that the substrate formulation exercised a considerable<br />
influence on the mycelium production and therefore on morphologic aspect of mycelium. The<br />
fact that the treatment 1, 2 and 4 (9.91, 16.24 and 5.98 mg.g-1 respectively) supported<br />
different yield biomass indicates that mycelium growth is related to the composition and<br />
structure of the substrate. The use of OTL supplemented with 50% OMW was limited for the<br />
bioavailability of carbohydrate which was reported as an important factor at the beginning of<br />
103
Résultats et discussion. Chapitre 3: Evaluation des facteurs de croissance<br />
cultivation. Moyson and Verachtert (1991) have demonstrated that the growth of L.edodes is<br />
initially associated to hemicellulose content of substrate, since hemicellulose and other water-<br />
soluble sugars are mainly utilized during the active growth phase, prior to the breakdown of<br />
lignin and cellulose. On the other hand, the use of OC supplemented with OMW caused in the<br />
oxygen (O2) depletion could be the cause which reduces mycelial biomass development in<br />
substrates (Ohga, 1990). Small air spaces present in OC supplemented may slow gas<br />
exchange from deep within the interior of the substrate to the surface (Royse and Sanchez-<br />
Vazquez 2001).<br />
In many cases, different factorial designs were used in the literature for assessing the<br />
effect of environmental factor on biological responses (Ranz and Lankmayr 2006; Swalley et<br />
al., 2006; Purama and Goyal 2008). The factorial design and statistical analysis were<br />
performed with NemrodW software (Mathieu and Phan-Tan-Luu, 2000). The combination of<br />
moisture content, which was maintained at 50% (v/v) with the incubation of substrate in oven<br />
for two days without forced aeration, allowed preventing any contamination of L. edodes<br />
culture with filamentous fungi which are known as major antagonistic agent in mushroom<br />
culturing (Savoie and Mata, 1999; Terashima et al., 2002; Kalmis and Sargin 2004). The<br />
supply of culture with humidified air allowed gradual increase of the substrate moisture<br />
contentand keeping the substrate with suitable moisture content.<br />
The influence of the composition of the substrate on mushroom production characters<br />
has been emphasized in recent studies (Philippoussis et al., 2001; Sunchez and Royse 2001;<br />
Philippoussis et al., 2003). In the framework of the present study, the nutriment added to the<br />
substrate and the spawn size were the main parameter that affected the growth of L. edodes.<br />
This can be attributed to the nitrogen content, imported with addition of the mineral solution,<br />
which exercised a negative effect on mycelial growth. Indeed, Nitrogen has been recognized<br />
as a growth limiting factor for the L. edodes cultivation (Boyle 1998; Kalberer 2000), while<br />
recently a positive correlation of mycelial growth to low C/N ratio values of substrates used<br />
for the cultivation has been demonstrated as well (Philippoussis et al., 2003; 2007). However,<br />
in L. edodes cultivation on synthetic composite media (OTL + OC + OMW), supplementary<br />
nutritive solution should not be used. Otherwise, when the nutritive solution was added, we<br />
observed that the risk of contamination of the growth media was increased considerably.<br />
Similar findings were reported by Schunemann (1988).<br />
The data obtained on polyphenol removal from the substrate by the culture of<br />
L. edodes, showed that there was a negative effect of mineral solution added to initial<br />
104
Résultats et discussion. Chapitre 3: Evaluation des facteurs de croissance<br />
substrate on polyphenol removal. These results are in accordance with previous findings<br />
demonstrating that high content of N had a negative effect of the degradation of procyanidins<br />
by Aspergillus fumigatus (Contreras-Dominguez, 2006). Our results, demonstrating decrease<br />
of pH in the substrate without supplement after the incubation period, are comparable to those<br />
obtained on oak-wood sawdust substrates (Philippoussis et al., 2003), and they confirm<br />
previous observations stating that a low pH value of the substrate is probably a prerequisite<br />
for primordia formation (Kalberer 1995).<br />
Spawn inoculation size was found an important factor that affects the mycelial growth.<br />
Compared to the straw, spawn is a relatively expensive item. Using the lowest inoculation<br />
size possible without sacrificing the mushroom yield would give the best economics. The<br />
spawn size is defined as the weight ratio of spawn to substrate on a fresh weight basis. The<br />
optimum spawn size depends on the type of substrate, mushroom species, spawn quality, and<br />
the cultivation conditions. Among the two spawn size (10 and 20%) tested, the 10% spawn<br />
size resulted in significantly lower mycelial biomass than the other size. The mycelial<br />
biomass at the 10% size was 37.5% lower than at the 20% size for the substrate without<br />
supplement. The mycelial growth at the 10% size showed contamination by green and black<br />
moulds during cultivation of the L.edodes Le119 strain. Therefore, based on these results, it<br />
would be recommended that at least 10% spawn level be used to achieve a colonization of the<br />
substrate by L. edodes without any mould contamination.<br />
5. Conclusion<br />
The growth of L. edodes Le119 on SOS using different mixture allowed us to<br />
determine the optimal formulation which was OTL (20%) + OC (30%) + OMW (50%)<br />
allowing mycelial biomass production of 16,24 mg.g -1 of substrate dry weight. The nutriments<br />
added to the substrate which negative effect and spawn size which positive effect were the<br />
main parameter that affected the growth of L. edodes Le119 on optimal SOS. Thus, the<br />
optimal condition of growth of L. edodes Le119 on SOS are the following; SOS composition<br />
(20% of OTL, 30% of OC and 50% of OMW), initial pH of 5, temperature of incubation<br />
25°C, aeration flow 20 mL.min -1 , inoculum age of 10 days, inoculum size 10%, 70% of<br />
particle size between 2 and 0.8 mm mixed with 30% of particle size less than 0.8 mm and<br />
with addition of nutrients.<br />
105
Résultats et discussion. Chapitre 4: Transformation de la matière organique<br />
Chapitre IV<br />
Transformation de la matière organique et détoxification des<br />
substrats oléicoles solides par la culture de Lentinula edodes par<br />
fermentation en milieu solide<br />
Dans le chapitre précédent, nous avons évalué l’influence de six facteurs de croissance<br />
de L. edodes Le119 sur le SOS en fermentation en milieu solide. Les résultats ont montré que<br />
l’ajout de la solution minérale en concentration élevée limite la croissance, tandis que le taux<br />
d’inoculum exerce un effet positif sur la croissance. Pour étudier la cinétique de croissance de<br />
L. edodes Le119 sur le SOS, on a choisi les conditions optimales de croissance qui sont les<br />
suivantes : pH initial 5, température d’incubation 25°C, débit d’aération 20 mL.min -1 , âge<br />
d’inoculum 10 jours, taille d’inoculum 10%, particules avec une taille entre 2 et 0,8 mm à<br />
raison de 70% et des particules avec une taille inferieure de 0,8 à raison de 30% , sans<br />
addition de la solution minérale et l’humidité du substrat a été ajustée à 50% avec les<br />
margines.<br />
Dans la littérature, de nombreuses études ont été menées sur la physiologie de<br />
croissance des champignons filamenteux en fermentation en milieu solide (Raimbault, 1980 ;<br />
Roussos, 1985 ; Brand, 2006 ; Hassouni, 2007), tandis que l’information sur la physiologie<br />
des basidiomycètes de pourriture blanche est peu nombreuse et en particulier celle de<br />
L. edodes cultivée sur des substrats oléicoles reste toujours rare (D’Annibale et al., 2004,<br />
Tomati et al., 2004). Par conséquent, il est apparu intéressant de mettre l’accent sur la<br />
physiologie de croissance et la respirométrie de L. edodes sur le SOS qui regroupe l’ensemble<br />
des substrats oléicoles: bois de taille, grignons d’olive et margines.<br />
106
Résultats et discussion. Chapitre 4: Transformation de la matière organique<br />
Dans ce nouveau chapitre sont donc présentés les résultats de l’étude de l’évolution<br />
des paramètres physico-chimiques et biologiques au cours de la croissance de L. edodes<br />
Le119 sur le SOS en fermentation en milieu solide. Il fait état des relations établies entre des<br />
paramètres classiques tels que biomasse mycélienne, sucres réducteurs, phénols totaux, teneur<br />
en carbone, teneur en azote, rapport C/N, les activités enzymatiques telles que l’activité<br />
cellulase, laccase, Mn-peroxydase, Lignine peroxydase, lipase et les caractéristiques du SOS<br />
fermenté déterminés par spectroscopie UV-visible et Résonance Magnétique Nucléaire 13 C du<br />
solide.<br />
107
1. Introduction<br />
Résultats et discussion. Chapitre 4: Transformation de la matière organique<br />
L’oléiculture représente une des plus anciennes activités agricoles dans le bassin<br />
méditerranéen. Toutefois, elle présente l’inconvénient de générer d’énormes quantités de<br />
déchets liquides dont la fraction organique complexe leur confère un pouvoir fortement<br />
polluant (Fiorentino et al., 2003). A côté des margines, l’industrie oléicole produit deux co-<br />
produits solides, les grignons d’olive et le bois de taille.<br />
Nombreuses études ont été menées en vue de trouver des solutions pour le traitement<br />
des déchets oléicoles, en particulier la réduction de la charge phénolique des margines par la<br />
culture de champignons. Les perspectives de ces études ont mis l’accent sur la valorisation<br />
des déchets de l’oléiculture par la culture des basidiomycètes en fermentation en milieu solide<br />
(FMS). Les basidiomycètes de pourriture blanche, en particulier Lentinula edodes (Berk.)<br />
Pegler, sont capables de se développer en présence de la lignine, et leur développement est<br />
associé à la production des enzymes phénoloxydases non spécifiques. Pour cette raison, les<br />
basidiomycètes ont été considérés comme de bons candidats pour permettre à la fois une<br />
valorisation des substrats oléicoles lignocellulosiques et à la réduction des polyphénols des<br />
margines (Zervakis et al., 1996, Kalmis et Sargin, 2009).<br />
L’objectif de l’étude est d’évaluer la transformation de la composition chimique et la<br />
réduction en polyphénols du mélange de substrats oléicoles solide (SOS) constitués de bois de<br />
taille, grignons d’olive et margines durant la croissance du mycélium de L. edodes Le119.<br />
Une attention particulière a été consacrée à l’évaluation de la production de CO2 durant la<br />
croissance mycélienne de la souche Le119 au changement de l'abondance relative des<br />
fractions de carbone, à la fois simples et complexes analysés par la technique de la CPMAS<br />
RMN 13 C soutenue par les analyses de routine, pour relier l’activité métabolique du<br />
champignon à la transformation des résidus oléicoles .<br />
2. Matériel et méthodes<br />
2.1. Origine des substrats<br />
Les margines d’olive, ainsi que les grignons d’olive, ont été prélevés dans une unité de<br />
trituration des olives avec le procédé triphasique fonctionnant avec le système de presse. Les<br />
standards phénoliques ont été achetés auprès d’Extrasynthèse (France). Tous les solvants<br />
utilisés pour l’extraction étaient de qualité HPLC (Sigma, France).<br />
108
Résultats et discussion. Chapitre 4: Transformation de la matière organique<br />
2.2. Microorganisme et préparation du spawn<br />
Une souche de L. edodes Le119 a été préalablement sélectionnée pour sa capacité de<br />
dégradation des polyphénols des margines (Lakhtar et al., 2009). La souche a été<br />
régulièrement repiquée sur un milieu PDA contenant 10% des margines. Le blanc spawn a été<br />
préparé sur les gains de blé contenant 10% de margines (Chapitre 3). La préparation du spawn<br />
a été effectuée dans les colonnes de Raimbault et l’incubation a été faite à 25°C pendant 10<br />
jours.<br />
2.3. Préparation des substrats SOS<br />
Le substrat a été préparé de la même façon précitée (Chapitre 4). Les grignons d’olive ont été<br />
homogénéisés mécaniquement, à l’aide d’un mixeur, tandis que le bois de taille a été broyé et<br />
tamisé à une taille de particule comprise entre 2 mm et 0,8 mm. Les particules ayant avec une<br />
taille inférieure à 0,8 mm ont aussi été utilisées dans la composition du substrat représentant<br />
30% du poids total du bois de taille. Les substrats ont été mélangés avec les margines et<br />
chauffés pour assurer l’absorption totale de margines. Le mélange (SOS) a été stérilisé à<br />
121°C pendant 30 min.<br />
2.4. Inoculation sur mélange composite pour L. edodes<br />
L’inoculation a été faite à raison de 10% (poids humide : poids humide), et le remplissage des<br />
colonnes a été fait en alternant des couches de substrat et de spawn, pour assurer une<br />
croissance homogène du mycélium. Les colonnes ont été incubées dans l’étuve sans aération<br />
forcée à 25°C pendant 2 jours. Elles ont été, par la suite, placées dans le dispositif de la FMS<br />
pour une durée d’incubation d’un mois. Deux colonnes témoins non inoculées ont été<br />
préparées et incubées dans les mêmes conditions.<br />
2.5. Montage du système de FMS<br />
Les colonnes ont été placées dans un bac rempli d’eau maintenue à 25°C ; l’air traversant le<br />
substrat a été humidifié 3 fois pour arriver à une humidité saturante de l’air. Le débit d’air a<br />
été réglé avec des microvannes et régulièrement ajusté à 20 mL.min -1 . La Figure 4 montre le<br />
fonctionnement du système. La fermentation a été suivie pendant 30 jours.<br />
109
Résultats et discussion. Chapitre 4: Transformation de la matière organique<br />
Figure 1. Dispositif de FMS connecté au Système de Respirométrie PNEO.<br />
2.6. Analyse des échantillons<br />
L’activité respiratoire a été mesurée trois fois par jour à l’aide du PNEO, qui permet de<br />
mesurer à la fois quatre paramètres ; le débit d’aération, l’humidité de l’air, la température de<br />
l’air et la teneur de l’air en CO2 à la sortie de la colonne. La biomasse fongique a été estimée<br />
par la quantification de l’ergostérol (Chapitre 3). Les sucres réducteurs ont été analysés par la<br />
méthode de Miller (1959) en utilisant le réactif de DNS et les sucres totaux ont été déterminés<br />
par la méthode de DuBois et al. (2002) en utilisant l’anthrone. La fraction lipidique a été<br />
extraite dans l’hexane à l’aide de l’appareil de Soxhlet (Cordova et al., 1998). L’extraction de<br />
la fraction phénolique a été réalisée dans une solution eau-méthanol (40-60); à un gramme du<br />
substrat fermenté lyophilisé et finement broyé, on a ajouté 10 mL de la solution organique. Le<br />
mélange a été homogénéisé et incubé pendant 1h à 4°C. L’extraction a été répétée trois fois.<br />
La quantification des phénols totaux a été faite suivant la méthode de Folin Ciocalteu<br />
(Bärlochar et Garça, 2005) en utilisant l’acide cafféique comme standard (250 µg.mL -1 ).<br />
Selon Chin et al., (1994), l’absorbance à 280 nm est une mesure approximative du degré<br />
d’aromaticité des matières organiques dissoutes.<br />
110
Résultats et discussion. Chapitre 4: Transformation de la matière organique<br />
2.7. Analyses chromatographiques<br />
L’extraction des monomères a été effectuée en deux étapes. La première étape consiste à<br />
l’extraction de la fraction phénolique par une solution eau-méthanol (40-60). La deuxième<br />
étape consiste en la purification des monomères phénoliques par l’extraction avec l’acétate<br />
d’éthyle suivant la méthode modifiée de De Marco et al. (2007). L’analyse des monomères a<br />
été réalisée par l’HPLC suivant la méthode modifiée de De Marco (2007). La plateforme<br />
HPLC est composée d’un dégazeur (1200 Series Degasser, Agilent technologies), d’une<br />
pompe quaternaire (1200 Series Quaternary Pump, Agilent technologies), d’un<br />
échantillonneur liquide automatique ou passeur (1200 Series Automatic Liquid Sampler,<br />
Agilent technologies) et d’un détecteur à barrette de diodes (1200 Series Diode-Array<br />
Detector, Agilent technologies). La séparation des composés a été opérée sur la colonne<br />
Atlantis d18 (5µm, 4,6 x 250 mm) Waters gardée à une température constante (température<br />
ambiante de 23±2°C). La détection des composés a été effectuée à 280 nm. Les pics ont été<br />
identifiés par comparaison avec les standards.<br />
2.8. Activités enzymatiques<br />
2.8.1. Extraction d’enzymes<br />
L’extraction des enzymes a été effectuée sur 3 g du substrat fermenté frais, décongelé,<br />
mélangé avec 10 mL de tampon acétate (0,1M, pH 5,0) contenant 5 mM de CaCl2, 0,05% du<br />
Tween 80 et 3% du polyvinylpolypyrrolidone insoluble (PVPP) (Sampedro et al., 2007). Le<br />
mélange a été incubé à la température ambiante pendant 1h et l’extrait aqueux a été récupéré<br />
par centrifugation (4500 rpm pendant 10 min) suivie d’une filtration sur papier Watman N°2.<br />
2.8.2. Activité enzymatique<br />
L’activité endoglucanase (cellulase, EC 3.2.1.4) a été déterminée selon la méthode de<br />
Mandels et al. (1976). L’activité enzymatique est exprimée en unité par g du substrat poids<br />
sec (SPS) (1U= 1 µmole de sucres réducteurs libérés par min). L’activité lipase a été<br />
déterminée suivant la technique rapportée par Goujard et al. (2009), en utilisant le p- laurate<br />
de nitrophényl comme substrat. L’activité tannase a été mesurée suivant la technique de<br />
Sharma et al. (2000). Les activités de Mn peroxydase, lignine peroxydase et laccase ont été<br />
mesurées suivant la technique rapportée par Lakhtar et al. (2009). La technique décrite par<br />
Santos et Linardi (2004) a été utilisée pour la détermination des activités catéchol-1,2-<br />
dioxygénase et protocatéchuate-3,4-dioxygénase. L’activité est exprimée en unité par g SPS<br />
(1U= 1 µmole de produit formé par min).<br />
111
Résultats et discussion. Chapitre 4: Transformation de la matière organique<br />
2.9. Analyses élémentaires : C et N<br />
Les teneurs en C et N, dans les échantillons de culture de L. edodes, ont été déterminées à<br />
l’aide d’un analyseur élémentaire CHN (Flash EA 1212 Elemental Analyzer, France).<br />
L’analyseur CHN a été calibré avec l’acide aspartique à différentes concentrations.<br />
2.10. Résonance Magnétique Nucléaire 13 C du solide (13 CP/MAS RMN)<br />
Les spectres haute résolution ont été réalisés sur le spectromètre Bruker DSX 400 kHz<br />
du Spectropole (Université Paul Cézanne). Les spectres 13 C CP/MAS ont été enregistrés à la<br />
fréquence de 100,7 MHz avec découplage du proton. Environ 300 mg de l’échantillon ont été<br />
placés dans une sonde de 7 mm inclinée à l’angle magique (angle de 54,7° par rapport au<br />
champ magnétique statique) tournant à 6 kHz. Les paramètres d’acquisition utilisés pour un<br />
enregistrement optimal sont une impulsion de 90° pour le proton de durée 2,8 µs, un délai de<br />
répétition de 3 s, un nombre d’acquisition de 2000, et enfin un temps de contact de 2 ms. La<br />
déconvolution des spectres a été effectuée grâce au logiciel DmFit (Massiot et al., 2002).<br />
3. Résultats et discussions<br />
3.1. Croissance de L. edodes sur SOS<br />
Le remplissage des colonnes de Raimbault avec le substrat SOS prétraité a été alterné<br />
avec l’ajout du spawn, qui permet un envahissement homogène et uniforme du substrat par le<br />
mycélium. L’incubation des colonnes pendant deux jours dans l’étuve à 25°C nous a permis<br />
d’éviter les contaminations par les moisissures. Après deux jours d’incubation, on a remarqué<br />
que tous les grains du spawn inoculés ont démarré par le développement du mycélium.<br />
L’envahissement total du substrat a été remarqué après 24 jours d’incubation. Par ailleurs,<br />
aucune contamination n’a été observée sur les deux colonnes témoins non inoculées.<br />
3.2. Analyses des effluents gazeux par PNEO<br />
A la sortie des colonnes de Raimbault , les effluents gazeux ont été collectés afin de<br />
suivre l’évolution de quatre paramètres (débit d’air, humidité relative, température et la<br />
