

guias de trabajos practicos y talleres - Facultad de Medicina
guias de trabajos practicos y talleres - Facultad de Medicina
guias de trabajos practicos y talleres - Facultad de Medicina

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
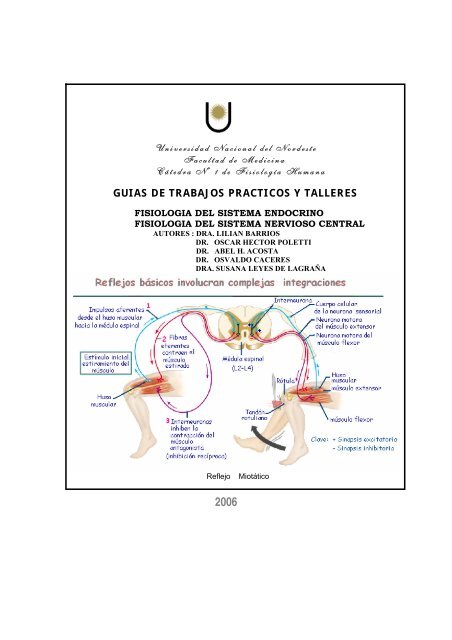
Universidad Nacional <strong>de</strong>l Nor<strong>de</strong>ste<strong>Facultad</strong> <strong>de</strong> <strong>Medicina</strong>Cátedra Nº 1 <strong>de</strong> Fisiología HumanaGUIAS DE TRABAJOS PRACTICOS Y TALLERESFISIOLOGIA DEL SISTEMA ENDOCRINOFISIOLOGIA DEL SISTEMA NERVIOSO CENTRALAUTORES : DRA. LILIAN BARRIOSDR. OSCAR HECTOR POLETTIDR. ABEL H. ACOSTADR. OSVALDO CACERESDRA. SUSANA LEYES DE LAGRAÑAReflejoMiotático2006
1Universidad Nacional <strong>de</strong>l Nor<strong>de</strong>ste<strong>Facultad</strong> <strong>de</strong> <strong>Medicina</strong>Cátedra Nº 1 <strong>de</strong> Fisiología HumanaGUIAS DE TRABAJOS PRACTICOS Y TALLERES(Correspondientes al Cuarto Examen Parcial)FISIOLOGIA DEL SISTEMA ENDOCRINO:Regulación <strong>de</strong> la glucemiaCorrelación endocrina en la mujerCorrelación endocrina en el hombreFISIOLOGIA DEL SISTEMA NERVIOSO CENTRALFisiología <strong>de</strong> los reflejosFisiología <strong>de</strong>l cerebeloRegulación <strong>de</strong> la postura2006Editor: Centro <strong>de</strong> Fotocopiado <strong>de</strong> la <strong>Facultad</strong> <strong>de</strong> <strong>Medicina</strong> <strong>de</strong> la Universidad Nacional <strong>de</strong>l Nor<strong>de</strong>ste
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 2REGULACION DE LA GLUCEMIA(Dres.: Abel ACOSTA y Osvaldo CACERES)OBJETIVOS: al finalizar el seminario elalumno <strong>de</strong>berá estar capacitado para:- Señalar la importancia <strong>de</strong> mantener una glucemia normal- Indicar las hormonas hipo e hiperglucemiantes y sumecanismo <strong>de</strong> acción.- Describir otros mecanismos físicos y químicos queparticipan en el metabolismo <strong>de</strong> la glucosa y quecontribuyen a mantener su concentración plasmática conmínima variabilidad.TRABAJO PRACTICODeterminación <strong>de</strong> la glucemia:El conocimiento <strong>de</strong>l metabolismo <strong>de</strong> loshidratos <strong>de</strong> carbono es muy importante entodo individuo, ya que el <strong>de</strong>tectaranormalida<strong>de</strong>s en el mismo nos orientaráhacia las alteraciones hormonalesimplicadas, fundamentalmente la diabetesque es una enfermedad prevalente en lapoblación.Un método que permite <strong>de</strong>tectar dichasanormalida<strong>de</strong>s es la <strong>de</strong>terminación <strong>de</strong> laglucemia, término que sigue siendo usado,aunque los métodos actuales <strong>de</strong> medición nousan sangre total sino suero o plasma queson equivalentes en sus valores, por lo queel término correcto sería glucosa sérica oplasmática, según como se obtenga lamuestra.La sangre total sólo se utiliza para la<strong>de</strong>terminación con tiras reactivas, técnicasemicuantitativa muy útil para elseguimiento.Los valores normales en ayunas <strong>de</strong> glucosaplasmática oscilan en un rango <strong>de</strong> 70 a 110mg/dL.El hallazgo <strong>de</strong> valores superiores a 126mg/dL permitirá diagnosticar que un pacientepa<strong>de</strong>ce diabetes, sobre todo si estos valoresse <strong>de</strong>tectan en más <strong>de</strong> una oportunidad.Estos valores aumentados pue<strong>de</strong>n ser<strong>de</strong>bidos a una acción disminuida y/o niveldisminuido <strong>de</strong> la hormona insulinaLa <strong>de</strong>terminación <strong>de</strong> la glucosa plasmáticapodrá realizarse:A - EN AYUNAS, por los métodos clásicos <strong>de</strong>laboratorio como:- Glucosa - oxidasa- Ortotoluidina- Somogy - Nelson, o Ferrocianuro <strong>de</strong> HoffmanValores Normales : 70 - 110 mg/dLEmbarazada : 60 - 95 m/dL.B - PRUEBA DE SOBRECARGA OTOLERANCIA ORAL (P.T.O.G.):Se sustenta en la ingestión <strong>de</strong> sustanciasexógenas como la glucosa, que estimula alpáncreas e induce a la secreción <strong>de</strong> insulina.Este estímulo exige al páncreas un esfuerzomáximo para mantener la glucemia <strong>de</strong>ntro <strong>de</strong>límites normales.Indicaciones De La Prueba De ToleranciaOrala) Antece<strong>de</strong>ntes <strong>de</strong> glucosurias o hiperglucemiasocasionales.b) Signos y síntomas diabéticos con glucemiasnormales.c) Antece<strong>de</strong>ntes hereditarios y obesidad con algúnelemento clínico <strong>de</strong> sospechad) Antece<strong>de</strong>ntes obstétricos anormales.e) Patología vascular precoz.f) Hipoglucemias reactivas preclínicas.g) Glucemia al azar o post -prandial anormal enembarazadas, enfermos o recibiendo drogashiperglucemiantes.METODOLOGÍA1 . Se administra 75 gr. <strong>de</strong> nutrosa en 375 ml<strong>de</strong> agua por vía oral (que pue<strong>de</strong>acidularse con jugo <strong>de</strong> limón para mejorarla tolerancia) a ingerir en 5 minutos2 . Obtener sangre a los 0, 60 y 120 min.Valores LimitesAyuno 60 min. 120 min.110 mg/dL < 190 mg/dL < 140 mg/dL3. Valores normales <strong>de</strong> glucemia en laP.T.G.O. para embarazadas (Criterio:National Diabetes Data Group) yConsenso Argentino <strong>de</strong> Diabetes.GLUCEMIAS PLASMA VENOSOEn ayunas< 105 mg/dL2 hora < 140 mg/dL
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 3C - DETECCIÓN RÁPIDA CON SANGRECAPILAR Y TIRAS REACTIVAS. (éstas podránser Detrostix. Glucostix y reflectómetro).METODOLOGÍA :1. Se limpiará el pulpejo <strong>de</strong> un <strong>de</strong>do conalcohol.2. Se efectuará la punción <strong>de</strong>l mismo conaguja estéril.3. Se colocará una gota <strong>de</strong> sangre sobre elsitio reactivo <strong>de</strong> la tira.4. Se <strong>de</strong>jará incubar un minuto y se secarácon algodón.5. Se realizará la lectura por comparacióncon los colores patrones a los 60segundos siguientes y se <strong>de</strong>terminaráasí el nivel <strong>de</strong> glucemia.6. El reflectómetro realiza la lectura <strong>de</strong> laglucemia en menos <strong>de</strong> 15‘’PREGUNTAS A CONTESTAR DURANTE LAAUTOINSTRUCCIÓN:1. GLUCEMIA: concepto, variacionesfisiológicas, valores normales según losmomentos <strong>de</strong>l día en relación a laingesta, métodos <strong>de</strong> <strong>de</strong>terminación.2. Distribución <strong>de</strong> la glucosa en elorganismo.3. Utilización <strong>de</strong> los recursos energéticosen el ayuno.4. Regulación <strong>de</strong> la glucemia :a) Insulinab) Hormonas hiperglucemiantes.Valores LimitesPostprandial o post PTOGAyuno 60 min. 120 min.4< 110 mg/dL < 190 mg/dL < 140 mg/dLTrabajo Practico:Se realizará esta evaluación cuantitativa en:1 . Alumno normal con ayuno mayor <strong>de</strong> 4 h.2 . Alumno normal en situación post - prandial (2h)luego <strong>de</strong> una comida <strong>de</strong>finida.3 . Alumno con antece<strong>de</strong>ntes familiares <strong>de</strong> diabetes y/osobrepeso, en situación post - prandial 2 h. luego <strong>de</strong>una comida <strong>de</strong>finida.D - CON TIRAS REACTIVAS PARAORINA se <strong>de</strong>terminará la presencia o no<strong>de</strong> glucosa. En condiciones normales,como toda la glucosa filtrada en elglomérulo renal es reabsorbida en lostúbulos contorneados proximales estamolécula <strong>de</strong>be estar ausente en orina(glucosuria negativa.Otras metodologías <strong>de</strong> valor clínico quesugieren alteraciones <strong>de</strong> la glucemiaE - HEMOGLOBINA GLICOSILADALa glucosa plasmática glicosila numerososmoléculas plasmáticas, entre ellas laHemoglobina. Cuando el valor <strong>de</strong> glucosaplasmática es normal, la glicosilación <strong>de</strong> laHgb es < <strong>de</strong>l 6 %.
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 4FISIOLOGIA GENITAL FEMENINA( Dra. Lilian BARRIOS)OBJETIVOS: Que el alumnoDescriba los componentes <strong>de</strong>l sistemareproductor femenino y sufuncionamiento.Explique la función <strong>de</strong> los distintosórganos que lo integran, <strong>de</strong>tallando lainterrelación que existe entre ellos.I<strong>de</strong>ntifique la repercusión somática ypsíquica <strong>de</strong> la función sexual en la mujer.Detalle los servomecanismos <strong>de</strong>regulación entre los distintos niveles <strong>de</strong>control <strong>de</strong> la función <strong>de</strong>l sistemareproductor femenino.PERIODO EMBRIONARIO - FETALSexo genético: (genotipo) elcontenido cromosómico <strong>de</strong> la hembra normales <strong>de</strong> 44 autosomas y 2 cromosomassexuales (XX)Sexo gonadal; (fenotipo): en lascinco primeras semanas <strong>de</strong> gestación, lasgónadas se <strong>de</strong>sarrollan sobre estructurassexualmente no diferenciables, por lamigración <strong>de</strong> 1000 a 2000 células que seoriginan en el endo<strong>de</strong>rmo <strong>de</strong>l saco vitelino(cerca <strong>de</strong>l extremo caudal <strong>de</strong>l embrión) yque emigran hacia la cresta genital pormecanismos que aún no son totalmenteconocidos. (Fig. 1)INTRODUCCIONEl patrón <strong>de</strong> evolución <strong>de</strong> lareproducción sexual humana involucra elmantenimiento y maduración <strong>de</strong> las célulasgerminales individuales <strong>de</strong> mujeres yhombres, su unión exitosa y el crecimiento y<strong>de</strong>sarrollo subsiguiente <strong>de</strong>l huevo.Des<strong>de</strong> este punto <strong>de</strong> vista, las basesfundamentales <strong>de</strong> la función ovárica giranalre<strong>de</strong>dor <strong>de</strong> la conservación <strong>de</strong> la especie.La función gonadal y la reproducciónen el ser humano están controladas en sumayor parte, por hormonas hipofisarias<strong>de</strong>nominadas gonadotrofinas, que se unencon sus receptores específicos en losovarios y testículos para regular laesteroidogénesis y la gametogénesis.Dichas hormonas a<strong>de</strong>nohipofisarias,a su vez, son reguladas por hormonashipotalámicas que constituyen una vía finalentre cerebro y a<strong>de</strong>nohipófisisconstituyéndose un eje endocrino hipotálamo- hipófiso - gonadalEl ovario, <strong>de</strong>ntro <strong>de</strong> este ejehipotálamo - hipófiso- ovárico, había sidoconsi<strong>de</strong>rado jugando un papel secundario,estimándose como glándula directora a lahipófisis. Sin embargo, a la luz <strong>de</strong> losconocimientos actuales, el ovario jugaría unpapel activo en la iniciación y mantenimiento<strong>de</strong> los ciclos reproductores a través <strong>de</strong> suscambiantes secreciones endocrinas y, seconsi<strong>de</strong>ra que el hipotálamo y laa<strong>de</strong>nohipófisis jugarían un papel permisivo.Fig. 1 Desarrollo <strong>de</strong>l ovarioCuando llegan a la crestagenital, las células germinales premeióticasreciben el nombre general <strong>de</strong> oogonios, ydurante las semanas 6a y 7a sufren unimportante proceso <strong>de</strong> mitosis llegando a unnúmero <strong>de</strong> alre<strong>de</strong>dor <strong>de</strong> 10.000constituyendoaún un estadio sexualmente nodiferenciado.A las 8 semanas hay un número <strong>de</strong>alre<strong>de</strong>dor <strong>de</strong> 600.000 oogonios.Si no hay diferenciación <strong>de</strong> gónadamasculina ( XY <strong>de</strong>pendiente) hacia la 7asemana <strong>de</strong> gestación , las células gonadalesindiferenciadas sufren un pasaje <strong>de</strong>oogonio a oocito primario <strong>de</strong>tenido enprofase <strong>de</strong> la primera división meióticahacia la 8a semana , dando i<strong>de</strong>ntificaciónfemenina a la gónada.A partir <strong>de</strong> la 8a semana seproducen simultáneamente fenómenos <strong>de</strong>mitosis, meiosis y atresia <strong>de</strong> los oogonios ,<strong>de</strong>ntro <strong>de</strong>l ovario en <strong>de</strong>sarrollo.Como resultado <strong>de</strong> todos estosprocesos, alre<strong>de</strong>dor <strong>de</strong>l 5º mes <strong>de</strong> embarazo,
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 5el ovario fetal tiene alre<strong>de</strong>dor <strong>de</strong> 6 a 7 x 10 6células germinales <strong>de</strong> las cuales un 66 %aproximadamente son oocitos primariosintrameióticos y un 34 % oogonios.En el período fetal , hacia la 16semana <strong>de</strong> gestación , aparecen losprimeros folículos primordiales , formadospor un oocito primario y células fusiformes(<strong>de</strong>nominadas <strong>de</strong> pregranulosa), con undiámetro <strong>de</strong> aproximadamente 30 µm <strong>de</strong> ∅.Se estima que este estadio esgonadotrofina in<strong>de</strong>pendienteAl quinto mes <strong>de</strong> gestación seproduce la conversión <strong>de</strong> los folículosprimordiales en folículos primarios, cuandolas células fusiformes <strong>de</strong> la pregranulosa <strong>de</strong>algunos folículos primordiales setransforman en una capa <strong>de</strong> células cuboi<strong>de</strong>s<strong>de</strong> la granulosa.Las células <strong>de</strong> la granulosa producenmucopolisacáridos que se dispone como unhalo translúcido alre<strong>de</strong>dor <strong>de</strong>l oocitoformando la zona pelúcida. A pesar <strong>de</strong> lapresencia <strong>de</strong> esta capa, las células <strong>de</strong> lagranulosa presentan prolongaciones que lasponen en contacto con el oocito permitiendosu nutrición.La conversión <strong>de</strong> folículo primordialen folículo primario implica un crecimiento yse observa el aumento <strong>de</strong> diámetro <strong>de</strong>loocito <strong>de</strong> unos 