production de CO2) en fonction du temps (Fig. 1). L’humidité relative et la température de<br />
l’air à la sortie des colonnes fluctuent entre deux valeurs (50 et 58 %) pour la l’humidité et<br />
(23 et 27°C) pour la température de l’air. Avant la mesure de CO2 à la sortie de la colonne, le<br />
débit de l’air traversant la colonne, a été ajusté à 20 mL/min.<br />
112
Humidité<br />
(%)<br />
Température<br />
(°C)<br />
Résultats et discussion. Chapitre 4: Transformation de la matière organique<br />
Figure 2. Evolution de trois paramètres; ( ) CO2 produit, ( ) température de l’air, ( )<br />
humidité relative de l’air, au cours de 30 jours de culture de L. edodes sur SOS à 25°C.<br />
Sur la figure 2, on remarque que l’évolution de CO2 suit quatre étapes alternées. Tout<br />
d’abord, durant les 6 premiers jours d’incubation sur le dispositif FMS, une augmentation<br />
continue de CO2 a été remarquée passant de 0 à 0,23%. Entre le 6 ème et 8 ème jour d’incubation,<br />
on observe un premier plateau de stabilisation situé à 0,23% de CO2 produit. Par contre, entre<br />
le 8 ème et le 14 ème jour d’incubation, une augmentation importante de la respiration du<br />
mycélium de Le119 a été remarquée allant de 0,23 à 0,7. Un deuxième plateau de stabilisation<br />
de CO2 a été observé entre le 14 ème et 16 ème jour de FMS. Enfin, à partir du 23 ème jour de FMS<br />
la respiration a été maximum et constante pendant une semaine avec un taux élevé de CO2<br />
produit (1,18%)<br />
Temps (jour)<br />
3.3. Evolution de la biomasse de L. edodes sur SOS<br />
L’estimation de la biomasse a été déterminée indirectement par la quantification de<br />
l’ergostérol contenu dans le substrat fermenté par L. edodes Le119. Le coefficient de<br />
conversion de l’ergostérol en biomasse de L. edodes a été évalué à 3,57 mg d’ergostérol pour<br />
1 g de biomasse pure (Chapitre 3). L’évolution de la biomasse au cours de la FMS (Fig.3) suit<br />
relativement l’allure de production de CO2 mesuré dans les effluents gazeux (Fig.2).<br />
CO2 (%)<br />
113
Biomasse (mg.g -1 SPS)<br />
Résultats et discussion. Chapitre 4: Transformation de la matière organique<br />
Figure 3. Evolution de la biomasse de la souche Le119 produite au cours de 30 jours<br />
d’incubation sur SOS à 25°C.<br />
L’évolution de la biomasse au cours de la FMS passe par trois phases de croissance<br />
rapide alternant avec deux phases de croissance relativement lente. La biomasse évolue de<br />
2.47 à 5 mg/g SPS du SOS pendant les 6 premiers jours d’incubation. Ensuite entre le 6 ème et<br />
le 12 ème jour d’incubation, une augmentation importante de la biomasse produite a été<br />
remarquée, allant de 5 à 11,4 mg.g -1 SPS du SOS. Par contre entre le 12 et 18 ème jour<br />
d’incubation, la biomasse passe de 11,4 à 14,5 mg.g -1 SPS. Cette évolution de biomasse<br />
correspond au deuxième plateau de CO2 produit durant le développement du mycélium<br />
(Fig.2). Entre 18 et 24 jours d’incubation, une augmentation importante a été de nouveau<br />
observée, allant de 14,5 à 20,5 mg.g -1 SPS du SOS. Cette augmentation correspond également<br />
à l’augmentation de CO2 observée dans la Figure 2. Enfin, un plateau de biomasse a été<br />
obtenu à partir de 24 jours de FMS pour arriver à une biomasse de 21,61 mg.g -1 après 30 jours<br />
d’incubation. Après un mois d’incubation, la biomasse estimée dans notre étude dépasse 2,5<br />
fois la biomasse du Pleurotus ostreatus produite sur les coques de café avec le même taux<br />
d’inoculation (10%) (Sobal, 2002).<br />
Temps (jour)<br />
114
Résultats et discussion. Chapitre 4: Transformation de la matière organique<br />
3.4. Evolution des sucres réducteurs, sucres totaux, pH du substrat fermenté<br />
L’évolution de sucres réducteurs, des sucres totaux et du pH du substrat inoculé avec la<br />
souche Le119 de L. edodes est présentée dans la Figure 4.<br />
Sucres mg.g -1 SPS<br />
Figure 4. Evolution des sucres réducteurs ( ), sucres totaux ( ) et du pH ( ) du substrat<br />
fermenté au cours de 30 jours de culture de L. edodes Le119 sur SOS à 25°C.<br />
On remarque une diminution de la teneur en sucres réducteurs et totaux dans le milieu<br />
après 12 jours d’incubation, suivie d’une augmentation graduelle pour arriver à 108 et 120<br />
mg.g -1 SPS du SOS respectivement. Accompagné de cette évolution, on remarque que les<br />
valeurs du pH du substrat fermenté restent très stables tout au long de la FMS de la souche de<br />
L. edodes sur SOS et fluctuent de 4,5 à 5.<br />
3.5. Evolution des phénols totaux<br />
Les phénols totaux ont été déterminés par la méthode standard de Folin Ciocalteu pour<br />
pouvoir comparer nos résultats à ceux de la littérature. Selon Chin et al., (1994), l’absorbance<br />
à 280 nm est une mesure approximative du degré d’aromaticité des matières organiques<br />
dissoutes.<br />
180<br />
160<br />
140<br />
120<br />
100<br />
80<br />
60<br />
40<br />
20<br />
0<br />
0 5 10 15 20 25<br />
0<br />
30<br />
Temps (jour)<br />
5<br />
4<br />
3<br />
2<br />
1<br />
pH<br />
115
mg.g -1 SPS du substrat fermenté<br />
45<br />
40<br />
35<br />
30<br />
25<br />
20<br />
15<br />
10<br />
5<br />
0<br />
Résultats et discussion. Chapitre 4: Transformation de la matière organique<br />
0 6 12 18 24 30<br />
Temps (jour)<br />
Figure 5. Evolution de la décoloration de l’extrait phénolique ( ) et des phénols totaux ( ) du<br />
substrat fermenté au cours de 30 jours de culture de L. edodes Le119 sur SOS à 25°C.<br />
Au cours de la culture de L. edodes Le119 sur SOS en FMS pendant 30 jours, une<br />
forte corrélation entre la dégradation des phénols et la décoloration du substrat a été observée.<br />
Le développement du champignon a permis une dégradation de 80% des phénols totaux et<br />
85% de décoloration du substrat (Fig. 5). Les spectres UV-vis des extraits d’échantillons<br />
prélevés durant la croissance du champignon sont présentés dans la Figure 6. Les résultats<br />
confirment ceux obtenus à partir de la décoloration et montrent l’apparition de molécules<br />
absorbant aux alentours de 500 nm. Ces molécules peuvent être responsables de<br />
l’augmentation légère des phénols totaux observée après 24 jours d’incubation. Sampedro et<br />
al. (2007) ont mesuré de semblables chutes de teneurs en phénol totaux des grignons humides,<br />
issues du système de trituration à deux phases, par la culture de Paecilomyces farinosus après<br />
20 semaines d’incubation. Selon Aloui et al. (2007), l’ajout de la bagasse de canne de sucre<br />
aux grignons humides favorise la dégradation des phénols totaux. En effet une réduction des<br />
phénols totaux de 60% a été observée après l’ajout de 30% de la bagasse de canne à sucre<br />
pour la culture de Phanerochaete chrysosporium après 6 jours d’incubation.<br />
1,6<br />
1,4<br />
1,2<br />
1<br />
0,8<br />
0,6<br />
0,4<br />
0,2<br />
0<br />
Absorbance à 280 nm<br />
116
Résultats et discussion. Chapitre 4: Transformation de la matière organique<br />
Figure 6 : Spectres UV-visible de la fraction phénolique extraite des échantillons du substrat<br />
fermenté au cours de 30 jours de culture de la souche Le119 sur SOS à 25°C.<br />
3.6. Les monomères phénoliques<br />
La disparition des monomères phénoliques, extraits du SOS fermenté par L. edodes<br />
Le119, est présentée dans le Tableau 1.<br />
Tableau 1. Concentration des monomères phénoliques (µmole.g -1 SPS du SOS) durant la<br />
culture de L. edodes pendant 1 mois d’incubation à 25°C.<br />
Temps Oleuropéine Hydroxy- Acide Tyrosol Acide trans- acide p-<br />
(jours)<br />
Absorbance<br />
tyrosol<br />
caféique<br />
cinnamique <br />
coumarique<br />
0 6,54 4,32 3,22 2,32 2,32 4,51<br />
6 4,21 2,35 3,21 2,12 2,23 3,54<br />
12 3,21 3,21 3,15 1,63 2,35 4,16<br />
18 2,54 2,32 2,36 1,33 2,16 5,54<br />
24 1,02 0,32 1,06 1,03 2,03 3,21<br />
30 0,95 0,32 1,01 0,03 1.97 3,32<br />
Témoin 5.98 4.45 3.11 2.22 2.25 4.02<br />
L’écart type a été estimé à 5%.<br />
Longueur d’onde (nm)<br />
Six monomères ont été identifiés dans l’extrait éthylique obtenu à partir du substrat<br />
fermenté. Leur quantification, déterminée par HPLC, a montré que l’Oleuropéine est le<br />
composé le plus abondant dans le SOS avant la fermentation (6,54 µmoles.g -1 SPS du SOS),<br />
suivie par l’acide p-coumarique et l’hydroxytyrosol (4,51 et 4,32 µmoles.g -1 SPS du SOS<br />
117
Résultats et discussion. Chapitre 4: Transformation de la matière organique<br />
respectivement). Apres un mois de culture de la souche Le119, à l’exception de l’acide trans-<br />
cinnamique qui présente une faible diminution de sa teneur (15 %), tous les composés<br />
analysés ont subi une réduction de leur teneur avec des rendements variables (92 et 26 % de<br />
réduction pour l’hydroxytyrosol et l’acide p-coumarique respectivement). Dans le SOS<br />
témoin, non inoculé et incubé pendant 30 jours avec un débit d’aération de 20 mL.min -1 dans<br />
les mêmes conditions que celles de la culture, on a remarqué une réduction partielle de tous<br />
les composés analysés,. Leur réduction peut être expliquée par l’oxydation non enzymatique<br />
qui pourrait être causée par le passage forcé de l’air dans le substrat durant son incubation<br />
dans le dispositif de la FMS. El Hajjouji et al. (2008) ont reporté qu’une augmentation de<br />
réduction des composés phénoliques est favorisée avec l’application d’une aération forcée sur<br />
les margines.<br />
3.7. Evolution de la composition élémentaire du substrat fermenté<br />
L’évolution de cinq paramètres conventionnels (humidité du substrat, C, N, C/N, Perte<br />
en matière) a été suivie à partir des prélèvements du substrat fermenté au cours de 30 jours<br />
d’incubation à 25°C. Les résultats sont présentés dans le Tableau 2.<br />
Tableau 2: Evolution de différents paramètres chimiques mesurés dans des échantillons du<br />
substrat fermenté au cours de 30 jours de culture de L. edodes sur SOS à 25°C.<br />
Temps<br />
(jours)<br />
C (%) N (%) C/N Humidité<br />
(%)<br />
Perte en<br />
Matière (%)<br />
0 48.52±1,3 0,95±0,10 51,07 44.30±0.49 -<br />
6 46.17±0,7 0.95±0,06 48,60 45.14±2.61 1.87±0.67<br />
12 43.89±1,4 0,96±0,04 45,72 45.94±1.14 4.99±1.01<br />
18 42.89±1,7 1,09±0,07 39,35 46.47±0.92 6.73±1.80<br />
24 41.32±1,2 1,31±0,08 31,54 48.39±0.37 9.48±1.59<br />
30 40.32±0,6 1,52±0,10 26,53 46.41±1.07 12.18±1.39<br />
Un test d’analyse de variance a été effectué pour l’ensemble des paramètres étudiés.<br />
Les résultats montrent une différence significative entre les prélèvements (p= 0.05). Le<br />
passage de l’air humidifié à travers les colonnes (volume utile de 200 mL) avec un débit de<br />
20 mL.min -1 a permis de maintenir une humidité de 46±2 %. Les résultats des analyses<br />
élémentaires, montrent que le carbone diminue en passant de 48.52 à 40,32 % pendant la<br />
durée d’incubation, alors que l’azote présente une augmentation de 0.95 à 1.52% de la matière<br />
118
Résultats et discussion. Chapitre 4: Transformation de la matière organique<br />
sèche. La diminution de carbone est caractéristique de la dégradation des matières organiques.<br />
En effet, de semblables profils de diminution de carbone ont été rapportés pour la dégradation<br />
des matières organiques (Mena et al., 2003, Huang et al., 2004). Le ralentissement de la<br />
diminution de la teneur en carbone peut être expliqué par la nature lignocellulosique du<br />
substrat qui est dégradé beaucoup plus lentement. Par ailleurs, l’augmentation d’azote est due<br />
à sa concentration engendrée par la dégradation des composés carbonés réduisant la masse<br />
totale du substrat durant la culture. De ce fait, le rapport C/N diminue au cours de la culture<br />
de 51,07 à 26.53. Cette diminution reflète le développement du champignon accompagné<br />
d’une part avec un dégagement de CO2 et, d’autre part, la décomposition de la matière<br />
organique, ainsi que l’enrichissement du substrat en matières azotées.<br />
3.8. Caractérisation des échantillons du SOS par RMN<br />
La caractérisation de la structure carbonée des prélèvements du substrat fermenté a été<br />
effectuée par RMN dans le but d’approfondir la connaissance du changement de la matière<br />
organique durant la croissance du champignon. Les spectres 13 C-RMN (CPMAS) de<br />
l’évolution du SOS durant la croissance de L. edodes Le119 sont présentés dans la Figure 7.<br />
0 jours 6 jours 12 jours 18 jours 24 jours 30 jours<br />
Figure 7 : Spectres RMN solide du 13 C des échantillons du substrat fermenté au cours de 30<br />
jours de culture de L. edodes sur SOS à 25°C.<br />
La déconvolution des 6 spectres RMN du substrat fermenté, effectuée à l’aide du<br />
logiciel DmFit (Massiot et al., 2002), a généré 21 pics correspondant à la distribution des<br />
119
Résultats et discussion. Chapitre 4: Transformation de la matière organique<br />
atomes de carbon dans les différentes structures chimiques. L’intégration de ces pics a permis<br />
de calculer la contribution de chaque type de carbone. Les résonances exprimées en<br />
déplacements chimiques (ppm) ont été assignées à des structures chimiques grâce à plusieurs<br />
études sur la caractérisation des matières organiques (Ait Baddi et al., 2004; Albrecht et al.,<br />
2008; Amir et al., 2004; Ait Baddi et al., 2003; El Hajjouji et al., 2008; Gilardi et al., 1995).<br />
Ainsi, les déplacements chimiques de RMN sont attribués à des groupes chimiques : C-alkyle<br />
(0-45 ppm), C-O-alkyle (45-110 ppm), C-aromatique (110-145 ppm), C-phénolique (145-165<br />
ppm) et C-carbonyle & carboxyle (165-210 ppm). Les résultats sont présentés dans le<br />
Tableau 3.<br />
Tableau 3 : Valeurs des groupements carbonés à partir des intégrations des 21 spectres RMN<br />
du substrat SOS fermenté avec L. edodes Le119, durant un mois d’incubation à 25°C.<br />
Temps<br />
(jour)<br />
C Alkyle<br />
0-45<br />
C O-Alkyl<br />
45-110<br />
C Aromatique<br />
110-145<br />
CPhénolique<br />
145-165<br />
CCarbonyle & Carboxyle<br />
165-210<br />
0 15,67 65,09 8,68 2,22 8,31<br />
6 15,55 64,95 8,89 1,77 8,82<br />
12 17,75 61,96 8,48 1,95 9,83<br />
18 16,89 62,54 8,44 2,06 10,05<br />
24 16,08 63,41 8,34 2,33 9,82<br />
30 16,32 62,94 8,29 2,18 10,25<br />
Des différences significatives ont été calculées entre les prélèvements de culture pour<br />
l’ensemble des groupements carbonés (p< 0,05). Cela signifie que touts les groupements ont<br />
subi une transformation durant le développement de la souche de L. edodes Le119. En effet, à<br />
l’exception du groupement CCarbonyle & Carboxyle qui présente une allure d’augmentation passant<br />
de 8,31 à 10,25% (à l’état initial et après un mois d’incubation respectivement), tous les autres<br />
groupements présentent de périodes d’augmentation et/ou de diminution alternées. Par<br />
exemple, le groupement CPhénolique présente une phase de diminution passant de 2,23 à 1,7%<br />
suivie par une phase d’augmentation allant de 1,77 à 2,33%.<br />
L’examen de déconvolution des spectres montre que le changement du groupement<br />
des CAlkyle est associé d’une part à la diminution des deux pics à 30 et 40 ppm assignés aux<br />
CH2 des protéines et des lipides (Ait Baddi et al., 2004 ; Albreich et al., 2007) et, d’autre part,<br />
à l’augmentation du pic 22 ppm correspondant au méthyl des groupement acétyl lié à la<br />
dégradation de la fraction aromatique (Giraldi et al., 1997) et des hémicelluloses (Wikberg et<br />
120
Résultats et discussion. Chapitre 4: Transformation de la matière organique<br />
Maunu, 2004). La première phase de diminution des CAlkyle semble indiquer que le<br />
champignon utilise, dans la première semaine de sa croissance, préférentiellement les<br />
constituants les plus facilement métabolisables, tandis que la deuxième phase d’augmentation<br />
semble être liée à la génération des produits de dégradation de la fraction aromatique. La<br />
première phase est perçue à travers l’augmentation du taux des CAromatique, notamment<br />
assignés à la lignine, alors que la deuxième phase, est perçue à travers la diminution du<br />
groupement CAromatique. Par ailleurs, les CO-alkyle sont classiquement assignés aux<br />
polysaccharides, car le signal combiné de la cellulose et des hémicelluloses écrase celui des<br />
carbones aliphatiques de la lignine selon Haw et al. (1984) et Wikberg & Maunu (2004). Les<br />
données des spectres, montrent la diminution de ce groupement qui se répercute sur<br />
l’augmentation des sucres réducteurs passant de 68 à 108 mg/g SPS du SOS, du 12 ème au 30 ème<br />
jour d’incubation de culture de la souche de L. edodes Le119.<br />
Tableau 4 : Evolution au cours de culture de l’indice d’aromaticité, de l’estimation du contenu<br />
en lignine, du rapport Syringyl sur Guaiacyl (S/G), du rapport 89/84 (ppm) et du degré de<br />
cristallinité, et de l’estimation du contenu en hémicellulose calculés à partir des pics de RMN.<br />
Temps Poly- Lignine Aromaticité Cristallinité S/G 89/84 C1<br />
jours<br />
saccharide<br />
Hémicellulose<br />
0 57,77 18,01 11,90 0,07 1,81 0,07 6,29<br />
6 57,87 17,60 11,70 0,05 1,87 0,05 6,25<br />
12 54,98 17,23 11,58 0,04 1,56 0,03 5,41<br />
18 55,41 17,34 11,68 0,08 1,51 0,08 5,10<br />
24 56,24 17,61 11,84 0,13 1,40 0,14 6,44<br />
30 53,72 17,30 11,68 0,13 1,73 0,15 5,88<br />
L’évolution de la cellulose et des hémicelluloses au cours du développement du<br />
champignon a été déterminée en calculant le coefficient de cristallinité (Wikberg et Maunu,<br />
2004) et l’intensité relative du pic 103 (Gilardi et al., 1995) respectivement. D’après les<br />
résultats obtenus, on remarque que le mycélium est capable de dégrader simultanément des<br />
hémicelluloses et de la cellulose au cours de sa croissance avec une attaque préférentielle de<br />
la cellulose amorphe, ce qui a été confirmée par l’augmentation du degré de cristallinité. Nos<br />
résultats sont en accord avec les résultats obtenus lors de la transformation du compostage des<br />
boues de station d’épuration et des déchets verts par Albercht et al. (2008), qui ont montré une<br />
augmentation de la cristallinité durant le procédé de co-compostage.<br />
121
Résultats et discussion. Chapitre 4: Transformation de la matière organique<br />
L’estimation de la lignine par le modèle de Giraldi et al. (1995) a permis de révéler la<br />
tendance de la dégradation de la lignine par le mycélium. En effet, la dégradation passe par<br />
deux étapes fondamentales: la première étape, entre le 6 ème et le 24 ème jour d’incubation,<br />
consiste en la dégradation de la lignine riche en sous-unités syringyl, alors que la deuxième<br />
étape, entre le 24 ème et 30 ème jour d’incubation, consiste en la dégradation de la lignine riche<br />
en sous-unités guaiacyl avec des taux différents. Vane et al. (2006) ont rapporté de semblables<br />
résultats et ceci pourrait être expliqué par une dégradation préférentielle des micro-<br />
organismes des unités syringyl par rapport aux unités guaiacyl. En effet, Rio et al., (2001,<br />
2002) ont suggéré que cette préférence de dégradation des unités syringyl est reliée à la<br />
prédominance de liaisons éther en position C4 du cycle aromatique, alors que les unités<br />
guaiacyl incluent une part de liaisons C-C en C5, (position libre i.e. sans groupement<br />
méthoxyl), de laquelle résulte le fort degré de condensation et de résistance aux attaques<br />
fongiques.<br />
De même que la lignine, les groupements CPhenolique subissent des modifications<br />
pendant le développement du mycélium de L. edodes. En effet, ces groupements diminuent de<br />
2,22 à 1,77 après une semaine de croissance et augmentent pour arriver à une valeur 2,18<br />
après un mois de croissance. Cette relative accumulation du groupement CPhenolic a été<br />
reportée par Chen et al. (1989) pendant le compostage et cette accumulation a été attribuée à<br />
la transformation de la lignine et à la polymérisation des monomères phénoliques (diminution<br />
du facteur de l’aromaticité).<br />
3.9. Activités enzymatiques<br />
Les activités enzymatiques mesurées durant la croissance du champignon sur le SOS<br />
sont présentées dans le Tableau 5. Parmi les activités enzymatiques impliquées dans la<br />
décomposition de la matière organique, l’activité des cellulases a été mesurée. Il y a<br />
apparition de cette activité après 6 jours d’incubation et elle atteint sa valeur maximale après<br />
18 jours (1,13 U.g -1 SPS). L’activité lipase présente une augmentation au début de la culture<br />
(de 0,01 à 0,02 U.g -1 SPS), et diminue après 18 jours de culture pour disparaître vers la fin de<br />
l’incubation. Les activités peroxydases, Mn peroxydase et lignine peroxydase, présentent une<br />
augmentation pour arriver à des valeurs maximales (0,231 et 0,321 UI.g -1 SPS du SOS après<br />
18 et 24 jours d’incubation respectivement). Ensuite, les activités peroxydase diminuent.<br />
122
Résultats et discussion. Chapitre 4: Transformation de la matière organique<br />
Les activités laccases suivent relativement la même évolution avec une augmentation de<br />
1,52 à 8,5 UI.g -1 SPS après 18 jours d’incubation, puis une diminution pour arriver à une<br />
valeur de 4,63 UI.g -1 SPS. D’après les résultats, on a remarqué que l’activité laccase est<br />
l’activité majeure parmi les activités enzymatiques impliquées dans la transformation des<br />
polyphénols. A partir de l’ensemble des résultats, on a également remarqué que les activités<br />
cellulases et les deux activités phénoloxydase présentent des tendances similaires avec une<br />
augmentation au début de la culture pour atteindre un maximum, puis une diminution et une<br />
stabilisation jusqu’à la fin de l’incubation. Cette tendance peut être expliquée par une<br />
complémentarité entre ces activités pour aboutir à la dégradation du complexe<br />
lignocellulosique. Par ailleurs, les activités catéchol-1,2-dioxygénase et protocatéchuate-3,4-<br />
dioxygénase n’ont été pas détectées dans les conditions de culture.<br />
Tableau 5 : Mesure des activités (UI.g -1 SPS) cellulase, lipase, laccase, Mn peroxydase,<br />
lignine peroxydase et tannase de L. edodes Le119 durant un mois d’incubation à 25°C.<br />
Temps Cellulase Lipase Laccase Mn Lignine Tannase<br />
(jour)<br />
peroxydase<br />
peroxydase<br />
0 0.02 0.001 1,52 0,04 0.001 0.004<br />
6 0.4 0.002 5,838 0,124 0,121 0.121<br />
12 0.82 0.0014 8,055 0,142 0,245 0.225<br />
18 1.13 0.0018 8,518 0,231 0,214 0.154<br />
24 0.76 0.010 7,602 0,1789 0,321 0.174<br />
30 0.41 nd 4,630 0,1245 0,281 0.110<br />
4. Conclusion<br />
Les résultats de l’étude ont montré une bonne croissance de L. edodes Le119 sur le<br />
SOS en FMS. L’inoculation du SOS avec un mycélium produit sur les grains de blé avec des<br />
margines à 10% (v/v) a permis d’éviter la contamination de culture par des antagonistes. Le<br />
suivi respiromètrique de L. edodes Le119 cultivée sur le SOS a mis en évidence la présence<br />
de trois phases de croissance. L’étude des paramètres de sucres réducteurs, sucres totaux,<br />
phénols totaux ainsi que les spectres RMN du 13 C a montré que la première phase de<br />
croissance a été attribuée à la minéralisation des matières facilement dégradables, suivie par la<br />
phase de dégradation des polysaccharides (hémicelluloses, cellulose). La dernière phase de<br />
croissance correspondait à la dégradation légère de la lignine provenant du bois de taille.<br />
L’étude des cinétiques enzymatiques au cours de la croissance de L. edodes Le119 en FMS<br />
123
Résultats et discussion. Chapitre 4: Transformation de la matière organique<br />
sur SOS a mis en évidence la présence d’un cocktail enzymatique qu’on pourrait utiliser pour<br />
des applications biotechnologiques en particulier dans l’industrie agroalimentaire.<br />
124
Résultats et discussion. Chapitre 5: Disponibilité des substrats oléicoles<br />
Chapitre V<br />
Disponibilité des substrats oléicoles : séchage solaire sous serre du<br />
mélange de grignons d’olive et de margines<br />
L’industrie oléicole génère d’énormes quantités de grignons d’olive et de margines<br />
dans une période courte de quatre mois entre Novembre et Février. Avec l’humidité élevée<br />
des grignons d’olive (35%) et des margines (90%), ces substrats sont facilement altérables par<br />
le développement de la microflore endogène (Lakhtar 2006 ; Roussos et al., 2006). Par<br />
conséquent, il est apparu nécessaire d’envisager un procédé permettant la réduction de leur<br />
humidité et assurer leur conservation pour une disponibilité pour toute l’année. Le séchage<br />
solaire pourrait être un processus d'un grand intérêt pour la stabilisation des ces substrats.<br />
Le séchage est l’une des méthodes courantes de conservation de la plupart des<br />
produits agroalimentaires et des fourrages, par diminution de leur teneur en eau jusqu’à des<br />
valeurs résiduelles où le développement de tout micro-organisme est inhibé. Le stockage est<br />
alors possible dans des conditions ambiantes (Kooli et al., 2007).<br />
Depuis les premiers travaux de Lewis (1921) et de Sherwood (1929) sur le séchage<br />
solaire, cette technique a fait l’objet de nombreuses communications scientifiques mais<br />
demeure encore de nos jours un domaine de recherche privilégié notamment dans les pays où<br />
l’utilisation de séchage solaire s'impose (Kooli et al., 2007).<br />
Dans ce chapitre, nous présentons une étude préliminaire d’un nouveau procédé<br />
consistant à mélanger les grignons d’olive frais avec les margines d’une unité de trituration<br />
triphasique avec le système de presse, et sécher ce mélange dans une serre solaire pour réduire<br />
l’humidité à 10%.<br />
Ce chapitre a fait l’objet d’un article accepté avec révisions dans la revue Journal of<br />
Environmental Management.<br />
125
Résultats et discussion. Chapitre 5: Disponibilité des substrats oléicoles<br />
Solar drying in greenhouse of a mixture of olive cake and olive mill<br />
wastewater, a solution for olive mill wastewater disposal?<br />
Authors:<br />
Hicham LAKHTAR 1 , Hervé MACARIE 1 , Mustapha ISMAILI-ALAOUI 2 , Isabelle<br />
PERRAUD-GAIME 1 , Sevastianos ROUSSOS 1*<br />
Author’s address:<br />
1 IRD, UMR <strong>IMEP</strong> (CNRS / IRD), Aix-Marseille Université, Faculté des Sciences de St.<br />
Jérôme, Boîte 441, 13397 Marseille cedex 20, France,<br />
Email: h.lakhtar@univ-cezanne.fr, herve.macarie@ird.fr, i.gaime@univ-cezanne.fr<br />
2 IAV –Hassan II, Laboratoire des Bioconversions ; BP. 6202-Instituts, 10101 Rabat,<br />
Morocco, m.ismaili@iav.ac.ma<br />
*Corresponding author:<br />
Sévastianos Roussos: Institut Méditerranéen d'Ecologie et de Paléoécologie UMR CNRS/IRD<br />
193; <strong>IMEP</strong> Case 441; FST Saint Jérôme; Université Paul Cézanne; Av. Escadrille<br />
Normandie-Niemen; 13397 Marseille cedex 20; France ;<br />
E-mail: s.roussos@univ-cezanne.fr<br />
Tel: (+33) 04 91 28 91 03<br />
Fax: (+33) 04 91 28 86 68<br />
126
Résultats et discussion. Chapitre 5: Disponibilité des substrats oléicoles<br />
Sechage solaire sous serre d’un mélange de grignons d’olive et de margines,<br />
une solution pour le traitement des margines?<br />
RESUME<br />
Au Maroc, pays producteur de l’huile d’olive, l’industrie oléicole connait actuellement<br />
une forte expansion. Les margines qui sont déversées dans l'environnement causent une<br />
pollution environnementale grave, tandis que les grignons d’olive sont traités par la filière de<br />
traitements appropriés. Dans le présent travail, un nouveau procédé intégré est décrit pour<br />
réduire les volumes des margines produites par un système triphasique utilisant la technique<br />
de presse. Notre procédé consiste à utiliser des grignons d'olive frais comme un agent<br />
absorbant de margines produites puis à sécher le mélange dans une serre pour obtenir une<br />
humidité finale de 10% (poids / poids). Les conditions optimales de mélange de grignons<br />
d’olive avec les margines ont été déterminées à la suite de plusieurs essais de mélange-<br />
séchage. Les performances expérimentales de séchage du mélange grignons d’olive-margines<br />
ont été évaluées. La température de séchage de l'air asséchant a varié de 10 à 53 °C pendant le<br />
séchage. Le mélange a été séché 3 fois plus vite dans la serre qu'à l'extérieur. Enfin, une<br />
caractérisation du mélange séché par l’analyse à la RMN a été effectuée pour prévoir son<br />
utilisation future et de déterminer si ce substrat lignocellulosique pourrait être valorisé dans<br />
les voies existantes pour les grignons d'olive brutes.<br />
Mots clé: grignons d’olive, margines, serre, séchage solaire, RMN.<br />
127
ABSTRACT<br />
Résultats et discussion. Chapitre 5: Disponibilité des substrats oléicoles<br />
Morocco is a country which produces olive oil extensively and this industry within the<br />
country is currently under huge expansion. The olive mill wastewater (OMW) which is being<br />
dumped in the environment results in serious environmental pollution while olive cake (OC)<br />
does have a proper disposal. In the present work, a new integrated process is described to<br />
reduce the OMW produced by a triphasic system using the technique of pressing. Our process<br />
consists in using olive cake as an absorbing agent of OMW produced and then to dry the<br />
mixture in a greenhouse to get a final moisture of 10% (weight/weight). The optimal<br />
conditions of mixing of OC with OMW were determined following various trials of mixing-<br />
drying. The experimental performance of OC-OMW mixture drying in greenhouse was<br />
evaluated. The drying air temperature varied from 10 to 53°C during drying. The mixture was<br />
3 times dryer in the greenhouse than outside. Finally, a characterization of the dried mixture<br />
by NMR analysis was performed to predict its future utilization and determine whether this<br />
lignocellulosic material might be valued in the ways existing for crude olive cake.<br />
Keywords: Olive cake, Olive mill wastewater, greenhouse, solar drying, NMR.<br />
128
1. Introduction<br />
Résultats et discussion. Chapitre 5: Disponibilité des substrats oléicoles<br />
Mediterranean basin concentrates 97% of the world olive oil production (Ismaili-<br />
Alaoui, 2006). Two technologies exist for oil extraction, the oldest three-phase and the more<br />
recent two-phase. The three-phase which generates two wastes (olive mill wastewater and<br />
olive cake) remains the dominating technology in the south of the Mediterranean bassin. It is<br />
estimated that some 3x107 m3 of olive mill wastewater, accompagnied by 5x105 tons of olive<br />
cake are generated each year as a result of olive oil production and this in a very short time of<br />
4 months (November to February) (Gögüs and Maskan, 2006). With a production of 50,000<br />
tonnes of olive oil per year, Morocco is presently located at the sixth rank of the world olive<br />
oil producers. Fifty percent of this production still comes from traditional mills commonly<br />
called “Maasra” which use the classic “three-phase system” with the technique of pressing.<br />
Due to its residual oil content (10-30 % w/w) which depends on the extraction<br />
technology and also to its other components, olive pit (42-54% w/w), skin (10-11% w/w) and<br />
pulp (21-33% w/w) (Lopez-Villatta, 1998), OC has been the subject of many valorization<br />
studies. To cite some of them, it has been evaluated for instance as animal feed (Haddadin et<br />
al., 2002), as raw material for glycolipids’ biosynthesis (Bednarski et al., 2004), as<br />
combustible in traditional furnaces of small building material factories (pottery works, brick<br />
manufactures, etc.) (Masghouni and Hassairi, 2000) and as substrate for mushroom<br />
cultivation (Zervakis et al., 1996) or enzyme production (Cordova et al., 1998). In a country<br />
such as Morocco, its main use is still limited to the recovery of oil by extraction with hexane<br />
for the production of soap (Serghini et al., 2007). The de-oiled OC which has excellent<br />
calorific properties is usually reused directly as combustible by the OC oil extraction units to<br />
dry OC to 6-10% (w/w) humidity before the extraction step. As a consequence, OC should<br />
not be considered as a waste and it is actually rarely dumped into the environment.<br />
Contrary to OC, OMW which contains high concentrations of toxic phenolic<br />
compounds (Di Serio et al., 2008) and organic matter (100-200 g COD/l) is extremely<br />
hazardous for the environment especially the water bodies. Until recently in Morocco, for<br />
instance the potabilization plants furnishing the water to the city of Fes, which concentrates<br />
40% of the country olive oil extraction capacities, had to be put out of operation during the<br />
olive milling campaigns. This was due to the worsening of the Sebou river water quality<br />
resulting from the direct discharge of OMW (Foutlane et al., 2002). Owing to economic<br />
constraints, up to now, no widely accepted solutions exist for the disposal of this waste.<br />
129
Résultats et discussion. Chapitre 5: Disponibilité des substrats oléicoles<br />
Nowadays, in most countries which still use the three phase extraction technology, the<br />
current practice to deal with OMW consists either to use it as amendment in olive fields or to<br />
store it in evaporation ponds in order to reduce the volume to dispose off. Field amendment<br />
which has the advantage to return to the soil part of the nutrients (N, P, minerals) exported<br />
with OMW is a highly controversial practice in the scientific community (Mekki et al., 2007),<br />
while the efficiency of the evaporation ponds is often very poor despite their high<br />
construction costs to avoid OMW infiltration into the ground water.<br />
OMW drying is actually a slow process since the olive oil and some mucilage cover<br />
the surface and make it impermeable to water (Jimenez and Lao, 2004). As so, OMW drying<br />
could be improved by increasing the surface of exchange with the atmosphere through the<br />
addition of a high surface absorbing solid matrix. The OC readily available from the same<br />
olive mill could be a good candidate for that. Thus, the amount of wastes, generated by olive<br />
mill, could be controlled, based on the assumption that the resulting product could be<br />
valorised in the same way as crude OC.<br />
Open sun drying is the common drying method of agricultural materials used in<br />
Morocco. However, this method results in long drying times and quality loses since the drying<br />
materials are directly exposed to environmental conditions (Erbay and Icier, 2009). On the<br />
other hand, artificial hot air dryers have high investment cost and high energy expenses<br />
(Colak and Hepbasli, 2007). However, solar drying in greenhouse can be considered as an<br />
advancement of natural sun drying and it is a more efficient technique of utilizing the<br />
potential solar energy (Müller et al., 1989; Jain and Tiwari, 2004; Kooli et al., 2007; Janjai et<br />
al., 2009). Many research and performance studies have been reported on solar drying in<br />
greenhouse (Kooli et al., 2007; Janjai et al., 2009). Therewith, certain studies were focused on<br />
OC drying for soap making, animal feed or fertilizer (Haddadin et al., 2002; Akgun and<br />
Doymaz, 2005). Nevertheless, there have been no reports on the drying of the mixture of olive<br />
cake with OMW in the literature.<br />
The objectives of the present study were to determine under which conditions all the<br />
OMW generated by a mill could be absorbed by the OC produced by the same mill and if the<br />
resulting OC-OMW mixture could be dried to 10% (w/w) humidity in order to obtain a solid<br />
matrix which could be valorized in the same way as crude OC. To achieve these objectives<br />
we (1) performed a mass balance of the different products generated by a traditional maasra<br />
mill (2) determined the OMW retention capacity by the mill OC, (3) performed several drying<br />
130
Résultats et discussion. Chapitre 5: Disponibilité des substrats oléicoles<br />
experiments in open sun and in a greenhouse under natural convection and (4) determined the<br />
final quality of the dried material and compared it to that of dried crude OC.<br />
2. Materials and methods<br />
2.1. Origin of samples<br />
OC and OMW were obtained from an olive oil mill plant located in Beni-Mellal in the<br />
centre of Morocco. This mill used a three phase system with press and operated mostly with<br />
the “picholine marocaine” which is the main olive variety cultivated in Morocco for<br />
production of both olive oil and table olives. As so this mill can be considered as<br />
representative of the ones used in the country.<br />
2.2. Description of the greenhouse dryer<br />
The greenhouse dryer, used in the study was located at HASSAN II Agronomic and<br />
Veterinary Institute, in Rabat, Morocco. It was built on a concrete floor and oriented in the<br />
north-south direction. It had a parabolic cross-section and a size of 7 (length), 3 (width) and<br />
2.5 m (height) giving on the ground a surface of 21 m2. It was covered with polycarbonate<br />
but the facade was made of transparent extern stratum. The air ventilation was obtained by<br />
some openings at different levels of the sides. One set of iron shelves with iron trays was used<br />
to place the products to be dried. The trays were located on three different levels (0.5, 1 and<br />
1.5 m from the ground) on the shelves. The free space between them was sufficient for<br />
loading and unloading the products. The trays were made of a wire netting to allow air<br />
circulation. A second set of identical shelves and trays were used for the outdoor drying. The<br />
structure of the greenhouse and of the shelves and trays is shown in Fig. 1.<br />
Figure 1. General view of the greenhouse solar dryer (left) and of its interior with the shelves<br />
(right). Only the lower and upper trays were used for loading the material to be dried.<br />
131
Résultats et discussion. Chapitre 5: Disponibilité des substrats oléicoles<br />
2.3. Determination of the OMW retention capacity by olive cake<br />
This test consisted to contact 3 kg of crude OC with 3 L of OMW under gentle<br />
periodic mixing. Every 15 min, an homogenous sample of the mixture was taken from which<br />
the solids were recovered by filtration from the liquid and weighted (Sebabi et al., 2003). The<br />
sampling was repeated until the weight of the solid phase did not change over 3 successive<br />
measurements. The moisture of the solid phase was also determined at each sampling by<br />
drying a subsample at 105°C in an oven for 24 hours. Humidity was determined in the same<br />
way for all the other experiments. Besides the estimation of the maximum amount of OMW<br />
which can be absorbed by OC, this test allowed to determine the minimal time required to<br />
reach this objective. These experiments were performed in triplicate.<br />
2.4. Drying experiments<br />
Outdoor and greenhouse drying were performed under identical conditions in order to<br />
be comparable. Saturated OC with OMW (no free liquid OMW present) were distributed<br />
uniformly in thin layer on the trays the bottoms of which were covered by plastic mesh. The<br />
solids on the trays were turned manually once every hour. The drying process was pursued<br />
until reaching the target moisture. For that, the whole solid present on each trays were<br />
weighted periodically at 1 h intervals using a digital balance and their moisture content was<br />
determined. Solar radiation and temperature inside and outside the greenhouse were<br />
determined periodically. In order to avoid the interference of rain on the outdoor drying<br />
process, it was planned to cover the shelves with a plastic sheet during rainfall events. Finally<br />
this was unnecessary since no rain occurred during all the experimental period (February<br />
2008).<br />
2.5. Chemical characterization of dried crude olive cake and OC-OMW mixture<br />
OC and the new product (OC-OMW mixture) both dried at 10% (w/w) humidity in the<br />
same conditions were characterized for their content in carbon, nitrogen (proteins), fats and<br />
total phenols. Carbon and nitrogen concentrations were determined by a combustion-gas<br />
chromatography technique (Flash EA 1212 Elemental Analyzer, France). The instrument was<br />
calibrated with aspartic acid standard. The amount of proteins was estimated from the<br />