20 a unos 70 a 80 µm <strong>de</strong>diámetro.El proceso <strong>de</strong> mitosis <strong>de</strong> losoogonios disminuye a partir <strong>de</strong>l 5o mes y seagota al final <strong>de</strong>l 7o., mientras que elproceso <strong>de</strong> atresia folicular se acentúa <strong>de</strong>manera que en el momento <strong>de</strong>l nacimiento,solo hay alre<strong>de</strong>dor <strong>de</strong> 1 a 2 x 10 6 célulasgerminales ,significando que durante el período fetal seha perdido el 80 % <strong>de</strong> la dotación <strong>de</strong> célulasgerminales que se había alcanzado al 5ºmes.A<strong>de</strong>más, si no hubo diferenciación<strong>de</strong> conductos genitales y genitales externosmasculinos (proceso que implica ladiferenciación <strong>de</strong> células <strong>de</strong> Sértoli a la 7asemana <strong>de</strong> gestación para producir el Factor<strong>de</strong> Inhibición Mülleriana que produce laatrofia <strong>de</strong>l conducto <strong>de</strong> Müller con<strong>de</strong>sarrollo <strong>de</strong>l conducto <strong>de</strong> Wölf y laaparición <strong>de</strong> Células <strong>de</strong> Leydig a la 8asemana) a la 9a semana <strong>de</strong> gestacióncomienza el <strong>de</strong>sarrollo y diferenciación <strong>de</strong>ltracto genital femenino.Este proceso consiste en ladiferenciación <strong>de</strong> las trompas <strong>de</strong> Falopio,útero, vagina y genitales externos y seconsi<strong>de</strong>ra hormono - <strong>de</strong>pendiente.Se agregan a los órganos antesmencionados otras dos estructuras: la mamay el eje hipotálamo- a<strong>de</strong>nohipofisario que sibien no son genitales, tienen estrecharelación con la actividad sexual.A nivel fetal, se ha podido<strong>de</strong>terminar la presencia <strong>de</strong> célulasproductoras <strong>de</strong> LH en la hipófisis humanahacia las semanas 10 <strong>de</strong> gestación , se hahallado GnRH en hipotálamos <strong>de</strong> 14semanas y se ha <strong>de</strong>tectado liberaciónpulsátil <strong>de</strong> GnRH en fetos <strong>de</strong> 20 a 23semanas.La concentración <strong>de</strong> LH en suero <strong>de</strong>hipófisis <strong>de</strong> fetos femeninos es mucho masalta que en fetos masculinos en el segundotrimestre <strong>de</strong> la gestación lo cuál genera laposibilidad <strong>de</strong> que el testículo fetal produzcauna acción <strong>de</strong> retroalimentación negativasobre las neuronas productoras <strong>de</strong> GnRHNIÑEZSe caracteriza por la secreción <strong>de</strong>escaso nivel, no pulsátil <strong>de</strong> gonadotrofinas yestrógenos.La secreción <strong>de</strong> gonadotrofinas haceuna curva en forma <strong>de</strong> U durante la niñez,con un nivel elevado <strong>de</strong> estas hormonasdurante el primer año <strong>de</strong> vida, seguido poruna disminución a niveles <strong>de</strong>hipogonadotrofismo a los 6 a 8 años , coninhibición central <strong>de</strong> la secreción <strong>de</strong> GnRHhipotalámica. (Fig.2)Fig.2: Secreción <strong>de</strong> GnRH.
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 6Hay crecimiento somático importante conpoco <strong>de</strong>sarrollo <strong>de</strong>l aparato genital.. En esteperíodo continúa la pérdida <strong>de</strong> célulasgerminales por atresia folicular quedandosolo alre<strong>de</strong>dor <strong>de</strong> 300.000 folículos alcomienzo <strong>de</strong> la pubertad.El comienzo <strong>de</strong> la pubertad se iniciacon un aumento pulsátil <strong>de</strong> la liberación <strong>de</strong>GnRH que al principio se produce solodurante el sueño y recién <strong>de</strong>spués <strong>de</strong> ciertotiempo, también aparece durante la vigilia.(Fig. 3)En este período existen todos losestadios <strong>de</strong> <strong>de</strong>sarrollo folicular, salvo losovulatorios.Los folículos secundarios ,caracterizado por la presencia <strong>de</strong> múltiplescapas <strong>de</strong> células granulosas que han<strong>de</strong>sarrollado receptores para FSH,estrógenos y andrógenos constituyen elpool <strong>de</strong> folículos preantrales a partir <strong>de</strong> loscuales se producirá el reclutamiento ,probablemente <strong>de</strong>pendiente <strong>de</strong>gonadotrofinas en los ciclos ováricos,<strong>de</strong>spués <strong>de</strong> la pubertad.Los folículos secundarios que estánen proceso <strong>de</strong> diferenciación migran hacia lamédula y adquieren un componente tecal.En general el tamañofolicular está claramente relacionado con laedad, y los mayores diámetros <strong>de</strong> folículosatrésicos se encuentran a nivel <strong>de</strong> ovariospremenárquicos.PUBERTADEn la mujer se iniciaaproximadamente a los 10 años <strong>de</strong> edad conun aumento <strong>de</strong> secreción <strong>de</strong> FSH con bajosniveles <strong>de</strong> LH.El primer signo físico es elcrecimiento <strong>de</strong> las mamas que coinci<strong>de</strong> conla primera elevación <strong>de</strong>tectable <strong>de</strong> estradiolen plasma.Este período culmina con la menarcaentre los 11 y 15 años <strong>de</strong> edad, momento enque los niveles <strong>de</strong> LH aumentanagudamente, apareciendo el patrón cíclico<strong>de</strong> secreción <strong>de</strong> gonadotrofinas.En este período se <strong>de</strong>sarrollan loscaracteres sexuales secundarios y coinci<strong>de</strong>con la inserción afectiva e intelectual <strong>de</strong>ladolescente en la sociedad <strong>de</strong> los adultos.Es un período fundamentalmente estrógeno -<strong>de</strong>pendiente.Fig.3: Desarrollo <strong>de</strong> la secreciónpulsátil <strong>de</strong> LHLa aparición <strong>de</strong> esta secreciónhormonal pulsátil en la pubertad se asociacon un aumento <strong>de</strong>l tono opioidérgico y unaexpresión aumentada <strong>de</strong>l gen <strong>de</strong> proopiomelanocortinaa nivel hipotálamo -hipofisario .También se involucran es esteperíodo al aspartato y al glutamato. Elglutamato aumenta la liberación <strong>de</strong> GnRH invitro y el N- metil -D-aspartato (un análogo<strong>de</strong>l aspartato) administrado IV en formaintermitente por períodos prolongados dacomo resultado el comienzo <strong>de</strong> la puberta<strong>de</strong>n monas prepúberes.La aparición <strong>de</strong> la parte ascen<strong>de</strong>nte<strong>de</strong> la curva <strong>de</strong> secreción <strong>de</strong> GnRH que seinicia aproximadamente a los 10 años <strong>de</strong>edad podría <strong>de</strong>berse tanto a una disminución<strong>de</strong> reguladores inhibidores como a unaumento <strong>de</strong> estimuladores .También es importante para laactivación <strong>de</strong>l ciclo ovárico el aumentodurante el sueño <strong>de</strong> la pulsatilidad <strong>de</strong> GnRHy <strong>de</strong> LH.Antes <strong>de</strong> la aparición <strong>de</strong> ciclosregulares, pue<strong>de</strong>n presentarse fases