nitrogen content using the general factor of 6.25 (Weinberg et al., 2008). Fats were obtained<br />
by Soxhlet extraction with hexane as solvent according to established procedures (Cordova et<br />
al., 1998). Phenol content was assessed as follows: 10 mL of 60% (v/v) ethanol-water were<br />
added to 1 g of milled solid sample. The mixture was homogenized, incubated for 1 h at 4°C<br />
132
Résultats et discussion. Chapitre 5: Disponibilité des substrats oléicoles<br />
and centrifuged at 4500 rpm for 10 min. The extraction was repeated three times. The<br />
supernatant was then filtered under vacuum. The non phenolic fraction still presents in the<br />
filtered liquid (e.g. fats) was eliminated by extraction with 10 mL of n-hexane, repeated three<br />
times. The total phenolic content of the remaining aqueous phase was quantified by the<br />
method of Folin and Ciocalteu (Bärlocher et al., 2005). The blue reactive complex formed<br />
was dosed spectrophotometrically at 760 nm. Caffeic acid being used as standard for the<br />
calibration, the phenolic content of the samples was expressed as caffeic acid equivalent.<br />
2.6. NMR analysis of dried crude olive cake and OC-OMW mixture<br />
Dried crude OC and the mixture OC-OMW were used directly in NMR experiments<br />
without special preparation. Owing to their size, the samples were gently ground (IKA<br />
Labortechnik, Germany) in order to fill the rotor. Solid-state 13 C CPMAS NMR spectra, of<br />
dried crude OC and dried mixture OC-OMW, were obtained on a Bruker Avance-400 MHz<br />
spectrometer (Bruker, Bremen, Germany) operating at a 13 C resonance frequency of<br />
100.7 MHz. Samples were placed in a 7-mm zirconium rotor and spun at the magic-angle at<br />
6 kHz. All measurements were made at room temperature. The 13 C chemical shifts were<br />
referenced to tetramethylsilane and calibrated with the glycine carbonyl signal, set at<br />
172.5 ppm.<br />
Deconvolution of the NMR spectra was performed using the DmFit software (Massiot<br />
et al., 2002). This software adjusts the spectra to obtain the linewidth and the peak positions<br />
(in ppm) and to integrate each peak so as to obtain the percentage of each contribution. The<br />
aromaticity of dried mixture OC-OMW and crude OC was calculated by expressing aromatic<br />
C (110–165 ppm) as a percentage of aliphatic C (0–110 ppm) + aromatic C (Wikberg and<br />
Liisa Maunu, 2004). The lignin content was calculated according to the model of Gilardi et al.<br />
(1995), which takes into account the total lignin carbon. These authors calibrated lignin<br />
content from aromatic carbons plus aliphatic carbons of lignin. The cellulose crystallinity (Xc)<br />
was expressed by the ratio between the signal at 84 ppm, assigned to the C-4 of amorphous<br />
cellulose, and the signal at 89 ppm, assigned to the C-4 of crystalline cellulose (Newman and<br />
Hemmingson, 1990).<br />
2.7. Statistical treatment of results<br />
Data of drying, chemical analysis and NMR spectra were subjected to analysis of<br />
variance (ANOVA). The error square was obtained from the analysis of variance and used to<br />
calculated the least significant difference (LSD, P=0.05). Variations between data are<br />
significant when the difference between is more than the LSD.<br />
133
3. Results and discussion<br />
Résultats et discussion. Chapitre 5: Disponibilité des substrats oléicoles<br />
3.1. Waste balance in the model traditional mill and liquid retention capacity of olive<br />
cake<br />
Three different pressing of 600 kg of olives in such a mill produced in average 127 kg<br />
of olive oil, 220 kg of OMW (volumetric mass of 1.014 kg/L) and 253 kg of OC with an<br />
initial moisture of 35% (w/w). This gave a production ratio of 87 kg OMW / 100 kg OC.<br />
OMW absorbed<br />
(L/100 kg of OC<br />
80<br />
60<br />
40<br />
20<br />
0<br />
0 20 40 60 80 100<br />
Figure 2. Retention capacity of olive mill wastewater by crude olive cake at 25°C. Error bars<br />
represent standard deviation.<br />
Time (min)<br />
The kinetics of OMW absorption by OC, indicated that 60 min were necessary to<br />
completely saturate OC with OMW and that the maximum retention capacity of OC was of 62<br />
L of OMW / 100 kg of OC giving a final product with moisture of 56% (w/w) (Fig. 2). This<br />
means that the amount of OC generated by this type of mill was only sufficient to absorb<br />
around 72% of the OMW produced at the same site. The sole way, to absorb the remaining<br />
OMW without the need of adding extra exogenous absorbing material, would be to recover<br />
the original absorption capacity of OC or at least part of it. This may be achieved through<br />
partial drying of the OC-OMW mixture obtained and its reuse as new absorbing agent.<br />
Therefore, the objective to absorb all the OMW generated by a mill on the OC may then be<br />
only achieved by a two-step absorption-drying process<br />
134
Résultats et discussion. Chapitre 5: Disponibilité des substrats oléicoles<br />
3.2. Drying of the mixture of OMW-Olive cake<br />
After a set of trials, it was found that the best way to perform the two step absorption-<br />
drying was the one shown in Table 1.<br />
Table 1. Optimized procedure for the two-step drying process of OMW absorbed on OC.<br />
The first step consisted to saturate OC with OMW (resulting product with 56%<br />
moisture w/w) and then to limit the drying of the mixture up to a final moisture of 40% (w/w).<br />
This level of humidity was found as a good compromise to achieve three objectives. It<br />
allowed the evaporation of enough water (42 L from 166 kg of OC-OMW mixture) to give to<br />
the partial dried OC-OMW mixture the capacity to absorb all the OMW remaining to absorb<br />
(25 L). It permeated to avoid the problems generated by a more intensive drying which could<br />
have affected the absorption capacity, because OC became more water repellent upon drying<br />
due to the shrink of the water absorbing solid material that it contains resulting in a loss of<br />
porosity of this material, porosity which is the driving motor of absorption (Liu and Lee,<br />
2006). It also allowed a fast drying for this first step since the drying from a humidity of 56%<br />
to 40% (w/w) concerned mostly to free water which is easy to be removed (Ekechukwu,<br />
1999). Quantitatively, the total mass of by-products (OC and OMW) generated by a maasra<br />
mill (78 kg / 100 kg of olives processes) can be reduced by 56.15% (final amount of 35-36 kg<br />
/ 100 kg of olives processes) thanks to such drying.<br />
135
Résultats et discussion. Chapitre 5: Disponibilité des substrats oléicoles<br />
3.3. Drying kinetics of OC-OMW mixture<br />
A typical pattern of the first and second step of drying of OC-OMW mixture outdoor<br />
and in the greenhouse is shown in figures 3 and 4. The data shown are for an OC-OMW<br />
loading of 10 kg.m -2 . Solar radiation varied from 5 to 10 W.m -2 during the drying period.<br />
Temperature (°C)<br />
Temperature (°C)<br />
Time (h)<br />
Time (h)<br />
Figure 3. Change of temperature inside the greenhouse ( ) and open sun ( ) during the (a)<br />
first drying and (b) second drying of OC-OMW mixture in Rabat-Morocco during February<br />
2008.<br />
The evolution of the temperature in the greenhouse was as expected from what is<br />
published in the literature for such systems (Ivanova and Andonov, 2001; Jain and Tiwari,<br />
2004; Kooli et al., 2007; Janjai et al., 2009). It was significantly different from the outdoor<br />
temperature during the day time and part of the night (9:00 am to 11:00 pm) but identical to<br />
the outdoor temperature after 11:00 pm and up to 9:00 am (Fig. 3). In average, the air in the<br />
dryer was at 16°C higher than outside during the first period. This difference reached a peak<br />
at noon when the air temperature inside the greenhouse became higher than or equal to 50°C,<br />
a condition which was maintained for 3 hours (12:00 am to 3:00 pm). At this temperature the<br />
a<br />
b<br />
136
Résultats et discussion. Chapitre 5: Disponibilité des substrats oléicoles<br />
heat gain in the greenhouse was 10% higher than the outside. In accordance with the<br />
increased temperature and heat gain, the drying of the OC-OMW mixture was much faster in<br />
the greenhouse than outside (Fig. 4).<br />
Moisture content (%)<br />
Figure 4. Typical drying curve of the OC-OMW mixture in the greenhouse ( ) and open sun<br />
( ) in Rabat-Morocco during February 2008; (a) First drying; (b) second drying.<br />
For the first drying, height hours of the day time were sufficient to reach the target<br />
moisture (40% w/w) in the greenhouse compared to 12 hours outside, which means that the<br />
drying process was some 1.5 times quicker in the greenhouse. For the second drying, the<br />
products obtained after 35 hours of drying including a night during which a very low loss of<br />
moisture occurred (less than 5% w/w in 10 hours) were also 3 times dryer in the greenhouse<br />
than outside (10% moisture against 30% w/w). The main limiting factor in the drying process<br />
corresponds to the very low drying efficiency achieved during the night period. This<br />
efficiency could be improved by implementing forced convection through the use of a fan<br />
powered with batteries recharged in the day by means of solar cell modules installed on the<br />
top of the greenhouse (Chow, 2009). Further, it is possible to imagine a complete automation<br />
of the absorption drying-process. OC may be loaded in the greenhouse with Archimedes<br />
screws and humidified with OMW through a dispositive of showers which could allow<br />
controlling precisely the humidity of the mixture.<br />
a<br />
Time (h)<br />
b<br />
137
Résultats et discussion. Chapitre 5: Disponibilité des substrats oléicoles<br />
3.4. Effect of the amount of OC-OMW mixture to be dried and location in the<br />
greenhouse<br />
To test the impact of these two parameters on the drying efficiency, five different<br />
amounts (10 to 35 kg/m 2 ) of OC-OMW mixture were loaded on the trays located in the lower<br />
and upper part of the greenhouse. The mixtures were dried during 2 days. Their final moisture<br />
is shown in Figure 5. Without surprise, higher was the amount to dry and slower was the<br />
drying process resulting in a product with higher moisture after the same time of drying. In<br />
this particular case, only the lowest amount tested (10 kg fresh weight/m 2 ) could be dried up<br />
to 10% moisture during the length of the experiment. A slight difference of moisture could be<br />
observed among the products dried in the lower and upper trays, but the difference was<br />
statistically not significant (P=0.05).<br />
Moisture content (%)<br />
Sample<br />
Figure 5. Effect of the amount of OC-OMW mixture to be dried and of its location in the<br />
greenhouse, bottom ( ) and top ( ) on the final moisture of the substrate after drying for 2<br />
days. The initial moisture of the mixture was of 56% (w/w).<br />
3.5. Chemical characterization of dried crude OC and OC-OMW mixture<br />
Chemical analysis results for the dried crude OC and the dried OC-OMW mixture are<br />
presented in table 2. Statistical analysis (P=0.05) showed significant differences in pH, C, N,<br />
C/N and total phenols, but no significant difference in fat content. Incorporation of OMW into<br />
OC allowed to increase by 24% of the protein content in the mixture compared to dried crude<br />
OC which is considered as a poor material for animal feed (Martin Garcia et al., 2003).<br />
However, this was counterbalanced by total phenol content which was 5 times higher than<br />
that in the dried crude OC since phenols are known for their anti-digestive properties<br />
(Grabber, 2009). On the other hand, this converted the OC-OMW mixture into a good raw<br />
Thickness (kg .m- 2 )<br />
138
Résultats et discussion. Chapitre 5: Disponibilité des substrats oléicoles<br />
material candidate for the extraction of antioxidants which are used in the food, cosmetic and<br />
pharmaceutical fields (Erbay and Icier, 2009). The fat content and dryness of the mixture<br />
indicates also that it could be used directly by the traditional oil recovery units in place of OC.<br />
Table 2. Global chemical composition of the OC-OMW mixture and crude OC both dried at<br />
10% moisture*<br />
Component OC-OMW dried mixture Dried crude OC<br />
pH 5.1± 0.1 4.9+0.2<br />
C (%) 52.49 ± 0.01 51.11 ± 0.18<br />
N (%) 1.06 ± 0.01 0.85 ± 0.09<br />
C/N 49.51 ± 0.64 59.77 ± 2.00<br />
Protein (%) 6.62 ± 0.08 5.34 ± 0.56<br />
Olive oil (%) 10.00 ± 1.02 10.00 ± 0.54<br />
Total Phenol (%) 4.1 ± 0.81 1.20 ± 0.15<br />
*All % in w/w, the values after ± represent standard deviation<br />
3.6. NMR analysis of OMW-olive cake<br />
The 13 C NMR spectra of dried crude OC and dried OMW-OC mixture are presented in<br />
Figure 6. The peak assignments and interpretations (Table 3) are based on previous NMR<br />
studies of organic matter (Albrecht et al., 2008; Baddi et al., 2004; Farnet et al., 2009).<br />
Figure 6. Solid-state 13 C NMR spectra of, (a) dried OC-OMW mixture and (b) dried<br />
crude OC.<br />
a<br />
b<br />
139
Résultats et discussion. Chapitre 5: Disponibilité des substrats oléicoles<br />
Table 3. 13 C-NMR spectral assignment of C functional groups<br />
Chemical shift (ppm) Assignment Compounds included<br />
0-45 Alkyl C Lipids, Amino-acids,<br />
45-60 N-Alkyl/methoxyl C Amino acids<br />
60-110 O-Alkyl C Celluloses, hemicellulose, polysaccharides<br />
110-145 Aryl C Polyphenols, Lignin (aromatic C)<br />
145-165 O-Aryl C Phenols, lignin (phenolic C)<br />
165-190 Carboxyl C Organic acids, amide and ester C<br />
Deconvolution of spectra allowed integration of 27 peaks. Figure 7 showed relative<br />
integration values for the major C-types from the dried substrates. A comparison of the<br />
spectra of dried OC-OMW mixture and crude OC showed that the major difference are in the<br />
Alkyl C (0-45 ppm), O-alkyl C (60-110 ppm) and Carboxyl C (165-190 ppm). The addition of<br />
OMW to OC led to an apparent low content of carbohydrate in the dried mixture. The high<br />
intensity of O-alkyl C in the dried mixture was consistent with total phenol content which was<br />
5 times higher than in OC. The dried mixture showed more intense peaks at 175 ppm, because<br />
of the carboxyl group which can result from the degradation of OMW upon drying,<br />
especially, during the first stage of drying. The intensities of the peaks at 84 and 89 ppm<br />
showed a very low degree of crystallinity of cellulose both for the dried OC-OMW mixture<br />
and OC and the lignin content was not affected by the additions of OMW (Table 4).<br />
Integration value (%)<br />
Chemical shift (ppm)<br />
Figure 7. Integration values for the major C-types in the 13 C NMR spectra of dried OC-OMW<br />
mixture ( ) and dried OC ( ).<br />
140
Résultats et discussion. Chapitre 5: Disponibilité des substrats oléicoles<br />
Table 4. Calculated parameters from NMR data of dried mixture OC-OMW and dried<br />
crude OC.<br />
Parameters OC-OMW dried mixture Dried crude OC<br />
Aromaticity % 17.42±0.26* 16.78±0.33<br />
Lignin% 25.23±1.24 25.81±1.31<br />
Carbohydrate% 47.11±1.02 53.00±2.32<br />
Crystallinity 89/84<br />
ppm 0.15±0.01 0.15±0.02<br />
*The values after ± represent standard deviation<br />
4. Conclusions<br />
The absorption of all the OMW produced by an olive mill on the OC generated by the<br />
same mill was found to be possible. It was carried out in two steps with alternate drying. The<br />
experimental results showed that the drying of the OC-OMW mixture in greenhouse was very<br />
effective, and that the drying period to achieve the same level of moisture was shortened<br />
relatively to the open sun drying. The homogeneity of air in the greenhouse under natural<br />
convection eliminated any difference in drying rate at different levels of trays. The chemical<br />
and NMR analysis of the dried OC-OMW mixture showed that the new product was<br />
sufficiently similar to OC to be at least valorised in the same way with the advantage to avoid<br />
the dumping of OMW in the environment.<br />
Acknowledgements:<br />
Hicham Lakhtar is thankful to DSF-IRD France for the award of a PhD fellowship. The<br />
authors are grateful to Dr. Fabio Ziarelli of Paul Cezanne University, France, for providing<br />
the NMR facility.<br />
141
Synthèse générale<br />
142
Synthèse générale<br />
Synthèse générale<br />
Cette partie de synthèse générale a pour but de récapituler les différents résultats<br />
obtenus. Au cours de ce travail, nous avons étudié la valorisation des substrats oléicoles (bois<br />
de taille, grignons d’olive, et margines) par la culture de Lentinula edodes en fermentation en<br />
milieu solide pour :<br />
1. La production de la biomasse mycélienne servant à la production du carpophore pour<br />
l’alimentation humaine<br />
2. Apporter une solution technique permettant la détoxification des effluents des huileries<br />
en particulier les margines d’olive, qui pourrait être destiné à l’alimentation animale<br />
3. Apporter une bonne compréhension de la physiologie de croissance de Lentinula<br />
edodes sur le mélange de substrats oléicoles solides.<br />
Pour atteindre ces objectifs, nous avons fait appel et développé, au cours de ce travail,<br />
des approches microbiologiques (FMS, respirométrie), physicochimiques (dosage des phénols<br />
totaux, sucres réducteurs, monomères phénoliques, ergostérol), enzymologiques (dosages<br />
d’activités, révélation d’isoformes), et spectroscopique (RMN solide du 13 C).<br />
Avec l’augmentation de la production de l’huile d’olive au Maroc, les huileries d’olive<br />
accentuent la pollution de l’environnement, en particulier dans les régions de production (Fès,<br />
Meknès, Marrakech, Béni-Mellal). Cette pollution est due principalement au rejet des<br />
margines d’olives qui contiennent des composés récalcitrants perturbant les écosystèmes<br />
microbiens des réceptacles environnementaux. En effet, ces composés récalcitrants, en<br />
particulier les molécules phénoliques qui exercent une action antimicrobienne et<br />
phytotoxique, limitent les procédés de valorisation de ces effluents qui sont de caractère<br />
saisonnier et produits en grands volumes.<br />
Dans cette vision de développement durable du secteur oléicole, nous avons souhaité<br />
mettre au point un procédé novateur qui converge vers une valorisation intégrale des sous-<br />
143
Synthèse générale<br />
produits de l’industrie oléicole (bois de taille, grignons d’olives et margines) dans le but de<br />
vérifier l’hypothèse suivante : « Est-il possible de valoriser les trois co-produits de l’industrie<br />
oléicole (bois de taille, grignons d’olives et margines) pour en faire un seul substrat oléicole<br />
solide utilisé pour la culture de Lentinula edodes en FMS». Le choix du L. edodes repose sur<br />
trois critères :<br />
La nature du substrat de culture : Contrairement aux champignons de Paris,<br />
Agaricus bisporus, Lentinula edodes n’exige pas un substrat composté et se développe sur des<br />
substrats lignocellulosiques ayant subi peu de transformations.<br />
La commercialisation du champignon : Lentinula edodes (Shiitake) est le deuxième<br />
champignon comestible, commercialisé sur le marché mondial avec une production qui<br />
dépasse deux millions de tonnes en 2002. Cette production augmente de 25% annuellement.<br />
La production des enzymes ligninolytiques : Le potentiel de L. edodes de<br />
production des enzymes ligninolytiques, en particulier les phénoloxydases, a été mis en<br />
évidence par plusieurs auteurs. Ainsi, D’Annibale et al. (2004) ont démontré l’implication des<br />
laccases, produites par L. edodes, dans la dégradation des composés phénoliques, favorisant<br />
ainsi une détoxification des margines.<br />
Pour la culture de L. edodes en FMS sur le SOS, une étude de criblage des souches a<br />
été réalisée pour la sélection d’une souche performante, tolérante aux margines, productrice<br />
d’une biomasse mycélienne importante et capable de dégrader des phénols totaux. Les<br />
résultats de criblage des souches de L. edodes en présence de margines à 10 et 20 % (v/v)<br />
(Lakhtar et al., 2009) ont permis de sélectionner la souche Le119. Cette souche a montré une<br />
tolérance aux margines à 20% avec une production de biomasse mycélienne de 4,49 mg.j -1 L -1 .<br />
En milieu liquide à base de margines à 10%, L. edodes Le119 était capable de dégrader les<br />
phénols totaux de margines à raison de 75% après 30 jours de culture à 25°C et une activité<br />
laccase de 1,5 (UI.mL -1 ) après 15 jours de culture. Ce résultat a permis également de mettre<br />
en évidence la relation d’induction de la production de laccase par la présence des margines.<br />
En effet, une augmentation de l’activité laccase a été remarquée après l’addition des margines<br />
dans les milieux de culture solides et liquides. Par ailleurs, il a été reporté que les laccases<br />
sont impliquées dans la dégradation des molécules phénoliques (D’Annibale et al., 2004) ainsi<br />
que dans le mécanisme de défense de L.edodes à l’égard d’antagonistes tels que les<br />
moisissures (Savoie et Mata, 2003). Une fois la souche de L. edodes Le119 sélectionnée, nous<br />
avons procédé à la production d’inoculum de cette souche sur Ebly ® , un substrat amylacé<br />
commercial.<br />
144
Synthèse générale<br />
Bien que la technique de préparation de l’inoculum des basidiomycètes (spawn) soit<br />
bien décrite dans la littérature (Royse et al., 1998), la complexité des substrats à fermenter<br />
conditionne cette technique pour faire face aux contaminations occasionnées par les<br />
antagonistes opportunistes (moisissures). En effet, les contaminations répétitives des cultures<br />
de L. edodes Le 119 sur le SOS nous a poussé à exploiter les deux critères que nous avons<br />
fixés, entre autres, pour la sélection de L. edodes Le119 (Chapitre 1) qui sont: (i) la tolérance<br />
aux margines et (ii) l’induction de la production des laccases (Chapitre 2).<br />
L’addition des margines à 10% (v/v) aux grains de blé Ebly ® durant la préparation de<br />
l’inoculum a favorisé une bonne production de mycélium avec une activité métabolique<br />
accentuée. En effet, la biomasse produite de L. edodes Le119 sur les grains d’Ebly ® avec<br />
margines 10% était trois fois plus élevée que celle obtenue en absence de margines (1,8 et 0,6<br />
mg.g -1 MS du «spawn» après 20 jours d’incubation respectivement). Quant à l’activité<br />
métabolique, la présence de margines a multiplié l’activité respiratoire du mycélium de<br />
L. edodes cultivé sur les grains d’Ebly ® avec les margines à 10%, tandis que l’activité laccase<br />
passait de 0,2 à 1,7 UI.g -1 SPS au 12 ème jour d’incubation pour le spawn produit sans et avec<br />
margines à 10% respectivement (Chapitre 2).<br />
La deuxième partie de chapitre 2 a concerné certains aspects méthodologiques relatifs<br />
à l’estimation indirecte de la biomasse mycélienne de L. edodes Le119 cultivé sur les grains<br />
d’Ebly ® en FMS par la détermination de l’ergostérol. Compte tenu qu’aucune méthodologie<br />
fiable n’existait dans la littérature, il a été nécessaire de mettre au point le protocole de<br />
l’estimation de la biomasse mycélienne en FMS. De plus, les conditions de culture peuvent<br />
affecter la fiabilité et la reproductibilité de l’estimation de la biomasse. Ainsi, il est apparu<br />
nécessaire d’étudier l’effet de la nature du substrat de culture ainsi que l’âge du mycélium sur<br />
la teneur de l’ergostérol dans le mycélium de L. edodes Le119.<br />
Les résultats obtenus ont montré qu’il ya une forte corrélation (r 2 = 0.987) entre la<br />
teneur en ergostérol et la quantité de biomasse mycélienne correspondante quelles que soient<br />
les conditions de culture (composition du milieu de culture et âge du mycélium). De plus, la<br />
corrélation directe de la teneur en ergostérol du mycélium des produits fermentés avec le taux<br />
de production de CO2 (r 2 = 0,88) a permis d’évaluer l’état physiologique du mycélium par la<br />
détermination de l’ergostérol. Ainsi, cette technique mise au point au cours de ce travail nous<br />
a servi pour l’estimation de la biomasse mycélienne de L. edodes sur le SOS (Chapitres 3<br />
et 4).<br />
145
Synthèse générale<br />
Suite à l’optimisation de la production d’inoculum « spawn » de L. edodes Le119,<br />
nous avons cherché, d’une part, à optimiser la composition du SOS afin d’obtenir une<br />
production importante de la biomasse mycélienne de L. edodes Le119. Le résultat de la<br />
biomasse estimée par quantification d’ergostérol a montré que le mélange optimal SOS est<br />
composé de 20% de bois de taille, 30% de grignons d’olive et 50% de margines. D’autre part,<br />
nous avons adopté la méthodologie des plans d’expériences afin de déterminer l’effet relatif<br />
de 6 facteurs sur la production de biomasse mycélienne ainsi que sur la dégradation rapide des<br />
phénols totaux. Les six facteurs (pH initial, aération, âge d’inoculum, taille d’inoculum, taille<br />
de particules et solution minérale) ont été choisis judicieusement pour recouvrir des<br />
conditions physiques, chimiques et biologiques. L’utilité des plans d’expérience réside dans<br />
le fait que toutes les données expérimentales sont utilisées simultanément pour calculer l’effet<br />
de chaque facteur avec une précision accrue et un faible nombre d'essais. Cette stratégie<br />
convient particulièrement à la culture lente de L. edodes sur le SOS qui demande un temps<br />
d’incubation très long (un à deux mois).<br />
Les deux facteurs essentiels ayant un effet sur les deux réponses, biomasse mycélienne<br />
et dégradation des phénols totaux, sont : (i) le taux d’inoculum et (ii) l’addition de la solution<br />
minérale. En effet, le taux d’inoculum a montré un effet positif sur la quantité de biomasse<br />
mycélienne produite ainsi que sur le taux des phénols dégradés. Un taux minimal de 10%<br />
(p/p) permet d’éviter la contamination des cultures par des antagonistes. Tandis que l’addition<br />
de la solution minérale (Chapitre 4) a présenté un effet négatif sur les mêmes réponses et que<br />
les margines, malgré leur toxicité, contiennent des éléments nutritifs, comme des sucres<br />
solubles, des protéines et des éléments minéraux, qui favorisent le développement de<br />
L. edodes Le119. Les autres facteurs testés n’ont pas montré d’effet sur les deux. Ainsi donc,<br />
les conditions optimales de culture de L. edodes Le119 sur le SOS sont les suivantes :<br />
la composition du SOS (20% de bois de taille, 30% de grignons d’olive et 50% de<br />
margines) ;<br />
pH initial 5 ;<br />
température d’incubation 25°C ;<br />
débit d’aération 20 mL.min -1 ;<br />
âge d’inoculum 10 jours ;<br />
taux d’inoculum 10% ;<br />
particules avec une taille entre 2 et 0,8 mm à raison de 70% et des particules avec une<br />
taille inferieure de 0,8 à raison de 30% ;<br />
sans addition de solution minérale.<br />
146
Synthèse générale<br />
Afin de suivre le développement du mycélium de L. edodes Le119 sur le SOS dans des<br />
conditions optimales des cultures en FMS été effectuées (Chapitre 4). Au cours des FMS nous<br />
avons adopté la technique de respirométrie qui permet le suivi de la croissance de L. edodes<br />
Le119 d’une manière non destructive en analysant le CO2 présent dans les effluents gazeux à<br />
la sortie des colonnes de Raimbault (De Araujo, 2002). Pour ce faire, on a mis au point un<br />
prototype nouveau « PNEO » qui permet de mesurer simultanément quatre paramètres (Débit<br />
d’air, température d’air, humidité relative d’air et taux de CO2). Le suivi respirométrique a<br />
montré que la croissance de L. edodes Le119 cultivé sur le SOS passe par trois phases de<br />
croissance rapide alternées par trois phases de croissance ralentie. La production de biomasse<br />
suit également cette évolution en trois phases pour arriver à une biomasse importante (21,61<br />
mg.g -1 ) après un mois d’incubation à 25°C. L’évolution de production de la biomasse<br />
mycélienne estimée par analyse d’ergostérol qui suit l’évolution de la production de CO2<br />
confirme l’adaptabilité de la technique développée (Chapitre 3) pour estimer l’état<br />
physiologique de L. edodes Le119 en FMS. Par ailleurs, l’analyse des échantillons fermentés<br />
a mis en évidence une dégradation des phénols totaux à 80% durant la croissance de L.<br />
edodes. Cette dégradation des phénols est accompagnée, d’une part, d’une décoloration du<br />
SOS de 85% et, d’autre part, d’une production élevée d’enzymes spécifiques.<br />
Les résultats des analyses spectroscopiques de la RMN ont mis en évidence le<br />
changement de composition en groupement carbonés du SOS durant le développement de<br />
L. edodes Le119 en FMS, en particulier les polysaccharides et la lignine. L’accumulation en<br />
sucres réducteurs enregistré après 12 jours d’incubation confirme la diminution observée dans<br />
les spectres de RMN correspondant aux polysaccharides en particulier aux hémicelluloses.<br />
Avec la composition diversifiée du substrat oléicole solide (SOS) mis au point dans notre<br />
étude (Chapitre 3), le mycélium de L. edodes Le119 a synthétisé un cocktail enzymatique<br />
varié et composé essentiellement de laccases, Mn peroxydases, lignine peroxydases et<br />
cellulases. L’évolution de la composition du cocktail enzymatique ainsi que les spectres de<br />
RMN accompagnés de l’évolution des sucres réducteurs, montre que le mycélium commence<br />
par la minéralisation des matières facilement dégradables (phase 1) et poursuit son<br />
développement en dégradant les polysaccharides, en particulier les hémicelluloses et la<br />
cellulose (phase 2). La dernière phase de croissance correspondrait à une légère dégradation<br />
de la lignine provenant principalement du bois de taille, tandis que chaque phase de<br />
ralentissement de croissance (Chapitre 4) serait attribuée à la phase d’organisation du<br />
mycélium pour la préparation de son arsenal enzymatique responsable de la dégradation des<br />
147
Synthèse générale<br />
molécules rencontrées. L’ensemble de ces résultats confirme bien notre hypothèse de départ.<br />
Il est donc possible de cultiver L. edodes Le119 sur un substrat oléicole solide SOS composé<br />
de bois de taille (20%), de grignons d’olive (30%) et de margines (50%). Ainsi, cette étude a<br />
permis de valoriser l’ensemble des substrats oléicoles par la culture de L. edodes.<br />
Enfin, la réussite de la culture de L. edodes Le1119 sur le SOS nous a poussés à<br />
rechercher une technique qui permet d’assurer la disponibilité durant toute l’année des<br />
substrats oléicoles (grignons d’olive et margines) qui sont facilement altérables et de caractère<br />
saisonnier (Novembre et Février). En effet, la teneur élevée en eau de ces substrats, grignons<br />
d’olive (35%) et margines (90%), ainsi que leur composition biochimique, présence des<br />
sucres, de protéines, pectines et autres nutriments, favorisent le développement de la<br />
microflore endogène et posent le problème de conservation de ce matériau. Dans le dernier<br />
chapitre de la thèse, nous avons envisagé d’appliquer le procédé de séchage solaire sous serre<br />
pour le mélange de grignons d’olive avec les margines provenant de la même unité de<br />
trituration. Ce procédé convient parfaitement au procédé de trituration triphasique ainsi que<br />
les conditions météorologiques du Maroc qui se caractérisent par un pouvoir asséchant élevé<br />
de l’air. Les premiers résultats ont montré que la capacité d’absorption des margines par les<br />
grignons d’olives (62 kg margines par 100 kg de grignons) est plus faible que la quantité des<br />
margines produites (87 kg margines par 100 kg de grignons). Cela nous a amené à procéder<br />
au séchage en deux étapes.<br />
La première étape consiste à humidifier les grignons d’olive par les margines jusqu'à<br />
56% (p/p) d’humidité, en incorporant ainsi environ de 72% de margines, et à leur appliquer<br />
un séchage pour ramener cette humidité à 40%. La deuxième étape de séchage consiste à ré-<br />
humidifier le mélange pour atteindre une humidité de 50% et à le faire sécher jusqu'à 10%. La<br />
saturation en margine des grignons de départ à 56% a permis d’accélérer le premier séchage et<br />
le fait de limiter ce premier séchage à 40% a permis d’incorporer plus facilement par la suite<br />
les margines sur les grignons en vue du deuxième séchage. La caractérisation biochimique du<br />
nouveau mélange séché a montré que l’incorporation des margines permet d’augmenter la<br />
teneur du mélange grignons-margines en protéines de 1,2 fois et en phénols totaux de 5 fois,<br />
comparée à la teneur en protéines et en phénols totaux des grignons bruts. Le spectre de la<br />
RMN solide du 13 C du mélange grignons-margines a été comparé à celui des grignons bruts<br />
séchés dans les mêmes conditions. Le résultat montre qu’il ya une seule différence enregistrée<br />
dans le groupement carboxyle (165-190 ppm) qui pourrait être due à la dégradation des<br />
margines durant la première phase de séchage (Chapitre 5).<br />
148
Synthèse générale<br />
En conclusion, ce travail nous a permis de démontrer qu’il possible de fabriquer, à<br />
partir de l’ensemble des co-produits oléicoles solides (bois de taille et grignons d’olives) et<br />
liquides (margines), un seul substrat oléicole solide (SOS) pour la culture de L. edodes en<br />
fermentation en milieu solide, et que les portions mises au point de chaque co-produit<br />
convient parfaitement aux quantités produites par l’industrie oléicole avec le procédé<br />
triphasique de presse.<br />
149
Conclusions et perspectives<br />
150
Conclusions et perspectives<br />
Conclusions et perspectives<br />
L’objectif de ce travail a été d’étudier la valorisation des substrats oléicoles (bois de<br />
taille, grignons d’olive, et margines) par la culture de Lentinula edodes en fermentation en<br />
milieu solide, pour i) la production de la biomasse mycélienne servant à la production du<br />
carpophore pour l’alimentation humaine et ii) pour la détoxification du SOS qui pourrait être<br />
destiné à l’alimentation animale.<br />
Malgré l’existence des composés récalcitrants en particulier les composés phénoliques<br />
qui confèrent la toxicité des co-produits oléicoles (grignons et margines d’olive), ces substrats<br />
contiennent également des éléments nutritifs qu’on peut exploiter. La culture d’un<br />
champignon qui tolère la présence des composés phénoliques peut tirer parti de ces substrats<br />
pour son développement. Les études de criblage de L. edodes sur des milieux contenant des<br />
margines ont permis de sélectionner une souche très performante et de prévoir l’addition des<br />
margines aux grains de blé durant la préparation de l’inoculum. Les résultats obtenus amènent<br />
à plusieurs conclusions.<br />
Tout d’abord, L. edodes Le119 a montré une bonne tolérance aux margines avec la<br />
production d’une biomasse importante. La présence des composés phénoliques dans les<br />
margines induit la production des phénoloxydases qui servent à la fois à la déphénolisation<br />
des margines et à la défense de L. edodes face aux antagonistes opportunistes tels que les<br />
moisissures.<br />
Les résultats de la méthode d’estimation de la biomasse mycélienne par quantification<br />
d’ergostérol ont clairement démontré l’applicabilité de cette technique à l’estimation de l’état<br />
physiologique du mycélium de L. edodes cultivé en FMS.<br />
De plus la conclusion principale de l’étude sur l’évaluation des facteurs de culture est<br />
que le taux d’inoculation est le facteur le plus important de la réussite de la culture de<br />
L. edodes sur le SOS. Il serait intéressant d’envisager la production de l’inoculum sur des<br />
sous-produits agricoles comme la bagasse de canne à sucre et le son de blé, afin de réduire le<br />
coût de production de l’inoculum.<br />
151
Conclusions et perspectives<br />
La mise au point du prototype du respiromètre « PNEO » a permis de suivre<br />
l’évolution de la production de CO2 par L. edodes Le119 cultivé sur le SOS en FMS. On peut<br />
envisager d’ajouter d’autres détecteurs pour élargir le champ d’application de ce nouveau<br />
dispositif essentiel au suivi en ligne de la croissance et la respirométrie des champignons<br />
filamenteux.<br />
L’association des phases d’activités biologiques (respirométrie, activité enzymatique)<br />
avec des phases de dégradation des constituants principaux du SOS a permis de conclure que<br />
la croissance de L. edodes Le119 sur le SOS n’est pas continue et mériterait d’être modélisée<br />
pour pouvoir orienter cette culture dans des applications biotechnologiques telles que<br />
l’enrichissement en protéines du SOS, la production d’un cocktail enzymatique et la culture<br />
des microorganismes en général.<br />
Ces résultats prometteurs doivent être validés à l’échelle pilote en particulier pour la<br />
production de carpophores. Cette technique de préparation du substrat SOS couplée à une<br />
culture de champignons comestibles et médicinaux ouvre des perspectives très intéressantes<br />
d’une part pour diminuer la pollution autour des moulins de trituration d’olive et, d’autre part,<br />
pour contribuer au développement durable du secteur oléicole non seulement pour le Maroc,<br />
mais aussi pour les autres pays méditerranéens.<br />
152
Références bibliographiques<br />
153
Références bibliographiques<br />
Références bibliographiques<br />
Achak A., Ouazzni N., Yaacoubi A. et Mandi L., 2008. Caractérisation des margines issues<br />
d’une huilerie moderne et essai de leur traitement par coagulation‑floculation par la<br />
chaux et le sulfate d’aluminium. Revue des Sciences de l’Eau. 21 : 53-57.<br />
Ait Baddi G., Cegarra J., Merlina G., Revel J. C. et Hafidi, M., 2009. Qualitative and<br />
quantitative evolution of polyphenolic compounds during composting of an olivemill<br />
waste-wheat straw mixture. Journal of Hazardous Materials. 165: 1119-1123.<br />
Ait Baddi G., Hafidi M., Cegarra J., Alburquerque J. A., Gonzalvez J., Gilard V. et<br />
Revel J.-C., 2004. Characterization of fulvic acids by elemental and spectroscopic<br />
(FTIR and 13C-NMR) analyses during composting of olive mill wastes plus straw.<br />
Bioresource Technology. 93: 285-290.<br />
Ait Baddi G., Hafidi M., Gilard V. et Revel J.C., 2003. Characterization of humic acids<br />
produced during composting of olive mill wastes: elemental and spectroscopic<br />
analyses (FTIR and C-13-NMR). Agronomie. 23: 661-666.<br />
Akgun N.A. et Doymaz I., 2005. Modelling of olive cake thin-layer drying process. Journal<br />
of Food Engineering. 68: 455-461.<br />
Al Mallah K., Asma O.J., Abu Lail N.I., 2000. Olive mills effluent (OME) wastewater posttreatment<br />
using activated clay. Separation and Purification Technology. 20: 225-234.<br />
Alaoui S.M., Merzouki M., Penninckx M.J. et Benlemlih M., 2008. Relationship between<br />
cultivation mode of white rot fungi and their efficiency for olive oil mill wastewaters<br />
treatment. Electronic Journal of Biotechnology. 11: 1-8.<br />
Alarcon-Gutiérrez E., 2007. Influence de facteurs abiotiques sur la régulation des<br />
paramètres microbiens impliqués dans la dégradation de la matière organique d’une<br />
litière forestière méditerranéenne. Thèse de Doctorat, Université Paul Cézanne, Aix-<br />
Marseille <strong>III</strong>, Marseille, 226 pages.<br />
Albrecht R., Ziarelli F., Alarcôn-Gutiérrez E., Le Petit J., Terrom G. et Perissol C.,<br />
2008. 13 C solid-state NMR assessment of decomposition pattern during cocomposting<br />
of sewage sludge and green wastes. European Journal of Soil Science.<br />
59: 445-452.<br />
Alexopoulos C.J. et Mims C.W., 1979. Introductory mycology. Third Edition. John Wiley<br />
and Sons, New York, 340-342.<br />
Alinsafi A., Khemis M., Pons M. N., Leclerc J. P., Yaacoubi A., Benhammou A. et<br />
Nejmeddine A., 2005. Electro-coagulation of reactive textile dyes and textile<br />
wastewater. Chemical Engineering and Processing. 44: 461-470.<br />
Aloui F., Abid N., Roussos S. et Sayadi S., 2007. Decolorization of semisolid olive residues<br />
of "alperujo" during the solid state fermentation by Phanerochaete chrysosporium,<br />
Trametes versicolor, Pycnoporus cinnabarinus and Aspergillus niger. Biochemical<br />