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 7luteínicas ineficientes y ciclos anovulatoriosen la fase puberal femeninaADULTEZConstituye el estadio don<strong>de</strong> la mujerestá capacitada para la reproducción.Depen<strong>de</strong> <strong>de</strong> la actividad cíclica yregular <strong>de</strong>l hipotálamo, la hipófisis, losovarios, el útero y el resto <strong>de</strong> las estructurasgenitales.También la a<strong>de</strong>cuada expresión <strong>de</strong>los genes <strong>de</strong> las distintas ca<strong>de</strong>nas <strong>de</strong> FSH yLH <strong>de</strong>pen<strong>de</strong> <strong>de</strong> una a<strong>de</strong>cuada frecuencia <strong>de</strong>pulsos <strong>de</strong> GnRH.NEURONAS PRODUCTORAS DE GnRHLas neuronas productoras <strong>de</strong> GnRH en elser humano están ubicadas en el núcleoparaventricular posterior <strong>de</strong>l hipotálamomedio basal y en el área preóptica <strong>de</strong>lhipotálamo . (Fig. 4)Dicha actividad cíclica se conocecomo CICLO GENITAL FEMENINO. Suduración promedio es <strong>de</strong> 28 días si bien surango <strong>de</strong> variación pue<strong>de</strong> ir <strong>de</strong> 21 a 35 o 45días.EJE HIPOTALAMO – HIPOFISO - GONADALHIPOTALAMO: neuronas hipotalámicas enforma <strong>de</strong> re<strong>de</strong>s laxas, distribuidasprincipalmente en el núcleo periventricular<strong>de</strong>l hipotálamo medial basal y el áreapreóptica <strong>de</strong> la parte anterior <strong>de</strong>lhipotálamo producen la liberina <strong>de</strong>gonadotrofinas hipofisarias (GnRH).Estas neuronas tienen conductarítmica con <strong>de</strong>scargas agudas <strong>de</strong> cortaduración, <strong>de</strong> actividad eléctrica <strong>de</strong>multiunida<strong>de</strong>sLa GnRH es un péptido <strong>de</strong> 10aminoácidos que estimula la secreción <strong>de</strong>las hormonas a<strong>de</strong>nohipofisarias FSH y LH.La GnRH se secreta en formapulsátil, y <strong>de</strong>termina una secrecióntambién pulsátil <strong>de</strong> las gonadotrofinashipofisarias lo cuál es un factor crítico parala activación <strong>de</strong> la función gonadal y lafunción reproductiva.Su efecto sobre los gonadotrofossolo es eficaz en pulsos 60 a 90 minutos loque produce un aumento <strong>de</strong>l número <strong>de</strong> suspropios receptores sobre sus células blanco.Una frecuencia menor produceanovulación y amenorrea y una frecuenciamayor produce regulación en menos <strong>de</strong>lnúmero <strong>de</strong> <strong>de</strong> sus propios receptores.Fig.4: Neuronas productoras <strong>de</strong> GnRHLos axones <strong>de</strong> estas neuronas seproyectan hacia muchos sitios <strong>de</strong>l cerebro.Una parte importante termina en laeminencia media y allí <strong>de</strong>jan salir suneurosecreción que , a través <strong>de</strong>l sistemaportal hipotálamo hipofisario , llegan a lascélulas gonadotrofas. Otras terminalesaxónicas secretoras <strong>de</strong> GnRH llegan alsistema límbico y otras áreas cerebrales .La GnRH se produce como una pre -pro -hormona <strong>de</strong> 92 aminoácidos que luegoes clivada.En forma experimental en monos seha establecido que el sistema neuronal <strong>de</strong>GnRH tiene una conducta rítmica con<strong>de</strong>scargas agudas y <strong>de</strong> corta duración <strong>de</strong>actividad eléctrica , pero no se ha podido<strong>de</strong>terminar si esto es una propiedadintrínseca <strong>de</strong> dichas neuronas o es reguladopor otras neuronas .Existe una gran sincronía entre lafrecuencia <strong>de</strong> pulsos <strong>de</strong> GnRH en la sangreportal hipotálamo a<strong>de</strong>nohipofisaria y lospulsos <strong>de</strong> LH en sangre circulanteLa periodicidad <strong>de</strong> pulsos <strong>de</strong> laGnRH humana ha sido estudiada enhipotálamo medial basal aislados habiéndose
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 8<strong>de</strong>terminado que pasa <strong>de</strong> una periodicidad<strong>de</strong> 60 minutos durante la vida fetal a 60 a100 minutos en el adultoEs importante señalar que lasneuronas hipotalámicas productoras <strong>de</strong>GnRH sufren la modulación <strong>de</strong> otras.neuronas <strong>de</strong> naturaleza adrenérgica,dopaminérgica y opiodoérgica . También laliberina <strong>de</strong> ACTH (CRH) inhibe laperiodicidad <strong>de</strong> los pulsos <strong>de</strong> GnRH. Elaspartato en cambio, estimula la liberación<strong>de</strong> GnRHLa GnRH por si misma, ejerce unsistema <strong>de</strong> retroalimentación <strong>de</strong> asa corta.Esta liberina <strong>de</strong> gonadotrofinasejerce su acción uniéndose a receptoresespecíficos, ubicados en la membrana <strong>de</strong> lascélulas gonadotrofas hipofisarias. Una vezfijada la GnRH a su receptor, estos sufrenun proceso <strong>de</strong> dimerización einternalización.Una vez <strong>de</strong>ntro <strong>de</strong> las células elcomplejo GnRH - Receptor se separa y losreceptores pue<strong>de</strong>n volver a la superficie enun proceso <strong>de</strong> reciclaciónEl receptor <strong>de</strong> la GnRH es una glucoproteína<strong>de</strong> 60 kDUna vez que la GnRH se ha unido asu receptor en el gonadotrofo, se produce laestimulación <strong>de</strong> la síntesis y liberación <strong>de</strong>las gonadotrofinas hipofisarias .El efecto <strong>de</strong> GnRH <strong>de</strong> estimular laliberación <strong>de</strong> gonadotrofinas hipofisarias selogra mediante la interacción <strong>de</strong>l complejoreceptor -GnRH con la proteína G <strong>de</strong>membrana, que, ante la presencia <strong>de</strong> estecomplejo , libera GDP y fija GTP.Una vez unida la proteína G a GTP,ella activa la Fosfolipasa C que forma IP3(trifosfato <strong>de</strong> inositol) y DAG (diacilglicerol)a partir <strong>de</strong> fosfatidilinositol <strong>de</strong> la membranacelular. (Fig. 5)Fig. 5 : Segundos mensajeros <strong>de</strong> GnRHEl IP3 libera Ca + intracelular <strong>de</strong><strong>de</strong>pósitos y abre canales <strong>de</strong> Ca + . Esteaumento <strong>de</strong> Ca+ intracelular permite elproceso <strong>de</strong> exocitosis <strong>de</strong> los gránulos <strong>de</strong>secreción <strong>de</strong>l godatotrofo, que contienenFSH y LH .Por otro lado, el DAG, activa a laproteinoquinasa C (PK-C) que activa latranscripción <strong>de</strong> los genes para la síntesis<strong>de</strong> proteínas.La regulación <strong>de</strong> la síntesis ysecreción <strong>de</strong> FSH