Engineering Journal. 35: 120-125.<br />
Altieri R. et Esposito A., 2008. Olive orchard amended with two experimental olive mill<br />
wastes mixtures: Effects on soil organic carbon, plant growth and yield. Bioresource<br />
Technology. 99: 8390-8393.<br />
154
Références bibliographiques<br />
Altieri R., Esposito A., Parati F., Lobianco A., et Pepi M., 2009. Performance of olive mill<br />
solid waste as a constituent of the substrate in commercial cultivation of Agaricus<br />
bisporus. International Biodeterioration and Biodegradation. 63: 993-997.<br />
Amir S., Hafidi M., Merlina G., Hamdi H., et Revel J. C., 2004. Elemental analysis, FTIR<br />
and C-13-NMR of humic acids from sewage sludge composting. Agronomie. 24: 13-<br />
18.<br />
Amirante P., Di Renzo G.C., Di Giovacchino L., Btanchiand B. et Catalano P., 1993.<br />
Evolution technologique des installations d'extraction de l'huile d'olives. Olivae. 48:<br />
43-53.<br />
Andreozzi R., 1998. Integrated of olive mill effluent (OME): study of ozonation coupled with<br />
anaerobic digestion. Water Resistance. 32: 2364 - 2367.<br />
Ayed L., Assas N., Sayadi S. et Hamdi M., 2005. Involvement of lignin peroxidase in the<br />
decolorization of black olive mill wastewaters by Geotrichum candidum. Letters in<br />
Applied Microbiology.40: 7-11.<br />
Azabou S., Najjar W., Gargoubi A., Ghorbel A. et Sayadi S., 2007. Catalytic wet peroxide<br />
photo-oxidation of phenolic olive oil mill wastewater contaminants: Part II.<br />
Degradation and detoxification of low-molecular mass phenolic compounds in model<br />
and real effluent. Applied Catalysis B: Environmental. 77: 166-174.<br />
Azbar N., Keskinet T., Eskinet T., et Catalkaya E.C., 2008. Improvement in anaerobic<br />
degradation of olive mill effluent (OME) by chemical pretreatment using batch<br />
systems. Biochemical Engineering Journal. 38: 379-383.<br />
Baçaoui A., Yaacoubi A., Dahbi A., Bennouna C., Phan Tan Luu R., Maldonado-Hodar<br />
F. J., Rivera-Utrilla J. et Moreno-Castilla C., 2001. Optimization of conditions for<br />
the preparation of activated carbons from olive-waste cakes. Carbon. 39: 425-432.<br />
Bagnon S.-M., 1999. Production de 6-pentil-α-pyrone par Trichoderma harzianum cultivé<br />
sur support solide. Thèse de Doctorat, Université de Montpellier II, 216 pages.<br />
Baldensperger J., Le Mer J., Hannibal L. et Quinto P. J., 1985. Solid state fermentation of<br />
banana wastes. Biotechnology Letters 7: 743-748.<br />
Balice V., Carrieri C., Cera O. et Rindon B., 1988. The fate of tannin-like compounds from<br />
olive mill effluents in biological treatment. in : Proceedings of the Fifth International<br />
Symposium: On Anaerobic Digestion. Hall E.R. and Hobson P.N. (eds.). Academic<br />
Press, Italie, 275-280.<br />
Barajas-Aceves M., 2000. Soil Ergosterol, Dimethyl Sulphide Reduction, and Microbial<br />
Biomass Along a Zn Concentrations Gradient in Soils from a Mine Spoil Tip.<br />
Bulletin of Environmental Contamination and Toxicology. 64: 786-793.<br />
Bärlocher F., Graça M.A.S., 2005. Total phenolics. In: Graça, M.A.S., Bärlocher, F.,<br />
Gessner, M.O., (Eds.), Methods to study litter decomposition, Springer,<br />
Berlin/NewYork, 97-100.<br />
Bednarski W., Adamczak M., Tomasik J. et Plaszczyk M., 2004. Application of oil<br />
refinery waste in the biosynthesis of glycolipids by yeast. Bioresource Technology.<br />
95: 15-18.<br />
Bellon-Maurel V., O. Orliac et P. Christen. 2003. Sensors and measurements in solid state<br />
fermentation: a review. Process Biochemistry. 38: 881-896.<br />
Ben Sassi A., Boularbah A., Jaouad A., Walker G. et Boussaid A., 2006. A comparison of<br />
Olive oil Mill Wastewaters (OMW) from three different processes in Morocco.<br />
Process Biochemistry. 41: 74-78.<br />
Benyahia N. et Zein K., 2003. Analyse des problèmes de l’industrie de l’huile d’olive et<br />
solutions récemment développées. Contribution spéciale de Sustainable Business<br />
Associates (Suisse) à SESEC II, 2-7.<br />
155
Références bibliographiques<br />
Bing U., Cini E., Cioni A. et Laurendi V., 1994. Smaltimento-recupero delle sanse<br />
de oliva proveniente da un due fasi mediante distribuzione in campo.<br />
Informatore Agrario. 47 : 75-78.<br />
Bouaziz M., Fki I., Jemai H., Ayadi M. et Sayadi S., 2008. Effect of storage on refined and<br />
husk olive oils composition: Stabilization by addition of natural antioxidants from<br />
Chemlali olive leaves. Food Chemistry. 108: 253-262.<br />
Boukhoubza F., Zenjari B., Hassani L., Loukouli I.L. et Nejmeddine A., 2007. Evaluation<br />
of the phenolic compounds toxicity in olive mill wastewater. Physical and Chemical<br />
News. 37: 133-140.<br />
Boyle D., 1998. Nutritional factors limiting the growth of Lentinula edodes and other whiterot<br />
fungi in wood. Soil Biology and Biochemistry. 30: 817-823.<br />
Casa R., D'Annibale A., Pieruccetti F., Stazi S.R., Sermanni G.G. et Lo Cascio B., 2003.<br />
Reduction of the phenolic components in olive-mill wastewater by an enzymatic<br />
treatment and its impact on durum wheat (Triticum durum Desf.) germinability.<br />
Chemosphere. 50: 959-66<br />
Cayuela M. L., Sánchez-Monedero M. A. et Roig A., 2006. Evaluation of two different<br />
aeration systems for composting two-phase olive mill wastes. Process Biochemistry.<br />
41: 616-623.<br />
Centi G., Perathoner S., Torre T. et Verduna M.G., 2000. Catalytic wet oxidation with<br />
H2O2 of carboxylic acids on homogeneous and heterogeneous Fenton type catalysts.<br />
Catalysis Today. 55: 61-69.<br />
Chalaux N., 1993. Approche expérimentale de la biodégradabilité des pailles de blé.<br />
Application à la quantification des matières premières et des composts pour la<br />
production de champignon de couche. Thèse de doctorat, Université de Bordeaux I,<br />
220 pages.<br />
Chamkha M., Patel B.K.C., Garcia J.L. et Labat M., 2001. Isolation of Clostridium<br />
biofermentants from oil mill wastewater converting cinnamic acid to 3phenylpropionic<br />
acid and emendation of species. Anaerobe. 7: 189-197.<br />
Chang S.T. 1999. Global impact of edible and Medicinal Mushrooms on human Welfare in<br />
the 21st century: Nongreen revolution. International Journal of Medicinal<br />
Mushrooms. 1: 1-7.<br />
Chen Y., Inbar Y., Hadar Y. et Malcolm R.L., 1989. Chemical properties and solid-state<br />
CPMAS 13C-NMR of composted organic matter. Science of Total Environment.<br />
81: 201-208.<br />
Chin Y. P., Alken G., et O'Loughlin E., 1994. Molecular weight, polydispersity, and<br />
spectroscopic properties of aquatic humic substances. Environmental Science and<br />
Technology. 28: 1853-1858.<br />
Chiofalo B., Liotta L., Zumbo A. et Chiofalo V., 2004. Administration of olive cake for<br />
ewe feeding: effect on milk yield and composition. Small Ruminant Research. 55:<br />
169-176.<br />
Chow T.T., 2009. A review on photovoltaic/thermal hybrid solar technology. Applied<br />
Energy. 87: 365-379.<br />
Claus H. et Filip Z., 1998. Degradation and transformation of aquatic humic substances by<br />
laccase-producing fungi Cladosporium cladosporioides and Polyporus versicolor.<br />
Acta hydrochimica et hydrobiologica. 26: 180-185.<br />
Claus H., Faber G. et König H., 2002. Redox-mediated decolorization of synthetic dyes by<br />
fungal laccases. Applied Microbiology and Biotechnology. 59: 672-678.<br />
Cohen R., Persky L. et Hadar Y., 2002. Biotechnological applications and potential of<br />
wood-degrading mushrooms of the genus Pleurotus. Applied Microbiology and<br />
Biotechnology. 58: 582-594.<br />
156
Références bibliographiques<br />
COI (Conseil Oléicole International), 2005. Amélioration de la qualité de l'huile d'olives.<br />
Madrid-Espagne, 10 pages.<br />
COI (Conseil Oléicole International), 2008. International course on water management and<br />
irrigation of olive orchards. Limassol-Cyprus, 20 pages.<br />
Colak N. et Hepbasli A., 2007. Performance analysis of drying of green olive in a tray dryer.<br />
Journal of Food Engineering. 80: 1188-1193.<br />
Collins P.J., Dobson A.D.W. et Field J.A., 1998. Reduction of the 2,2'-Azinobis(3-<br />
Ethylbenzthiazoline-6-Sulfonate) Cation Radical by Physiological Organic Acids in<br />
the Absence and Presence of Manganese. Applied and Environmental Microbiology.<br />
64: 2026-2031.<br />
Contreras-Dominiguez M., 2007. Dégradation des tanins condensés par des champignons<br />
filamenteux : analyse structurelle et enzymatique. Thèse de doctorat, Université de<br />
Provence, Université Aix-Marseille I, 161 pages.<br />
Cordova J., Nemmaoui M., Ismaïli-Alaoui M., Morin A., Roussos S., Raimbault M. et<br />
Benjilali B., 1998. Lipase production by solid state fermentation of olive cake and<br />
sugar cane bagasse. Journal of Molecular Catalysis B: Enzymatic. 5: 75-78.<br />
Cossu R., Blakey N., Cannas P., 1993. Influence of co-disposal of municipal solid waste and<br />
olive vegetation water on anaerobic digestion of sanitary landfill. Water Sciences<br />
Technology. 27: 261-271.<br />
Cossu R., Haarstad K., Lavagnolo M. C. et Littarru P., 2001. Removal of municipal solid<br />
waste COD and NH4-N by phyto-reduction: A laboratory-scale comparison of<br />
terrestrial and aquatic species at different organic loads. Ecological Engineering. 16:<br />
459-470.<br />
Couri S., Merces E. P., Neves B. C. V. et Senna L. F., 2006. Digital image processing as a<br />
tool to monitor biomass growth in Aspergillus niger 3T5B8 solid-state fermentation:<br />
preliminary results. Journal of Microscopy. 224: 290-297<br />
Criquet S., Tagger S., Vogt G., Iacazio G., Le Petit J., 1999. Laccase activity of forest<br />
litter. Soil Biology and Biochemistry. 31: 1239–1244.<br />
Crognale S., D’Annibale A., Federici F., Fenice M., Quaratino D. et Petruccioli M., 2006.<br />
Olive oil mill wastewater valorization by fungi. Journal of Chemical Technology and<br />
Biotechnology. 81: 1547-1555.<br />
Curvetto N.R., Figlas D., Devalis R. et Delmastro S., 2002a. Growth and productivity of<br />
different Pleurotus ostreatus strains on sunflower seed hulls supplemented with N-<br />
NH 4+ and/or Mn(II). Bioresource Technology. 84: 171-176.<br />
Curvetto N., Figlas D. et Delmastro S., 2002b, Sunflower seed hulls as substrate for the<br />
cultivation of shiitake mushrooms, Horticulture Technology. 12: 652–655.<br />
Czub J. et Baginski M., 2006. Comparative Molecular dynamics study of lipid membranes<br />
containing cholesterol and ergosterol. Biophysical Journal. 90: 2368-2382.<br />
D'Angelo S., Ingrosso D., Migliardi V., Sorrentino A., Donnarumma G., Baroni A.,<br />
Masella L., Antonietta Tufano M., Zappia M. et Galletti P., 2005.<br />
Hydroxytyrosol, a natural antioxidant from olive oil, prevents protein damage<br />
induced by long-wave ultraviolet radiation in melanoma cells. Free Radical Biology<br />
and Medicine. 38: 908-919.<br />
D'Annibale A., Casa R., Pieruccetti F., Ricci M. et Marabottini R., 2004. Lentinula<br />
edodes removes phenols from olive-mill wastewater: impact on durum wheat<br />
(Triticum durum Desf.) germinability. Chemosphere. 54: 887-894.<br />
D'Annibale A., Crestini C., Vinciguerra V. et Sermanni G.G., 1998. The biodegradation<br />
of recalcitrant effluents from an olive mill by a white-rot fungus. Journal of<br />
Biotechnology. 61: 209-218.<br />
157
Références bibliographiques<br />
D'Annibale A., Stazi S. R., Vinciguerra V. et Giovannozzi Sermanni G., 2000. Oxiraneimmobilized<br />
Lentinula edodes laccase: stability and phenolics removal efficiency in<br />
olive mill wastewater. Journal of Biotechnology. 77: 265-273.<br />
De Araujo A.A. et Roussos S., 2002. A technique for mycelial development of<br />
ectomycorrhizal fungi on agar media. Applied Biochemistry and Biotechnology. 98:<br />
311-318.<br />
De Araujo A.A., Aquiahual-Ramos M.A. et Roussos, S., 2000. Physiological studies on<br />
mycorrhizal fungi production. In: Sera, T., Soccol, C.R., Pandey, A., Roussos, S.,<br />
(Eds.), Coffee Biotechnology and Quality, Kluwer, Dordrecht, The Netherlands, pp.<br />
245-253.<br />
De Araujo A.A., Lepilleur C., Delcourt S., Colavitti P. et Roussos S., 1997. Laboratory<br />
scale bioreactors for study of fungal physiology and metabolism in solid state<br />
fermentation system. In Roussos, S., Lonsane, B.K., Raimbault, M. and Viniegra-<br />
Gonzalez, G. (Eds.), Advances in Solid State Fermentation, Kluwer Academy.<br />
Dordrecht, 93-111<br />
De Marco E., Savarese M., Paduano A. et Sacchi R., 2007. Characterisation and<br />
fractionation of phenolic compounds extracted from olive mill wastewaters. Food<br />
Chemistry. 104: 858–867.<br />
De Ridder-Duine A.S., Smant V., Van Der Wal A., Van Veen J.A. et De Boer W., 2006.<br />
Evaluation of a simple, non-alkaline extraction protocol to quantify soil ergosterol.<br />
Pedobiologia. 50: 293-300.<br />
Deschamps F., Raimbault M. et Senez J.C., 1982. Solid state fermentation in the<br />
development of agro-food by-products. Industry and Environment. 5: 27-30.<br />
Di Giovacchino L., Mascolo A., et Seghiti I., 1988. Sulle caratteristiche delle acque di<br />
vegetazione delle olive. Rivista Italana delle Sostanze Grasse. 65: 481-488.<br />
Di Serio M. G., Lanza B., Mucciarella M. R., Russi F., Iannucci E., Marfisi P. et Madeo<br />
A., 2008. Effects of olive mill wastewater spreading on the physico-chemical and<br />
microbiological characteristics of soil. International Biodeterioration and<br />
Biodegradation. 62: 403-407.<br />
DPV, direction de la production végétale, 2009, département lié au ministère de<br />
l'agriculture, Rabat, Maroc.<br />
Dubois M., Gilles K. A., Hamilton J. K., Rebers P. A. et Smith F., 1956.Colorimetric<br />
Method for Determination of Sugars and Related Substances. Analytical Chemistry.<br />
28: 350-356.<br />
Durán N. et Esposito E., 2000. Potential applications of oxidative enzymes and<br />
phenoloxidase-like compounds in wastewater and soil treatment. Applied Catalysis<br />
B-Environmental. 28: 83–99.<br />
Durand A. 2003. Bioreactor designs for solid state fermentation. Biochemical Engineering<br />
Journal. 13: 113-125.<br />
Eichlerova I., Homolka L. et Nerud F., 2006. Ability of industrial dyes decolorization and<br />
ligninolytic enzymes production by different Pleurotus species with special attention<br />
on Pleurotus calyptratus, strain CCBAS 461. Process Biochemistry. 41: 941-946.<br />
Ekechukwu O.V., 1999. Review of solar-energy drying systems I: an overview of drying<br />
principles and theory. Energy Conversion and Management. 40: 593-613.<br />
El Hajjouji H., Ait Baddi G., Yaacoubi A., Hamdi H., Winterton P., Revel J.C. et Hafidi<br />
M., 2008. Optimisation of biodegradation conditions for the treatment of olive mill<br />
wastewater. Bioresource Technology. 99: 5505-5510.<br />
El Hajjouji H., Fakharedine N., Baddi G.A., Winterton P., Bailly J.R., Revel J.C. et<br />
Hafidi M., 2007. Treatment of olive mill waste water by aerobic biodegradation: an<br />
158
Références bibliographiques<br />
analytical study using gel permeation chromatography, ultraviolet–visible and<br />
Fourier transform infrared spectroscopy. Bioresource Technology. 98: 3513-3520<br />
Elisashvili V., Penninckx M., Kachlishvili E., Tsiklauri N., Metreveli E., Kharziani T. et<br />
Kvesitadze G., 2008. Lentinus edodes and Pleurotus species lignocellulolytic<br />
enzymes activity in submerged and solid-state fermentation of lignocellulosic wastes<br />
of different composition. Bioresource Technology. 99: 457-462.<br />
Enguita F.J., Martins L.M., Henriques A.O. et Carrondo M.A., 2003. Crystal structure of<br />
a bacterial endospore coat component. A laccase with enhanced thermostability<br />
properties, Journal of Biological Chemistry. 278: 19416–19425.<br />
Erbay Z. et Icier F., 2009. Optimization of hot air drying of olive leaves using response<br />
surface methodology. Journal of Food Engineering. 91: 533-541.<br />
Ergül F.E., Sargin S., Ongen G. et Sukan F.V., 2009. Dephenolisation of olive mill<br />
wastewater using adapted Trametes versicolor. International Biodeterioration and<br />
Biodegradation. 63: 1-6.<br />
Eroglu E., Eroglu I., Gündüz U. et Yücel M., 2008. Effect of clay pretreatment on<br />
photofermentative hydrogen production from olive mill wastewater. Bioresource<br />
Technology. 99: 6799-6808.<br />
Farnet A.M., Criquet S., Cigna M., Gil G. et Ferré E., 2004. Purification of a laccase from<br />
Marasmius quercophilus induced with ferulic acid: reactivity towards natural and<br />
xenobiotic aromatic compounds. Enzyme and Microbial Technology. 34: 549-554.<br />
Farnet A.M., Prudent P., Ziarelli F., Domeizel M. et Gros R., 2009. Solid-state 13C NMR<br />
to assess organic matter transformation in a subsurface wetland under cheese-dairy<br />
farm effluents. Bioresource Technology. 100: 4899-4902.<br />
Fayed A.M., El-Ashry M.A. et Aziz H.A., 2009. Effect of Feeding Olive Tree Pruning By-<br />
Products on Sheep Performance in Sinai. World Journal of Agricultural Sciences. 5:<br />
436-445.<br />
Fenice M., Giovannozzi Sermanni G., Federici F. et D'Annibale A., 2003. Submerged and<br />
solid-state production of laccase and Mn-peroxidase by Panus tigrinus on olive mill<br />
wastewater-based media. Journal of Biotechnology. 100: 77-85.<br />
Fiestas J.A. et Borja R., 1992. Use and treatment of olive mill wastewater: Current situation<br />
and prospects in Spain. Grasas y Acietes. 43: 101-106.<br />
Fki I., Allouche N. et Sayadi S., 2005. The use of polyphenolic extract, purified<br />
hydroxytyrosol and 3,4-dihydroxyphenyl acetic acid from olive mill wastewater for<br />
the stabilization of refined oils: a potential alternative to synthetic antioxidants. Food<br />
Chemistry. 93: 197-204.<br />
Flouri F., Sotichos F. P., Ioannidou S. et balis C., 1997. Decoloration of olive mill liquid<br />
wastewater by chemical and biochemical means. International Biodeterioration and<br />
Biodegradation. 38: 189-192.<br />
Fountoulakis M.S., Dokianakis S.N., Kornaros M.E., Aggelis G.G. et Lyberatos, G.,<br />
2002. Removal of phenolics in olive mill wastewaters using the white-rot fungus<br />
Pleurotus ostreatus. Water Research. 36: 4735–4744.<br />
Foutlane A., Saadallah M., Echihabi L. et Bourchich, L., 2002. Pollution by wastewater<br />
from olive oil mills and drinking-water production. Case study of the Sebou river in<br />
Morocco. East Mediterranean Health Journal. 8: 129-140.<br />
Freire R.S., Durán N. et Kubota L.T., 2001. Effects of fungal laccase immobilization<br />
procedures for the development of a biosensor for phenol compounds. Talanta. 54:<br />
681-686.<br />
Garcia G. I., Jiménez Pena P.R., Bonilla Venceslada J.L., Martin Martin A., Martin<br />
Santos M.A. et Ramos Gomez E., 2000. Removal of phenol compounds from olive<br />
159
Références bibliographiques<br />
mill wastewater using Phanerochaete chrysosporium, Aspergillus niger, Aspergillus<br />
terreus and Geotrichum candidum. Process Biochemistry. 35: 751-758.<br />
Gervais P. et Bensoussan M., 1994. Solid-sate fermentation of the genus Aspergillus. In:<br />
Aspergillus, Smith, J. E. (Ed.). Plenum, Press, London, 101-140.<br />
Gessner M.O. et Chauvet E., 1993. Ergosterol-to-biomass conversion factors for aquatic<br />
hyphomycetes. Applied and Environmental Microbiology . 59: 502-507.<br />
Gilardi G., Abis L. et Cass, A.E.G., 1995. Carbon-13 CP/MAS solid-state NMR and FT-IR<br />
spectroscopy of wood cell wall biodegradation. Enzyme and Microbial Technology.<br />
17: 268-275.<br />
Gögüs, F. et Maskan M., 2006. Air drying characteristics of solid waste (pomace) of olive<br />
oil processing. Journal of Food Engineering.72: 378-382.<br />
Gong P., Guan X., et Witter E., 2001. A rapid method to extract ergosterol from soil by<br />
physical disruption. Applied Soil Ecology. 17: 285-289.<br />
Goujard L., Ferre E., Gil G., Ruaudel F. et Farnet A.M., 2009. A method to quantify<br />
transesterification activities of lipases in litters. Journal of Microbiological Methods.<br />
78: 127-130.<br />
Grabber J.H., 2009. Protein fractions in forage legumes containing protein-binding<br />
polyphenols: Freeze-drying vs. conservation as hay or silage. Animal Feed Science<br />
and Technology. 151: 324-329.<br />
Gu Y.H. et Belury M.A., 2005. Selective induction of apoptosis in murine skin carcinoma<br />
cells (CH72) by an ethanol extract of Lentinula edodes. Cancer Letters. 220: 21-28.<br />
Hadar Y. et Dosoretz C.G., 1991. Mushroom mycelium as a potential source of food<br />
flavour. Trends in Food Science and Technology. 2: 214-218.<br />
Haddadin M.S., Al-Natour R., Al-Qsous S. et Robinson R.K., 2002. Bio-degradation of<br />
lignin in olive pomace by freshly-isolated species of Basidiomycete. Bioresource<br />
Technology. 82: 131-137.<br />
Hama S., Yamaji H., Fukumizu T., Numata T., Tamalampudi S., Kondo A., Noda H. et<br />
Fukuda H., 2007. Biodiesel-fuel production in a packed-bed reactor using lipaseproducing<br />
Rhizopus oryzae cells immobilized within biomass support particles.<br />
Biochemical Engineering Journal. 34: 273-278.<br />
Hamden K., Allouche N., Damak M. et Elfeki A., 2009. Hypoglycemic and antioxidant<br />
effects of phenolic extracts and purified hydroxytyrosol from olive mill waste in<br />
vitro and in rats. Chemico-Biological Interactions. 180: 421-432.<br />
Hamdi M., 1992. Toxicity and biodegradability of olive millwastewater in batch anaerobic<br />
digestion. Applied. Biotechnology. 2: 155-163.<br />
Hamdi M., 1996. Anaerobic digestion of olive mill wastewaters. Process Biochemistry. 31:<br />
105-110.<br />
Hamdi M. et Ellouz R., 1993. Treatment of detoxified olive mill wastewaters by anaerobic<br />
filter and aerobic fluidized bed processes. Environmental Technology. 14: 183-188.<br />
Hassouni H., 2007. Physiologie de la sporulation des champignons filamenteux pour la<br />
production de spores et d'enzymes en fermentation en milieu solide. Thèse de<br />
doctorat, Institut Agronomique et Vétérinaire Hassan II, Rabat, 165 pages.<br />
Hattenschwiler S. et Vitousek P.M., 2000. The role of polyphenols in terrestrial ecosystem<br />
nutrient cycling. Trends in Ecology and Evolution. 15: 238-243.<br />
Hatvani N., 2001. Antibacterial effect of the culture fluid of Lentinus edodes mycelium<br />
grown in submerged liquid culture. International Journal of Antimicrobial Agents.<br />
17: 71-74.<br />
Haw J.F., Maciel G.E. et Schroeder, H.A., 1984. C-13 Nuclear Magnetic-Resonance<br />
Spectrometric Study of Wood and Wood Pulping with Cross Polarization and Magic-<br />
Angle Spinning. Analytical Chemistry. 56: 1323-1329.<br />
160
Références bibliographiques<br />
Hesseltine C.W., 1987. Solid state fermentation. An Overview. International<br />
Biodeterioration and Biodegradation. 23: 79-89.<br />
Hirasawa M., Shouji N., Neta T., Fukushima K. et Takada K., 1999. Three kinds of<br />
antibacterial substances from Lentinus edodes (Berk.) Sing. (Shiitake, an edible<br />
mushroom). International Journal of Antimicrobial Agents. 11: 151-157.<br />
Huang G.F., Wong J.W.C., Wu Q.T. et Nagar, B.B., 2004. Effect of C/N on composting of<br />
pig manure with sawdust. Waste Management. 24: 805-813.<br />
Inan I., Dimoglo A., Simsek H. et Karpuzcu M., 2004. Olive mill wastewater treatment by<br />
means of electro‑coagulation. Separation and Purification Technology. 36: 23-31.<br />
International Olive Oil Council, 2008. World olive oil figures, on line at<br />
http://www.internationaloliveoil.org/<br />
FODEP (Fonds de Dépollution Industrielle), 2004. Industrie oléicole au Maroc et son<br />
impact sur l’environnement: Propositions d’actions de lutte contre la pollution<br />
générée par les huileries d’olives, cas de la province de Taounate, 15 pages.<br />
http://www.environnement.gov.ma/fodep/pdf/SUMMARY.pdf<br />
Isidori M., Lavorgna M., Nardelli A. et Parrella A., 2005. Model study on the effect of 15<br />
phenolic olive mill wastewater constituents on seed germination and Vibrio fischeri<br />
metabolism. Journal of Agricultural and Food Chemistry. 53 : 8414-8417.<br />
Ismaili-Alaoui M. et Heddoun A., 2006. Tentative de modernisation des Maâsra<br />
traditionnelles. Unité mobile d’extraction des huiles d’olives. In : Ismaili-Alaoui,M.,<br />
Roussos, S., Perraud-Gaime, I., (Eds.), Biotechnology and quality of Olive tree<br />
products around the Mediterranean basin, Actes Editions, Rabat, Maroc, 243-258.<br />
Israilides C., Kletsas D., Arapoglou D., Philippoussis A., Pratsinis H., Ebringerova A.,<br />
Hribalova V. et Harding S.E., 2008. In vitro cytostatic and immunomodulatory<br />
properties of the medicinal mushroom Lentinula edodes. Phytomedicine. 15: 512-<br />
519.<br />
Ivanova D., Andonov K., 2001. Analytical and experimental study of combined fruit and<br />
vegetable dryer. Energy Conversion and Management. 42: 975-983.<br />
Jain D., et Tiwari G.N., 2004. Effect of greenhouse on crop drying under natural and forced<br />
convection I: Evaluation of convective mass transfer coefficient. Energy Conversion<br />
and Management. 45: 765-783.<br />
Janjai S., Lamlert N., Intawee P., Mahayothee B., Bala B.K., Nagle M. et Müller J.,<br />
2009. Experimental and simulated performance of a PV-ventilated solar greenhouse<br />
dryer for drying of peeled longan and banana. Solar Energy. 83: 1550-1565.<br />
Jaouani A., Sayadi S., Vanthournhout M. et Penninckx M.J., 2003. Potent fungi for<br />
decolourisation of olive oil mill wastewaters. Enzyme and Microbial Technology. 33:<br />
802-809.<br />
Jaouani A., Vanthournhout M. et Penninckx M.J., 2005. Olive oil mill wastewater<br />
purification by combination of coagulation-flocculation and biological treatments.<br />
Environmental Technology. 26: 633-641.<br />
Jasinghe V.J. et Perera C.O., 2005. Distribution of ergosterol in different tissues of<br />
mushrooms and its effect on the conversion of ergosterol to vitamin D2 by UV<br />
irradiation. Food Chemistry. 92: 541-546.<br />
Jasinghe V.J. et Perera C.O., 2006. Ultraviolet irradiation: The generator of Vitamin D2 in<br />
edible mushrooms. Food Chemistry. 95: 638-643.<br />
Jimenez S. et Lao M. T., 2004. Use of dry olive-mill wastewater like organic amendment in<br />
soil for horticultural greenhouse crop. Journal of Food Agriculture and Environment.<br />
2: 348-352.<br />
161
Références bibliographiques<br />
Jo J.H., Lee D.S., Park D. et Park J.M. 2008. Biological hydrogen production by<br />
immobilized cells of Clostridium tyrobutyricum JM1 isolated from a food waste<br />
treatment process. Bioresource Technology. 99: 6666-6672.<br />
Kabbaj W., 1997. Physiologie de la croissance et production d'arôme par des cultures<br />
mycéliennes de Pleurotus et de Morchella sur milieu solide. Thèse de doctorat,<br />
Université de Bourgogne, 165 pages.<br />
Kalberer P.P., 1995. An investigation of the incubation phase of a shiitake (Lentinus edodes)<br />
culture. Mushroom Science. 14: 375–383.<br />
Kalberer P.P., 2000. Influence of urea and ammonium chloride on crop yield and fruit body<br />
size of shiitake (Lentinula edodes). Mushroom Science. 15: 361–366.<br />
Kalmis E. et Sargin S., 2004. Cultivation of two Pleurotus species on wheat straw substrates<br />
containing olive mill waste water. International Biodeterioration and<br />
Biodegradation. 53: 43-47.<br />
Kalmis E., Azbar N., Yildiz H. et Kalyoncu F., 2008. Feasibility of using olive mill effluent<br />
(OME) as a wetting agent during the cultivation of oyster mushroom, Pleurotus<br />
ostreatus, on wheat straw. Bioresource Technology. 99: 164–169.<br />
Khoufi S., Aloui F. et Sayadi S., 2008. Extraction of antioxidants from olive mill wastewater<br />
and electrocoagulation of exhausted fraction to reduce its toxicity on anaerobic<br />
digestion. Journal of Hazardous Materials. 151: 531-539.<br />
Kitzberger C.S.G., Smânia J.A., Pedrosa R.C. et Ferreira S.R.S., 2007. Antioxidant and<br />
antimicrobial activities of shiitake (Lentinula edodes) extracts obtained by organic<br />
solvents and supercritical fluids. Journal of Food Engineering. 80: 631-638.<br />
Klamer M. et Bååth E., 2004. Estimation of conversion factors for fungal biomass<br />
determination in compost using ergosterol and PLFA 18:2Ω6,9. Soil Biology and<br />
Biochemistry. 36: 57-65.<br />
Kooli S., Fadhel A., Farhat A. et Belghith A., 2007. Drying of red pepper in open sun and<br />
greenhouse conditions.: Mathematical modeling and experimental validation.<br />
Journal of Food Engineering. 79: 1094-1103.<br />
Kuforiji O.O. et Fasidi I.O., 2009. Influence of light and spawn quantity on the growth of<br />
Nigerian mushroom Pleurotus tuber-regium. Journal of Environmental Biology. 30:<br />
605-608.<br />
Lakhtar H., Zaouia N., Salih G., Tantaoui-Elaraki A., Lamrani K., Cheheb M.,<br />
Hassouni H., Ismaili-Alaoui M., Verhé F., Gaime-Perraud I., Augur C. et<br />
Roussos S., 2006. Diversité des champignons filamenteux isolés dans les maâsra au<br />
Maroc. International Congress on Bioprocesses in Food Industries (ICBF-2006).<br />
Patras, Grèce.<br />
Lakhtar, H., Ismaili-Alaoui, M., Philippoussis A., Perraud-Gaime I., Roussos S., 2009.<br />
Screening of strains of Lentinula edodes grown on model olive mill wastewater in<br />
solid and liquid state culture for polyphenol biodegradation. International<br />
Biodeterioration and Biodegradation (in press) (DOI: 10.1016/j.ibiod.2009.10.006)<br />
Le Dréau Y., Dupuy N., Gaydou V., Joachim J. et Kister J., 2009. Study of jojoba oil<br />
aging by FTIR. Analytica Chimica Acta. 642: 163-170.<br />
Lechner B.E. et Papinutti V.L., 2006. Production of lignocellulosic enzymes during growth<br />
and fruiting of the edible fungus Lentinus tigrinus on wheat straw. Process<br />
Biochemistry. 41: 594-598.<br />
Lee O.H., Lee B.Y., Lee J., Lee H.B., Son J.Y., Park C.S., Shetty K. et Kim Y.C., 2009.<br />
Assessment of phenolics-enriched extract and fractions of olive leaves and their<br />
antioxidant activities. Bioresource Technology. 100: 6107-6113.<br />
Leontievsky A.A., Myasoedova N.M., Golovleva L.A., Sedarati M. et Evans C.S., 2002.<br />
Adaptation of the white-rot basidiomycete Panus tigrinus for transformation of high<br />
162
Références bibliographiques<br />
concentrations of chlorophenols. Applied Microbiology and Biotechnology. 59: 599-<br />
604.<br />
Levanon D., Rothschild N., Danai O. et Masaphy S., 1993. Strain selection for cultivation<br />
of Shiitake mushrooms (Lentinus edodes) on straw. Bioresource Technology. 45: 9-<br />
12.<br />
Lewis G.A., Mathieu D. et Phan-Tan-Luu R., 1999. Pharmaceutical Experimental Design.<br />
Marcel Dekker, New York.<br />
Lewis G.N., 1921. The rate of drying of solid Materials. Journal of Industrial Engineering<br />
and Chemistry. 13: 427-432.<br />
Liu Z. et Lee C., 2006. Drying effects on sorption capacity of coastal sediment: The<br />
importance of architecture and polarity of organic matter. Geochimica et<br />
Cosmochimica Acta. 70: 3313-3324.<br />
Longhi P., Vodopivec B. et Fiori G., 2001. Electrochemical treatment of olive oil mill<br />
wastewater. Annali di Chimica. 91: 169-174.<br />
Lonsane B.K., Ghildyal N.P., Budiatman S. et Ramakrishna S.V., 1985. Engineering<br />
aspects of solid state fermentation. Enzyme and Microbial Technology. 7: 258-265.<br />
Lonsane B.K., Saucedo-Castañeda G., Raimbault M., Roussos S., Viniegra-González G.,<br />
Ghildyal N.P., Ramakrishna M. et Krishnaiah M.M., 1992. Scale-up strategies<br />
for solid state fermentation systems. Process Biochemistry. 27: 259-273.<br />
Lopez-Villatta L.C., 1998. The Olive Tree, the Oil, the Olive (International Olive Oil<br />
Council Publications, Madrid).<br />
Mamiro D.P. et Royse D.J., 2008. The influence of spawn type and strain on yield, size and<br />
mushroom solids content of Agaricus bisporus produced on non-composted and<br />
spent mushroom compost. Bioresource Technology. 99: 3205-3212.<br />
Mandels M., Andreotti R, Roche C., 1976. Measurement of saccharifying cellulase.<br />
Biotechnology and Bioengineering Symposium. 6: 21-33.<br />
Manna C., Galletti P., Cucciolla V., Montedoro G. et Zappia V., 1999. Olive oil<br />
hydroxytyrosol protects human erythrocytes against oxidative damages. Journal of<br />
Nutritional Biochemistry. 10: 159-165.<br />
Mansur M., Arias M.E., Copa-Patino J.L., Flardh M. et Gonzalez A.E., 2003. The whiterot<br />
fungus Pleurotus ostreatus secretes laccase isozymes with different substrate<br />
specificities. Mycologia. 95: 1013-1020.<br />
Martin Garcia A.I., Moumen A., Yanez Ruiz D.R. et Molina Alcaide E., 2003. Chemical<br />
composition and nutrients availability for goats and sheep of two-stage olive cake<br />
and olive leaves. Animal Feed Science and Technology. 107: 61-74.<br />
Martínez-Carrera D., Guzmán G. et Soto C., 1985. The effect of fermentation of coffee<br />
pulp in the cultivation of Pleurotus ostreatus in Mexico. Mushroom Newsletter for<br />
the Tropics. 6: 21-28.<br />
Masghouni M., Hassairi M., 2000. Energy applications of olive-oil industry by products: the<br />
exhaust foot cake. Biomass and Bioenergy. 18: 257–262.<br />
Massiot D., Fayon F., Capron M., King I., Calvé S.L., Alonso B., Durand, J.O., Bujoli B.,<br />
Gan Z. et Hoatson G. 2002. Modelling one- and two-dimensional solid-state NMR<br />
spectra. Magnetic Resonance in Chemistry. 40: 70-76.<br />
Mata G. et Savoie J.M., 1998. Extracellular enzyme activities in six Lentinula edodes strains<br />
during cultivation in wheat straw. World Journal of Microbiology and<br />
Biotechnology. 4: 513–519.<br />
Mata M.G., 1997. Culture du champignon parfumé, shiitake (Lentinula edodes), sur paille de<br />
blé : étude sur les mécanismes d’adaptation nutritive et antagonisme avec<br />
Trichoderma sp. Thèse de doctorat, Institut national polytechnique de Toulouse, 168<br />
pages.<br />
163
Références bibliographiques<br />
Matcham S.E., Jordan B.R. et Wood D.A., 1985. Estimation of fungal biomass in a solid<br />
substrate by three independent methods. Applied Microbiology and Biotechnology.<br />
21: 108-112.<br />
Mathieu D. et Phan-Tan-Luu R., 1997. Plans d’expériences: Application à l’entreprise,<br />
Technipresse, Paris, 528.<br />
Mathieu D., Nony J. et Phan-Tan-Luu R., 2000. Nemrodw software, LPRAI, Marseille.<br />
Matos A.J.F.S., Bezerra R.M.F. et Dias A.A., 2007. Screening of fungal isolates and<br />
properties of Ganoderma applanatum intended for olive mill wastewater<br />
decolourization and dephenolization. Letters in Applied Microbiology. 45: 270–275.<br />
Mayer A.M. et Staples R.C., 2002. Laccase: new functions for an old enzyme.<br />
Phytochemistry. 60: 551-565.<br />
Mekki A., Dhouib A. et Sayadi, S., 2007. Polyphenols dynamics and phytotoxicity in a soil<br />
amended by olive mill wastewaters. Journal of Environmental Management. 84 :<br />
134-140.<br />
Mena E., Garrido A., Hernandez T. et Garcia C., 2003. Bioremediation of sewage sludge<br />
by composting. Communications in Soil Science and Plant Analysis. 34: 957-971.<br />
Mille-Lindblom C., Von Wachenfeldt E. et Tranvik L.J., 2004. Ergosterol as a measure of<br />
living fungal biomass: persistence in environmental samples after fungal death.<br />
Journal of Microbiological Methods. 59: 253-262.<br />
Miller G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugars.<br />
Analytical Chemistry. 31: 426-428.<br />
Mishra C. et Leatham G.F., 1990. Recovery and fractionation of the extracellular<br />
degradative enzymes from Lentinula edodes cultures cultivated on a solid<br />
lignocellulosic substrate. Journal of Fermentation and Bioengineering. 69: 8-15.<br />
Mitchell D.A., Krieger N., Stuart D.M. et Pandey A., 2000. New developments in solid<br />
state fermentation: II. Rational approaches to the design, operation and scale up of<br />
bioreactors. Process Biochemistry. 35: 1211-1225.<br />
Molina-Alcaide E., 2008. Effect of different drying procedures on the nutritive value of olive<br />
(Olea europaea var. europaea) leaves for ruminants. Animal Feed Science and<br />
Technology. 142: 317-329.<br />
Moo-Young M., Moreira A.R. et Tengerdy R.P., 1983. Principles of solid substrate<br />
fermentation. In: The Filamentous Fungi: Fungal Technology. Smith, J. E., Berry, D.<br />
R. et Kristiansen, B., (Eds.), Edward Arnold Publisher, London. 117-144.<br />
Morgan S.L., 2001. Calcium and Vitamin D in Osteoporosis. Rheumatic Disease Clinics of<br />
North America. 27: 101-130.<br />
Mouncif M., Tamoh S., Faid M. et Achkari B. A., 1993. A study of chemical and<br />
microbiological characteristics of olive mill wastewater in morocco. Grasas y<br />
Aceites. 44: 335-338.<br />
Müller J., Reisinger G., Kisgeci J., Kotta E., Tesic M. et Mühlbauer W., 1989.<br />
Development of a greenhouse-type solar dryer for medicinal plants and herbs. Solar<br />
and Wind Technology. 6: 523-530.<br />
Munoz, C., Guillen F., Martinez A.T., Martinez M.J., 1997. Induction and characterization<br />
of laccase in the ligninolytic fungus Pleurotus eryngii. Current Microbiology. 34: 1-<br />
5.<br />
Narahara H., Koyama Y., Yoshida T., Pichangkura S., Ueda R. et Taguchi H., 1982.<br />
Growth and enzyme production in a solid-state culture of Aspergillus oryzae. Journal<br />
of Fermention and Technology. 60: 311-319.<br />
Nefzaoui A., 1995. Feeding value of Mediterranean ruminant feed resources. Advanced<br />
course. Syria 12-23 March 1995.<br />
164
Références bibliographiques<br />
Newman R. H. et Hemmingson J.A., 1990. Determination of the Degree of Cellulose<br />
Crystallinity in Wood by Carbon-13 Nuclear Magnetic Resonance Spectroscopy.<br />
Holzforschung. 44: 351-356.<br />
Niemenmaa O., Galkin S. et Hatakka A., 2008. Ergosterol contents of some wood-rotting<br />
basidiomycete fungi grown in liquid and solid culture conditions. International<br />
Biodeterioration and Biodegradation. 62: 125-134.<br />
Ohga S., 1990. Growth rate of mycelium of shiitake, Lentinus edodes, in relation to water<br />
potential of medium. Journal of the Faculty of Agriculture, Kyushu University. 34:<br />
413–420.<br />
Olivieri G., Marzocchella A., Salatino P., Giardina P., Cennamo G. et Sannia G., 2006.<br />
Olive mill wastewater remediation by means of Pleurotus ostreatus. Biochemical<br />
Engineering Journal. 31: 180-187.<br />
Ooijkaas L.P., Tramper J. et Buitelaar R.M., 1998. Biomass Estimation of Coniothyrium<br />
Minitans in Solid-State Fermentation. Enzyme and Microbial Technology. 22: 480-<br />
486.<br />
Oriol E., 1987. Croissance d’Aspergillus niger sur milieu solide: Importance de l'eau et de<br />
l'activité de l'eau. Thèse de Doctorat, Institut national des sciences appliquées,<br />
Toulouse, 115 pages.<br />
Otjen L., Blanchette R. et Effland M., Assessment of 30 white-rot basidiomycetes for<br />
selective lignin degradation. Holzforschung. 41: 343–349.<br />
Özçelik E. et Peksen A., 2007. Hazelnut husk as a substrate for the cultivation of shiitake<br />
mushroom (Lentinula edodes). Bioresource Technology. 98: 2652-2658.<br />
Pandey A., Soccol C.R. et Mitchell D., 2000. New developments in solid state fermentation:<br />
I. Bioprocesses and products. Process Biochemistry. 35: 1153-1169.<br />
Paredes C., Cegarra J., Bernal M.P. et Roig A., 2005. Influence of olive mill wastewater in<br />
composting and impact of the compost on a Swiss chard crop and soil properties.<br />
Environment International. 31: 305-312.<br />
Pasanen A.L., Yli-Pietila K., Pasanen P., Kalliokoski P. et Tarhanen J., 1999. Ergosterol<br />
content in various fungal species and biocontaminated building materials. Applied<br />
and Environmental Microbiology. 65: 138-142.<br />
Perraud-Gaime I., 1995. Cultures mixtes en milieu solide de bactéries lactiques et de<br />
champignons filamenteux pour la conservation et la décaféination de la pulpe de<br />
café. Thèse de doctorat, Université de Montpellier II, 210 pages.<br />
Philippoussis A., 2009. Production of mushrooms using agro-industrial residues as<br />
substrates. In: Sing N.P., Pandey A., (Eds), Biotechnology for Agro-industrial<br />
Residues Processing, Springer, pp. 163-196.<br />
Philippoussis A., Diamantopoulou P. et Israilides C., 2007. Productivity of agricultural<br />
residues used for the cultivation of the medicinal fungus Lentinula edodes.<br />
International Biodeterioration and Biodegradation. 59: 216-219.<br />
Philippoussis A., Zervakis G. et Diamantopoulou P., 2001. Bioconversion of agricultural<br />
lignocellulosic wastes through the cultivation of the edible mushrooms Agrocybe<br />
aegerita, Volvariella volvacea and Pleurotus spp. World Journal of Microbiology<br />
and Biotechnology. 17: 191-200.<br />
Philippoussis A., Zervakis G., Diamantopoulou P. et Ioannidou S., 2000 Potential for the<br />