y LH por medio <strong>de</strong> lospulsos <strong>de</strong> GnRH se integra con lainfluencia <strong>de</strong> los estrógenos, laprogesterona, la inhibina, la activina y lafolistatina , todas hormonas producidas porel ovario y que actúan sobre losgonadotrofos para producir la secrecióndiferencial <strong>de</strong> FSH y LH.La GnRH es <strong>de</strong>gradada porpeptidasas hipotalámicas e hipofisariasExisten agonistas <strong>de</strong> GnRH queproducen regulación en menos <strong>de</strong>l ejeformado por las gonadotrofinas y lasgónadas, precedido por un período <strong>de</strong>estimulación <strong>de</strong> la secreción durante las dosprimeras semanas <strong>de</strong> tratamiento.También existen antagonistas <strong>de</strong> laGnRH , que tienen la ventaja <strong>de</strong> disminuirinmediatamente el nivel <strong>de</strong> secreción <strong>de</strong> lasgonadotrofinas circulantes.El conocimiento <strong>de</strong> lascaracterísticas <strong>de</strong> la secreción <strong>de</strong> GnRH ysu disponibilidad y la <strong>de</strong> moléculasagonistas para fines terapéuticos ha tenidoaplicación clínica para pacientes con<strong>de</strong>ficiencia <strong>de</strong> formación <strong>de</strong> GnRH endógena: pubertad precoz, endometriosis y síndrome
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 9<strong>de</strong> ovario poliquístico, esterilidad, cáncerginecológico y mamarioADENOHIPOFISIS:La función gonadal y la reproducciónen la mujer están controladasprincipalmente por las hormonasgonadotrofinas hipofisarias FSH, LH. (Fig.6)El peso molecular <strong>de</strong> FSH es <strong>de</strong> 33kD y el <strong>de</strong>LH <strong>de</strong> 28 kD.Las células a<strong>de</strong>nohipofisariasresponsables <strong>de</strong> sintetizar gonadotrofinasFSH y LH (gonadotropos) aparecen enperíodos mas tempranos <strong>de</strong> la vida fetal quelas productoras <strong>de</strong> prolactina.En la hipófisis <strong>de</strong> la mujer adulta, losgonadotropos están distribuidos en toda lapars distales.Un 60 % <strong>de</strong> estas células en la ratas,contiene tanto FSH como LH; un 18 %contiene solo LH y un 23 % solo FSH.También los gonadotropos humanospresentan FSH y LH en la misma poblacióncelular y quizás ello explique el efecto <strong>de</strong>GnRH tanto sobre FSH como sobre LH .Fig.6: Secreción <strong>de</strong> FSH y LH duranteel ciclo menstrualLa regulación <strong>de</strong> la síntesis <strong>de</strong>gonadotrofinas FSH y LH se realiza mediantela integración <strong>de</strong> señales estimuladoras<strong>de</strong>pendientes <strong>de</strong> GnRH e inhibidoras<strong>de</strong>pendientes <strong>de</strong> estrógenos, progesterona,activina, inhibina y folistatina. (Fig.7)La FSH actúa sobre las célulasgranulosas en los ovarios promoviendo el<strong>de</strong>sarrollo <strong>de</strong>l folículo.Por su parte, la LH, participa en laovulación produciendo la ruptura <strong>de</strong>l folículopreovulatorio y en la producción <strong>de</strong>hormonas sexuales por parte <strong>de</strong> las célulasesteroidogénicas ovaricas.Las gonadotrofinas FSH y LH sonhormonas a<strong>de</strong>nohipofisarias glucoprotéicasconstituídas por dos subunida<strong>de</strong>spolipeptídicas glicosiladas <strong>de</strong>nominadasca<strong>de</strong>nas α y β que están unidas en formano covalente.La subunidad α es igual para las doshormonas (también para la gonadotrofinacoriónica) , mientras que la subunidad β esdistintiva para cada hormona y su unión a laca<strong>de</strong>na α es necesaria para que seproduzca la actividad hormonal específica.El contenido <strong>de</strong> hidratos <strong>de</strong> carbono<strong>de</strong> las gonadotrofinas es importante tantopara su actividad biológica y su <strong>de</strong>puraciónmetabólica.Fig.7: Regulación <strong>de</strong>l eje hipotálamo –hipófiso - ovárico
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 10El resultado final <strong>de</strong> estas señalesreguladoras se expresa en la tasa <strong>de</strong>expresión <strong>de</strong> los genes <strong>de</strong> lasgonadotrofinas.En general las secreciones ováricasejercen un efecto inhibidor sobrea<strong>de</strong>nohipófisis, lo cual se pone <strong>de</strong> manifiestopor el aumento <strong>de</strong> secreción <strong>de</strong>gonadotrofinas en los sujetos castrados ocon insuficiencia gonadal.La frecuencia <strong>de</strong> <strong>de</strong>scarga pulsátil <strong>de</strong>FSH y LH es importante para el <strong>de</strong>sarrollo <strong>de</strong>un ciclo menstrual normal.La frecuencia <strong>de</strong> <strong>de</strong>scarga <strong>de</strong> LHaumenta <strong>de</strong> 1 a 2 pulsos por hora <strong>de</strong>s<strong>de</strong> lafase folicular temprana hasta la fasefolicular tardía , con aumento <strong>de</strong> LH <strong>de</strong> basey disminuye 1 pulso cada 4 horas durante lafase lútea con una correspondientedisminución <strong>de</strong> LH basal .Estos cambios probablemente se<strong>de</strong>ban a los pulsos <strong>de</strong> secreción <strong>de</strong> GnRH ya la acción <strong>de</strong> los mecanismosretroalimentadores ováricos, sobre todo , losesteroidogénicos; ya que , según se hacomprobado en forma experimental enovejas, los estrógenos aumentan los pulsos<strong>de</strong> LH plasmática , mientras que laprogesterona los disminuye (probablemente<strong>de</strong>bido a una acción sobre los pulsos <strong>de</strong>GnRH.Adicionalmente, los estrógenostienen efecto sobre la sensibilidad <strong>de</strong> losreceptores gonadotropos a GnRH, en unarespuesta bifásica que primero disminuyedicha sensibilidad y luego la aumenta,estando ambas respuestas aumentadas enpresencia <strong>de</strong> progesterona.En el momento <strong>de</strong> la oleada <strong>de</strong> LH yFSH necesaria para la producción <strong>de</strong> laovulación a partir <strong>de</strong>l folículo predominanteprobablemente se junten: el efecto <strong>de</strong>homoregulación en más producido por GnRHsobre sus propios receptores, el efecto <strong>de</strong> lasensibilidad aumentada <strong>de</strong> los receptores <strong>de</strong>GnRH producido por los estrógenos y un altocontenido <strong>de</strong> hormonas en los gonadotropos.La fuerte liberación <strong>de</strong>gonadotrofinas que se produce en esemomento <strong>de</strong>l ciclo, produce una <strong>de</strong>plecióntransitoria <strong>de</strong> gonadotrofinas hipofisarias.El estradiol tiene efectos positivos ynegativos sobre la síntesis y liberación <strong>de</strong>gonadotrofinas: aumenta la síntesis <strong>de</strong>subunida<strong>de</strong>s beta <strong>de</strong> la LH y la secreción <strong>de</strong>LH inducida por GnRH , pero inhibe laliberación <strong>de</strong> GnRH a nivel hipotalámico.En el momento <strong>de</strong> la oleadahormonal, por otro lado, el estradiolpromovería la síntesis y liberación <strong>de</strong> GnRHhipotalámico.Otra molécula reguladora fisiológica,por retroalimentación <strong>de</strong> la secreción <strong>de</strong> lasgonadotrofinas sería el péptido ováricoInhibina , producido por las célulasgranulosas y lúteas ováricas. Este péptido( una glucoproteína <strong>de</strong> 32 kD formado pordos subunida<strong>de</strong>s <strong>de</strong>nominadas α y β ) esestimulado en su formación por FSH y él, asu vez, inhibe la síntesis y secreción <strong>de</strong>FSH.Las inhibina plasmática, aumentamarcadamente durante la estimulacióngonadotrófica <strong>de</strong> los ovarios <strong>de</strong> pacientes enpreparación para fertilización in vitro ydurante el embarazo (su fuente principal <strong>de</strong>producción, en este caso, es la placenta).Otro péptido ovárico, la Activina,está formado por dos subunida<strong>de</strong>s β <strong>de</strong>inhibina y actúa estimulando la secreción <strong>de</strong>FSH, aumentando la síntesis <strong>de</strong> sus ca<strong>de</strong>nasβ y liberando FSH completa.Otro péptido ovárico: la Folistatina,aislado <strong>de</strong> líquido folicular, inhibe lasecreción <strong>de</strong> FSH por medio <strong>de</strong> la inhibición<strong>de</strong> la síntesis <strong>de</strong> su ca<strong>de</strong>na β.La naturaleza pulsátil <strong>de</strong> laliberación <strong>de</strong> las gonadotrofinas se <strong>de</strong>tecta<strong>de</strong> forma rápida en muestras <strong>de</strong> sangreseriadas. Los pulsos se caracterizan por unaumento rápido <strong>de</strong> secreción <strong>de</strong> 10 a 15minutos, seguida por una caída que duraalre<strong>de</strong>dor <strong>de</strong> 50 minutos para LH y algo máspara FSH (esta última tiene una <strong>de</strong>puraciónplasmática más lenta).Los pulsos <strong>de</strong> FSH son maspequeños que los <strong>de</strong> LH.Las gonadotrofinas circulantescambian durante el ciclo sexual . La LHplasmática aumenta levemente en la fasefolicular tardía, sufre un fuerte aumento en
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 11el período preovulatorio y luego <strong>de</strong>clina enla fase lútea . La FSH aumenta en la fasefolicular temprana, presenta un pico en elmomento preovulatorio, menor al <strong>de</strong> LH yaumenta <strong>de</strong> nuevo en la fase lútea tardía(Fig. 8)En la menopausia las gonadotrofinasaumentan <strong>de</strong> forma marcada, con un nivel <strong>de</strong>FSH que supera al <strong>de</strong> LH, manteniéndose lasecreción pulsátil. (Fig. 9)por medio <strong>de</strong>l aumento <strong>de</strong> cAMP y poraumento <strong>de</strong>l número <strong>de</strong> receptores a FSH.FSH, por su lado, aumenta el número<strong>de</strong> receptores a LH en ratas inmaduras.A<strong>de</strong>más, FSH estimula laaromatización <strong>de</strong> los andrógenos a 17βestradiol. El hecho <strong>de</strong> que FSH aumenta laproducción <strong>de</strong> estrógenos mediante laestimulación <strong>de</strong> aromatasa que utiliza losandrógenos formados por la teca estimuladapor LH, ha llevado al concepto <strong>de</strong> doscélulas y dos gonadotrofinas para el control<strong>de</strong> la síntesis <strong>de</strong> los estrógenos. (Fig. Nº10)Fig. 9: secreción <strong>de</strong> gonadotrofinas enlas distintas etapas <strong>de</strong> la vidaLa vida media <strong>de</strong> FSH y LH es <strong>de</strong>aproximadamente 1 horaLH actúa sobre receptores <strong>de</strong>membrana <strong>de</strong> alta especificidad que tambiénligan Gonadotrofina coriónica, y que activana<strong>de</strong>nilato ciclasa al igual que FSH.Recor<strong>de</strong>mos que en circulación hayuna microheterogeneidad <strong>de</strong> FSH y LH<strong>de</strong>bido, en parte, a el grado <strong>de</strong> glucosilacióny la disponibilidad <strong>de</strong> ca<strong>de</strong>nas α y β.Función <strong>de</strong> FSH y LHLa FSH ejerce sus efectos biológicossobre las células granulosas <strong>de</strong> los ovarios.Estimula el crecimiento y maduración <strong>de</strong> losfolículos, actuando en forma conjunta con LHy estrógenos, hasta mas allá <strong>de</strong>l estadio <strong>de</strong>antro.Los estrógenos aumentan larespuesta <strong>de</strong> las células granulosas al FSHPor otro lado, los andrógenosformados en ovario , en realidad,dihidrotestosterona, pues actúa una αreductasa ovárica, podrían participar con unefecto inhibitorio <strong>de</strong>l <strong>de</strong>sarrollo folicular yprogresión hacia la atresia.Es importante recordar que lascélulas <strong>de</strong> la granulosa ovárica presentanreceptores para andrógenos, indicando queestas células son blanco <strong>de</strong> esta hormona.Los receptores para FSH solo seencuentran en las células granulosasováricas
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 12En cambio los receptores <strong>de</strong> LH sehallan en: células granulosas, célulasintersticiales, células lúteas y en la teca.Cuando aparecen los receptores paraLH (estimulados por FSH) en las células <strong>de</strong>la granulosa <strong>de</strong> los folículos preovulatoriosque ya han formado el antro, los receptores<strong>de</strong> FSH <strong>de</strong>saparecen, junto con la a<strong>de</strong>nilatociclasa FSH- <strong>de</strong>pendiente.La maduración folicular se asociacon la aparición <strong>de</strong> los receptores a LH y lasenzimas esteroidogénicas que produciránestrógenos y progesterona .La acción biológica <strong>de</strong> LH <strong>de</strong> estimular laesteroidogénesis está mediada por cAMP.Los mecanismos que <strong>de</strong>terminan quefolículo/s va/n a <strong>de</strong>sarrollarse y ovular ycuales van a sufrir atresia no son aúntotalmente conocidos y probablemente<strong>de</strong>pen<strong>de</strong>n <strong>de</strong> las interacción entre masa <strong>de</strong>folículos inmaduros y en <strong>de</strong>sarrollo con lasconcentraciones plasmáticas <strong>de</strong> FSH y LH yla concentración intraovárica <strong>de</strong> estrógeno.maduración folicular y la biosíntesisesteroidogénica antes y <strong>de</strong>spués <strong>de</strong> laovulación).La producción <strong>de</strong> la ovulación<strong>de</strong>pen<strong>de</strong> en forma crítica <strong>de</strong> la oleadapreovulatoria <strong>de</strong> LH. Si bien algunoshallazgos experimentales indican que la FSHinduciría la ovulación, LH es su estímuloprimario.En el folículo que va a ovular,previamente se produce una<strong>de</strong>polimerización <strong>de</strong> mucoproteínas <strong>de</strong>llíquido folicular