cultivation of exotic mushroom species by exploitation of Mediterranean agricultural<br />
wastes. Mushroom Science. 15: 523–530.<br />
Philippoussis A.N., Diamantopoulou P. et Zervakis G.I., 2003. Correlation of the<br />
properties of several lignocellulosic substrates to the crop performance of the shiitake<br />
mushroom Lentinula edodes. World Journal of Microbiology and Biotechnology. 19:<br />
551-557.<br />
165
Références bibliographiques<br />
Pietro C. Nazzaro N. and Seba C., 1988. Olive mill wastewater treatement by combined<br />
membrane processus. Water Resistance. 22: 1491-1494.<br />
Pintado J., Lonsane B. K., Gaime-Perraud I. et Roussos S., 1998. On-line monitoring of<br />
citric acid production in solid-state culture by respirometry. Process Biochemistry.<br />
33: 513-518.<br />
Purama R. K. et Goyal A., 2008. Screening and optimization of nutritional factors for higher<br />
dextransucrase production by Leuconostoc mesenteroides NRRL B-640 using<br />
statistical approach. Bioresource Technology. 99: 7108-7114.<br />
Raimbault M. et Alazard D., 1980. Culture method to study fungal growth in solid<br />
fermentation. European Journal of Applied Microbiology and Biotechnology. 9: 199-<br />
209.<br />
Raimbault M., 1980. Solid Fermentation in Bioconversion of Agro-Industrial Raw Materials,<br />
Documents ORSTOM, Paris, 147 pages.<br />
Ramos-Cormenzana A., Jirez-Jiménez B. et Garcia-Pareja M.P., 1996. Antimicrobial<br />
activity of olive mill wastewaters (alpechin) and biotransformed olive oil mill<br />
wastewater. International Biodeterioration et Biodegradation. 38: 283-290.<br />
Ranalli A., 1991. L’effluent des huiles d’olives : propositions en vue de son utilisation et son<br />
épuration. Références aux normes italiennes en la matière. Olivae. 39: 18-34.<br />
Ranz A. et Lankmayr E., 2006. Screening and optimization of the derivatization of polar<br />
herbicides with trimethylanilinium hydroxide for GC-MS analysis. Journal of<br />
Biochemical and Biophysical Methods. 69: 3-14.<br />
Rao J.R., Millar B.C. et Moore J.E., 2009. Antimicrobial properties of shiitake mushrooms<br />
(Lentinula edodes). International Journal of Antimicrobial Agents. 33: 591-592.<br />
Rio J.C., Gutierrez A., Martinez M.J. et Martinez A.T., 2001. Py-GC/MS study of<br />
Eucalyptus globulus wood treated with different fungi. Journal of Analytical and<br />
Applied Pyrolysis. 58: 441-452.<br />
Rio J.C., Speranza M., Gutierrez A., Martinez M.J. et Martinez A.T., 2002. Lignin attack<br />
during eucalypt wood decay by selected basidiomycetes: a Py-GC/MS study. Journal<br />
of Analytical and Applied Pyrolysis. 64: 421-431.<br />
Rosario L., Ana R., Bernardo M., Gerardo A.C. et Antonio G., 1999. Biodegradation of<br />
olive mill wastewaters (OMW) by strains of Scopulariopsis spp. isolated from<br />
OMW-contaminated habitats. Toxicology and Environmental Chemistry. 72: 127-<br />
134.<br />
Roussos S., 1985. Croissance de Trichoderma harzianum par fermentation en milieu solide:<br />
physiologie, sporulation, et production de cellulases. Thèse de Doctorat, Université<br />
de Provence, Aix-Marseille I, 193 pages.<br />
Roussos S. et Raimbault M., 1982. Hydrolysis of cellulose by fungi. 1. Screening of<br />
cellulolytic strains. Annal Microbiology. 133: 455-464.<br />
Roussos S., Aquiáhualt M.A., Brizuela M.A., Olmos A., Rodríguez W. et Viniegra-<br />
González G., 1989. Production, conservation and viability of filamentous fungi<br />
inoculum for solide substrate fermentation. Micologia. Neotropical Aplicada. 2: 3-<br />
17.<br />
Roussos S., Bresson E., Saucedo-Castañeda G., Martínez P., Olivier J.M. et<br />
Guimberteau J., 1997. Production of mycelial cell inoculum of Pleurotus opuntiae<br />
on natural support in solid state fermentation. In: Advances in Solid State<br />
Fermentation. Roussos, S., Lonsane, B. K., Raimbault, M. and Viniegra-González,<br />
G. (Eds.), Kluwer Academic Publishers, Dordrecht, Netherlands, 481-502.<br />
Roussos S., Zaouia N., Salih G., Tantaoui-Elaraki A., Lamrani K., Cheheb M., Hassouni<br />
H., Verhe F., Perraud-Gaime I., Augur C. et Ismaili-Alaoui M., 2006.<br />
Characterization of filamentous fungi isolated from olive and olive cake:<br />
166
Références bibliographiques<br />
Toxinogenic potential of Aspergillus strains. Molecular Nutrition and Food<br />
Research. 50: 517-523<br />
Royse D.J., 2001. Cultivation of shiitake on synthetic and natural logs. College of<br />
Agricultural Sciences, Cooperative Extension, Pennsylvania State University,<br />
University Park, USA, 12 pages.<br />
Royse, D.J. et Chalupa W., 2009. Effects of spawn, supplement and phase II compost<br />
additions and time of re-casing second break compost on mushroom (Agaricus<br />
bisporus) yield and biological efficiency. Bioresource Technology. 100: 5277-5282.<br />
Royse D.J. et Sanchez-Vazquez J.E., 2001. Influence of substrate wood-chip particle size on<br />
shiitake (Lentinula edodes) yield. Bioresource Technology. 76: 229-233.<br />
Royse D.J. et Sanchez-Vazquez J.E., 2003. Influence of precipitated calcium carbonate<br />
(CaCO3) on shiitake (Lentinula edodes) yield and mushroom size. Bioresource<br />
Technology. 90: 225-228.<br />
Royse D.J. et Sanchez-Vazquez J.E., 2007. Ground wheat straw as a substitute for portions<br />
of oak wood chips used in shiitake (Lentinula edodes) substrate formulae.<br />
Bioresource Technology. 98: 2137-2141.<br />
Saadi I., Laor Y., Raviv M. et Medina S., 2007. Land spreading of olive mill wastewater:<br />
effects on soil microbial activity and potential phytotoxicity. Chemosphere. 66: 75–<br />
83.<br />
Sampedro I., Marinari S., D'Annibale A., Grego S., Ocampo J. A. et Garça-Romera, I.,<br />
2007. Organic matter evolution and partial detoxification in two-phase olive mill<br />
waste colonized by white-rot fungi. International Biodeterioration and<br />
Biodegradation. 60: 116-125.<br />
Santos V.L. et Linardi, V.R., 2004. Biodegradation of phenol by a filamentous fungi<br />
isolated from industrial effluents--identification and degradation potential. Process<br />
Biochemistry. 39: 1001-1006.<br />
Saucedo-Castañeda G., 1995. Contrôle du métabolisme de Schwanniomyces castelli cultivé<br />
sur support solide. Thèse de Doctorat, Université Montpellier II, 212 pages.<br />
Savoie J.M. et Mata G., 1999. The antagonistic action of Trichoderma sp. hyphae to<br />
Lentinula edodes hyphae changes lignocellulolytic activities during cultivation in<br />
wheat straw. World Journal of Microbiology and Biotechnology. 15: 369-373<br />
Savoie J.M. et Mata G., 2003. Trichoderma harzianum metabolites pre-adapt mushrooms to<br />
Trichoderma aggressivum antagonism. Mycologia. 95: 191-199.<br />
Savoie J.M., Mata G. et Mamoun M., 2001. Variability in brown line formation and<br />
extracellular laccase production during interaction between white-rot basidiomycetes<br />
and Trichoderma harzianum biotype Th2. Mycologia. 93: 243-248.<br />
Sayadi S. et Ellouz R., 1993. Screening of white rot fungi for the treatment of olive mill<br />
waste-water. Journal of Chemical Technology and Biotechnology. 57: 141–146.<br />
Sayadi S. et Ellouz R., 1995. Role of lignin peroxidase and manganese peroxidase from<br />
Phanerochaete chrysosporium in the decolourization of olive mill waste-water,<br />
Applied and Environmental Microbiology. 61: 1098–1103.<br />
Sayadi S., Allouche N., Jaoua M. et Aloui F., 2000. Detrimental effects high molecularmasse<br />
polyphenols on olive mill wastewater biotreatment. Process Biochemistry. 35:<br />
725-735.<br />
Schunemann G., 1988. Biotechnological investigations on the possibilities of intensive<br />
culture of shiitake (Lentinus edodes) (Berk.) Sing. Horticulture Abstracts. 59: 7-182 .<br />
Scotti C.T., Vergoignan C., Feron G. et Durand A., 2001. Glucosamine measurement as<br />
indirect method for biomass estimation of Cunninghamella elegans grown in solid<br />
state cultivation conditions. Biochemical Engineering Journal. 7: 1-5.<br />
167
Références bibliographiques<br />
Sebabi Y., Dheilly R.M. et Queneudec M., 2003. Study of the water-retention capacity of a<br />
lime-sand mortar: Influence of the physicochemical characteristics of the lime.<br />
Cement and Concrete Research. 33: 689-696.<br />
Seitz L.M., Mohr H.M., Burroughs R. et Sauer D.B., 1977. Ergosterol as indicator of<br />
fungal invasion in grains. Cereal Chemistry. 54:. 1207–1217.<br />
Serghini H., Arrach R. et Aw-Hassan A., 2007. http://www.vulgarisation.net/agdumed2009<br />
/Serghini_Olive_Value_Chain_Analysis_Haouz_Area.pdf<br />
Sharma S., Bhat T.K. et Dawra R.K., 2000. A Spectrophotometric method for assay of<br />
tannase using rhodanine. Analytical Biochemistry. 279: 85-89.<br />
Shen J., Tanida M., Fujisaki Y., Horii Y., Hashimoto K. et Nagai K., 2009. Effect of the<br />
culture extract of Lentinus edodes mycelia on splenic sympathetic activity and cancer<br />
cell proliferation. Autonomic Neuroscience. 145: 50-54.<br />
Shen Q., Liu P., Wang X. et Royse D.J., 2008. Effects of substrate moisture content, log<br />
weight and filter porosity on shiitake (Lentinula edodes) yield. Bioresource<br />
Technology. 99: 8212-8216.<br />
Sherwood T.K., 1929. The drying of Solid. Journal of Engering and chemistry. 21: 976-980.<br />
Shicheng C., Dengbo M., Wei G., John A.B., 2003, Induction of laccase activity in the<br />
edible straw mushroom Volvariella volvacea. FEMS Microbiology Letters. 218: 143-<br />
148.<br />
Silva E.M., Machuca A. et Milagres A.M.F., 2005. Effect of cereal brans on Lentinula<br />
edodes growth and enzyme activities during cultivation on forestry waste. Letters in<br />
Applied Microbiology. 40: 283-288.<br />
Silva E.M., Martins S.F. et Milagres A.M.F., 2008. Extraction of manganese peroxidase<br />
produced by Lentinula edodes. Bioresource Technology. 99: 2471-2475.<br />
Singhania R.R., Patel A.K., Soccol C.R. et Pandey A., 2009. Recent advances in solid-state<br />
fermentation. Biochemical Engineering Journal. 44: 13-18.<br />
Singhania R.R., Soccol C.R. et Pandey A., 2007. Application of tropical agro-industrial<br />
residues as substrates for solid-state fermentation processes. In: Pandey A., Soccol<br />
C.R. and Larroche C., Editors, Current Developments in Solid-state Fermentation,<br />
Springer Science/Asiatech Publishers, Inc., New York, USA/New Delhi, India, 412–<br />
442.<br />
Sobal M., 2002. Physiologie de la croissance mycélienne de Pleurotus ostreatus et<br />
dégradation des polyphénols de la pulpe et des coques de café. Thèse de doctorat,<br />
Université de Provence, Aix-Marseille I, 207 pages.<br />
Sobal M., Martínez-Carrera D., Rio B., Roussos S., 2002. Screening of edible mushrooms<br />
for polyphenol degradation and tannase production from coffee pulp and coffee husk.<br />
In: Roussos, S., Soccol C.R., Pandey A., Augur C., (Eds.), New Horizons in<br />
Biotechnology, Kluwer, Dordrecht, Netherlands, 89-95<br />
Sukumaran R.K., Singhania R.R. et Pandey A., 2005. Microbial cellulases production,<br />
applications and challenges. Journal of Scientific and Industrial Research. 64: 832-<br />
844.<br />
Sunchez J.E. et Royse, D.J., 2001. Adapting substrate formulas used for shiitake for<br />
production of brown Agaricus bisporus. Bioresource Technology. 77: 65-69.<br />
Swalley S.E., Fulghum, J.R. et Chambers S.P., 2006. Screening factors effecting a response<br />
in soluble protein expression: Formalized approach using design of experiments.<br />
Analytical Biochemistry. 351: 122-127.<br />
Teichmann A., Dutta P.C., Staffas A. et Jägerstad M., 2007. Sterol and vitamin D2<br />
concentrations in cultivated and wild grown mushrooms: Effects of UV irradiation.<br />
LWT-Food Science and Technology. 40: 815-822.<br />
168
Références bibliographiques<br />
Terashima Y., Igusa H. et Ohga S., 2002. Influence of contamination by Penicillium<br />
brevicompactum and Trichoderma harzianum during Lentinula edodes spawn run on<br />
fruiting in sawdust-based substrates. Mycoscience. 43: 277-280.<br />
Tomati U., Belardinelli M., Galli E., Iori V., Capitani D., Mannina L., Viel S., et Segre<br />
A., 2004. NMR characterization of the polysaccharidic fraction from Lentinula<br />
edodes grown on olive mill waste waters. Carbohydrate Research. 339: 1129-1134.<br />
Trejo-Hernandez M.P., 1992. Physiologie de croissance de souches de Claviceps:<br />
Production d’alcaloides par fermentation en milieu solide. Thèse de Doctorat,<br />
Université de Provence, Aix-Marseille I, 164 pages.<br />
Trejo-Hernandez M.R., Lopez-Munguia A. et Quintero Ramirez R., 2001. Residual<br />
compost of Agaricus bisporus as a source of crude laccase for enzymic oxidation of<br />
phenolic compounds. Process Biochemistry. 36: 635-639.<br />
Tsai S.Y., Huang S.J., Lo S.H., Wu T.P., Lian P.Y. et Mau J.L., 2009. Flavour<br />
components and antioxidant properties of several cultivated mushrooms. Food<br />
Chemistry. 113: 578-584.<br />
Tsioulpas A., Dimou D., Iconomou D. et Aggelis G., 2002. Phenolic removal in olive oil<br />
mill wastewater by strains of Pleurotus spp. in respect to their phenol oxidase<br />
(laccase) activity. Bioresource Technology. 84: 251–257.<br />
Van Lier J.J.C., Van Ginkel J.T., Straatsma G., Gerrits J.P.G. et Van Griensven<br />
L.J.L.D., 1994, Composting of mushroom substrate in a fermentation tunnel:<br />
compost parameters and a mathematical model. Netherlands Journal of Agricultural<br />
Science. 42: 271–292.<br />
Vane C.H., Drage T.C. et Snape C.E., 2006. Bark decay by the white-rot fungus Lentinula<br />
edodes: Polysaccharide loss, lignin resistance and the unmasking of suberin.<br />
International Biodeterioration and Biodegradation. 57: 14-23.<br />
Velazquez-Cedeño M., Farnet A.M., Mata G. et Savoie J.M., 2008. Role of Bacillus spp.<br />
in antagonism between Pleurotus ostreatus and Trichoderma harzianum in heattreated<br />
wheat-straw substrates. Bioresource Technology. 99: 6966-6973.<br />
Velazquez-Cedeño, M. A., 2005. Compétition entre Pleurotus ostreatus et Trichoderma sp.<br />
en culture sur paille de blé: rôle des communautés bactériennes du substrat et des<br />
laccases de Pleurotus. Thèse de doctorat. Université Paul Cézanne, Aix-Marseille <strong>III</strong>,<br />
165 pages.<br />
Wariishi H., Valli K. et Gold M.H., 1992. Manganese oxidation by manganese peroxidase<br />
from the basidiomycete Phanerochaete chrysosporium, Journal of Biological<br />
Chemistry.267: 23688–23695.<br />
Weinberg Z.G., Chen Y., Weinberg P., 2008. Ensiling olive cake with and without<br />
molasses for ruminant feeding. Bioresource Technology. 99: 1526-1529.<br />
Widsten P. et Kandelbauer A., 2008. Laccase applications in the forest products industry: A<br />
review. Enzyme and Microbial Technology. 42: 293-307.<br />
Wikberg H. et Maunu L.S., 2004. Characterisation of thermally modified hard and<br />
softwoods by 13 C CPMAS NMR. Carbohydrate Polymers. 58: 461-466.<br />
Wu K.J., Chang C.F. et Chang J.S., 2007. Simultaneous production of biohydrogen and<br />
bioethanol with fluidized-bed and packed-bed bioreactors containing immobilized<br />
anaerobic sludge. Process Biochemistry. 42: 1165-1171.<br />
Yuan J.P., Wang J.H., Liu X., Kuang H.C. et Zhao S.Y., 2007. Simultaneous<br />
determination of free ergosterol and ergosteryl esters in Cordyceps sinensis by<br />
HPLC. Food Chemistry. 105: 1755-1759.<br />
Zenjari B., El Hajjouji H., Baddi G.A., Bailly J.R., Revel J.C., Nejmeddine A. et Hafidi,<br />
M., 2006. Eliminating toxic compounds by composting olive mill wastewater-straw<br />
mixtures. Journal of Hazardous Materials. 138: 433-437.<br />
169
Références bibliographiques<br />
Zervakis G., Yiatras P. et Balis C., 1996. Edible mushrooms from olive oil mill wastes.<br />
International Biodeterioration and Biodegradation. 38: 237-243.<br />
Zhang C.K., Gong F. et Li D.S., 1995. A note on the utilisation of spent mushroom<br />
composts in animal feeds. Bioresource Technology. 52: 89-91.<br />
Zhang R., Li X. et Fadel J.G. 2002. Oyster mushroom cultivation with rice and wheat straw.<br />
Bioresource Technology. 82: 277-284.<br />
Zouari-Mechichi H., Mechichi T., Dhouib A., Sayadi S., Martinez A.T. et Martinez M.J.,<br />
2006. Laccase purification and characterization from Trametes trogii isolated in<br />
Tunisia : decolorization of textile dyes by the purified enzyme. Enzyme Microbiology<br />
and Technology. 39: 141-148.<br />
170
Culture du Lentinula edodes (Berk.) Pegler sur résidus oléicoles en fermentation en<br />
milieu solide : Transformation des polyphénols des margines<br />
Résume :<br />
L’objectif du travail a été d’étudier la valorisation de sous-produits oléicoles au Maroc par la culture<br />
en fermentation en milieu solide de champignons comestibles avec une double finalité: (i) la<br />
production de la biomasse fongique pour l’alimentation humaine et (ii) la détoxification des margines<br />
par la dégradation des composés phénoliques. A partir de seize souches de L. edodes, une souche<br />
performante à la détoxification des margines a été sélectionnée pour mener la culture de L. edodes. La<br />
production de l’inoculum sur les grains de blé (spawn) a été optimisée. Un mélange des substrats<br />
oléicoles solides (SOS), composé de bois de taille d’olivier, de grignons d’olive et de margines, a été<br />
mis au point et les conditions de croissance de L. edodes sur ce mélange ont été optimisées. Le<br />
changement de la matière organique durant le développement de L. edodes sur le mélange dans des<br />
conditions optimales a été analysé par les techniques spectroscopiques de la RMN solide du 13 C<br />
couplées aux analyses biochimiques. Une technique de séchage solaire sous serre a été mise au point<br />
permettant la réduction en volume et le conditionnement des grignons d’olives mélangés aux<br />
margines. De ce travail, nous concluons la possibilité de valoriser l’ensemble des co-produits de<br />
l’industrie oléicole par la culture de L. edodes permettant, d’une part, la production d’une biomasse<br />
mycélienne importante et, d’autre part, la dégradation des composés phénoliques présents dans les<br />
margines. Ces résultats novateurs ouvrent des perspectives sérieuses pour la production de L. edodes<br />
sur le SOS.<br />
Mots clés : Industrie oléicole, Bois de taille, Grignons d’olive, Margines, Lentinula edodes,<br />
Fermentation en milieu solide, Enzymes<br />
Culture of Lentinula edodes (Berk.) Pegler on olive résidues in solid state fermentation:<br />
Transformation of polyphenols existing in olive mill wastewater<br />
Abstract :<br />
The main objective of this research was to study the recovery of olive by-products in Morocco by<br />
culturing, in solid state fermentation (SSF), of edible mushroom having double purpose: (i) production<br />
of fungal mycelial biomass for food and (ii) detoxification of olive mill wastewater (OMW) by<br />
degradation of phenolic compounds. From sixteen strains of L. edodes, one efficient strain for OMW<br />
detoxification was selected to perform further experimental work in SSF. The production of L. edodes<br />
spawn was optimized. Mixture of solid olive substrate (SOS), including olive twigs, leaves, olive cake<br />
and OMW, was developed and the growth conditions of L. edodes on this mixture was then optimized.<br />
The change in organic matter of SOS during L. edodes growth under optimal conditions was analyzed<br />
using solid state 13C NMR technique combined to biochemical analysis. A solar drying in greenhouse<br />
was developed in order to reduce the volume of OMW and to store the OC mixed to OMW. In this<br />
work, we conclude the possibility to valorize all co-products of olive industry by growing L. edodes,<br />
firstly, for production of high mycelial biomass, on the other hand, degradation of phenolic<br />
compounds existing in OMW. These results open up the possibility to produce L. edodes on SOS.<br />
Key words: Olive industry, Olive tree pruning, Olive cake, Olive mill wastewater, Lentinula edodes,<br />
Solid state fermentation, Enzymes<br />
Discipline: Biologie des populations et Ecologie<br />
U.F.R Faculté des Sciences et Techniques, <strong>IMEP</strong> Equipe d’Ecologie Microbienne, CNRS UMR 6116,<br />
Service 441, Avenue Escadrille Normandie-Niémen F13397, Marseille cedex 20