con aumento <strong>de</strong> la presiónosmótica y aumento <strong>de</strong>l tamaño folicular.También se observa en este folículo una<strong>de</strong>gradación <strong>de</strong>l tejido conectivo <strong>de</strong> la paredfolicular, y formación <strong>de</strong> colagenasa. (Fig.Nº 11)La concentración <strong>de</strong> FSH y LH yestrógenos en el líquido folicular secorrelaciona con las concentracionesplasmáticas <strong>de</strong> dichas hormonas.Los folículos pequeños ( 8 mm ∅)contienen altos niveles <strong>de</strong> estradiol, y FSH(a pesar <strong>de</strong> que esta última hormona estádisminuyendo en plasma, probablemente suconcentración folicular aumenta <strong>de</strong>bido alaumento <strong>de</strong>l número <strong>de</strong> receptores <strong>de</strong> FSHpor acción <strong>de</strong>l estradiol).Durante la fase folicular tardía losfolículos gran<strong>de</strong>s presentan altasconcentraciones <strong>de</strong> FSH, LH, estradiol yprogesteronaEstos hallazgos sugieren que FSH esimportante en el <strong>de</strong>sarrollo <strong>de</strong> los folículospequeños y que FSH, LH y estradiol sonimportantes en el <strong>de</strong>sarrollo <strong>de</strong> los folículosgran<strong>de</strong>s y, adicionalmente, que el <strong>de</strong>sarrollofolicular <strong>de</strong>pen<strong>de</strong> <strong>de</strong> secuencias <strong>de</strong>presencias hormonales <strong>de</strong>finidas.De manera que la secuencia FSH;FSH + LH; FSH + LH + estradiol +progesterona) sería crítica para laFig. 11: OvulaciónLa colagenasa es primordial para laruptura <strong>de</strong> la pared folicular en el momento<strong>de</strong> la ovulación.También se ha <strong>de</strong>tectado que lascélulas <strong>de</strong> la granulosa producen gran<strong>de</strong>scantida<strong>de</strong>s <strong>de</strong> activador <strong>de</strong> plasminógenoque actuando sobre el plasminógeno <strong>de</strong>llíquido folicular produciría plasmina que consu actividad proteolítica, también jugaría unpapel en la ruptura folicular.La oleada preovulatoria <strong>de</strong> LHtambién produce la reanudación <strong>de</strong> ladivisión meiótica que se había <strong>de</strong>tenidodurante el período fetal, (probablemente<strong>de</strong>bido a un inhibidor producido por lascélulas <strong>de</strong> la granulosa, ya que los oocitosseparados <strong>de</strong> las células granulosas pue<strong>de</strong>madurar espontáneamente in vitro) por unmecanismo que parece involucrar a cAMP yprostaglandinas formadas en las células <strong>de</strong>la granulosa.Al reanudarse la división meiótica, eloocito primario, pier<strong>de</strong> un cuerpo polar y setransforma en oocito secundario.
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 13En el momento <strong>de</strong> la ovulación, eloocito secundario ro<strong>de</strong>ado <strong>de</strong> sus célulasgranulosas circundantes (cúmulo prolígero)es eliminados <strong>de</strong>l folículo e ingresan a latrompa <strong>de</strong> Falopio.OVARIOSPresenta una zona dominante: lacorteza, ro<strong>de</strong>ada por el epitelio germinal(Fig. 12)Hay evi<strong>de</strong>ncias <strong>de</strong> que las células<strong>de</strong> la granulosa no son uniformes y quehabría por lo menos tres poblacionesdiferentes.Las células <strong>de</strong> la granulosa cercanasa la membrana basal serían las mas activaspara la esteroidogénesis, presentando altocontenido <strong>de</strong> 3β hidroxiesteroi<strong>de</strong><strong>de</strong>shidrogenasa,glucosa-6-fosfato<strong>de</strong>shidrogenasa y citocromo P450 y tendríanmayor cantidad <strong>de</strong> receptores para LH.Esto difiere con las células cercanasal oocito y las que ro<strong>de</strong>an al antro. Sepostula que las células <strong>de</strong>l cúmulo podríanser células madres encargadas <strong>de</strong> multiplicarlas células granulosasPor otro lado, las célulasintersticiales y <strong>de</strong> la teca <strong>de</strong>l ovario, tienencapacidad <strong>de</strong> sintetizar andrógenos.Las células intersticiales se ubicanen la corteza y en la médula ovárica .Fig. 12: Desarrollo folicularContiene los oocitos, cada unoencapsulado <strong>de</strong>ntro <strong>de</strong> un folículo ro<strong>de</strong>ado<strong>de</strong> estroma , en todos los períodos <strong>de</strong><strong>de</strong>sarrollo. Se observa un alto número <strong>de</strong>folículos atrésicos.Otra zona: la médula, contieneestroma y el hilio <strong>de</strong>l órgano.El oocito primario y las células <strong>de</strong> lagranulosa <strong>de</strong>scansan sobre una membranabasal que los separa <strong>de</strong>l estromacircundante.De esta manera, las células <strong>de</strong> lagranulosa no poseen flujo sanguíneo directohasta el momento <strong>de</strong> la rotura folicular.Estas células se caracterizan porconectarse entre sí a través <strong>de</strong> uniones anivel <strong>de</strong> membrana, lo cual las transformaen un sincicio funcional.Adicionalmente, estas células emitenfinas prolongaciones citoplasmáticas quelas unen al oocito y se estima que a través<strong>de</strong> estas uniones las células <strong>de</strong>l cúmuloprolígero ejercen un control inhibitoriosobre la reanudación <strong>de</strong> la meiosis <strong>de</strong>loocito primario.Se han i<strong>de</strong>ntificado varias clases <strong>de</strong>células intersticiales, entre las que po<strong>de</strong>mosmencionar:células intersticiales primarias, que enforma transitoria producen andrógenosen la médula ovárica fetal, hasta las 20semanas <strong>de</strong> gestacióncélulas intersticiales y <strong>de</strong> la teca: sonlos principales productores <strong>de</strong>andrógenoscélulas intersticiales secundarias: soncélulas <strong>de</strong> la teca interna <strong>de</strong> folículosatrésicos, no producen andrógenos(tienen inervación noradrenérgica)células intersticiales <strong>de</strong>l hilio: tambiénson productoras <strong>de</strong> andrógenos.Un fenómenos llamativo en el ovarioes la infiltración <strong>de</strong> leucocitos que aumentaa medida que progresa la fase folicular.Esta infiltración compren<strong>de</strong> lallegada temprana <strong>de</strong> macrófagos ymastocitos .En el momento <strong>de</strong> pico agudo <strong>de</strong>secreción <strong>de</strong> LH preovulatoria los mastocitossufren un proceso <strong>de</strong> <strong>de</strong>granulación que
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 14produce hiperemia por acción <strong>de</strong> histaminay PGE2Estas moléculas causanvasodilatación y aumento <strong>de</strong> permeabilidadcapilar.Al formarse el cuerpo lúteo, sepresenta una infiltración <strong>de</strong>l mismo coneosinófilos y linfocitos y se ha postulado quesustancias citotóxicas formadas por loseosinófilos podrían generar lesiones <strong>de</strong> lacélula lútea que terminaría a su muerte. Porotro lado, los macrófagos estaríanencargados <strong>de</strong> fagocitar las células muertas.Adicionalmente, el bFGF, TGFα yTGFβ producidos por los macrófagos queinfiltran el ovario, podían ejercer efectosreguladores <strong>de</strong>l crecimiento y función <strong>de</strong> lascélulas <strong>de</strong> la granulosa y lúteas ováricas.DESARROLLO FOLICULAREn los ovarios <strong>de</strong> la mujer adulta seobservan distintos tipos <strong>de</strong> folículos:folículos primordiales ( <strong>de</strong> 30 a 60 µm<strong>de</strong> ∅) que contienen un oocitoprimario ro<strong>de</strong>ado <strong>de</strong> una capa <strong>de</strong>células <strong>de</strong> pregranulosa, aplanadasfolículos primarios ( <strong>de</strong> más <strong>de</strong> 60 µm<strong>de</strong> ∅) que contienen un oocitoprimario ro<strong>de</strong>ado <strong>de</strong> una sola capa <strong>de</strong>células granulosas cuboi<strong>de</strong>sfolículos secundarios ( <strong>de</strong> 120 µm <strong>de</strong>∅) que contienen un oocito primarioro<strong>de</strong>ado <strong>de</strong> capas <strong>de</strong> célulasgranulosas cuboi<strong>de</strong>sEsta clasificación, ya establecida porGougeon, esta <strong>de</strong>finida en gran medida porlas variaciones en el número <strong>de</strong> células <strong>de</strong> lagranulosa que presenta el folículo en elproceso <strong>de</strong> maduración, a los cuales se losclasifica <strong>de</strong> estadio 1 a 8. (Fig. 13)El proceso por el cual se produce laconversión <strong>de</strong> folículos clase 1 en folículo<strong>de</strong> clase 4 (antrales ) con diámetro <strong>de</strong> 2 mmse <strong>de</strong>nomina período tónico <strong>de</strong>l <strong>de</strong>sarrollofolicular.El crecimiento se caracteriza por unaumento <strong>de</strong> 600 veces en la cantidad <strong>de</strong>células <strong>de</strong> la granulosa junto con unaumento <strong>de</strong> 15 veces en el tamaño folicular .Este crecimiento se produce enforma lenta en presencia <strong>de</strong> cantida<strong>de</strong>srelativamente bajas <strong>de</strong> gonadotrofinas .Con la adquisición <strong>de</strong> teca interna,los folículos <strong>de</strong> clase 1 se vuelvensensibles a la estimulación gonadotróficaPara la <strong>de</strong>scripción <strong>de</strong> un ciclo <strong>de</strong>crecimiento <strong>de</strong> folículo <strong>de</strong> clase 1 a 8, seespecula que un folículo secundario setransforma en folículo <strong>de</strong> la clase 1 durantela parte temprana <strong>de</strong> la fase lútea (días 15 a19 <strong>de</strong>l ciclo) varios días <strong>de</strong>spués <strong>de</strong> laovulación <strong>de</strong> un ciclo sexual cualquiera<strong>de</strong>spués <strong>de</strong> la menarca.A este ciclo se lo <strong>de</strong>nomina CICLO1 para la <strong>de</strong>scripción <strong>de</strong> este proceso.Alre<strong>de</strong>dor <strong>de</strong> 25 días <strong>de</strong>spués ( es<strong>de</strong>cir entre los días 11 y 15 <strong>de</strong>l cicloovulatorio siguiente (CICLO 2 ) seproduce la conversión <strong>de</strong> los folículos <strong>de</strong>clase 1 en folículos <strong>de</strong> clase 2 (antraltemprano)20 días mas tar<strong>de</strong>, en la fase lútea<strong>de</strong>l CICLO 2 , los folículos <strong>de</strong> clase 2 seconvierten en folículos <strong>de</strong> clase 3 .15 días <strong>de</strong>spués, (durante la fasefolicular tardía <strong>de</strong>l siguiente CICLO 3 losfolículos <strong>de</strong> clase 3 se convierten enfolículos <strong>de</strong> clase 4 .
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 15Se estima que la conversión <strong>de</strong> clase4 a 5 se produce en la fase lútea avanzada<strong>de</strong>l CICLO 3 (días 25 a 28 <strong>de</strong> dicho ciclo).Cuando el folículo mi<strong>de</strong> alre<strong>de</strong>dor <strong>de</strong>200 µm se hace evi<strong>de</strong>nte la aparición <strong>de</strong>lagunas entre las células <strong>de</strong> la granulosa.La aparición <strong>de</strong> una cavidad con contenidolíquido , <strong>de</strong>nominada antro, transforma al folículo en unpequeño folículo antral, con el oocito ocupando unaposición excéntrica ro<strong>de</strong>ado por dos o tres capas <strong>de</strong>células granulosas que reciben el nombre <strong>de</strong> cúmuloprolígero. (Fig. 12)Los gran<strong>de</strong>s folículos antrales se<strong>de</strong>nominan folículos <strong>de</strong> Graaf.El líquido antral contiene estrógenos,progesterona, proteínas plasmáticas yováricas, proteoglucanos (producidos por lascélulas <strong>de</strong> la granulosa) y electrolitos.Si bien es probable que el <strong>de</strong>sarrollo<strong>de</strong> todos los estadios foliculares <strong>de</strong>pendan<strong>de</strong> las gonadotrofinas (en mayor o menormedida), los folículos mas gran<strong>de</strong>s seríangonadotrofinas -<strong>de</strong>pendientes para sumaduración.CICLO MENSTRUALLa liberación cíclica <strong>de</strong> lasgonadotrofinas <strong>de</strong>termina un <strong>de</strong>sarrollocíclico <strong>de</strong> folículos que van a ovular concambios regulares a nivel <strong>de</strong>l endometriouterino,Estos cambios reciben el nombre <strong>de</strong>ciclo menstrual,Ellos duran alre<strong>de</strong>dor <strong>de</strong> 28 díasdurante los años reproductivos, pero seproduce un aumento <strong>de</strong> los intervalosintermenstruales tanto en la adolescenciacomo en la transición menopáusica.En ambas épocas, hay secreciónaberrante <strong>de</strong> estradiol y gonadotrofinas yesto da como resultado la asincronía <strong>de</strong> loseventos <strong>de</strong>l ciclo que pue<strong>de</strong>n manifestarsecomo ciclos anovulatoriosCICLO OVARICOEl ciclo ovárico pue<strong>de</strong> dividirse en lassiguientes fases:Fase folicular (subdividida en temprana ytardía). Dura aproximadamente 13 días ydará como resultado la génesis <strong>de</strong> unfolículo preovulatorio mientras los otrosexperimentaran el proceso <strong>de</strong> atresiaFase ovulatoriaFase lútea (subdividida en temprana ytardíaFase menstrualFase folicularEl aumento <strong>de</strong> la secreción <strong>de</strong> FSH,junto con el retorno <strong>de</strong> la frecuencia rápida<strong>de</strong> la LH, inicia el reclutamiento <strong>de</strong> losfolículos durante los primeros 4 a 5 días <strong>de</strong>la fase folicular.Los folículos <strong>de</strong> clase 5, serían lacohorte <strong>de</strong> la cual van a ser reclutados losfolículos a ovular y este tardaríaaproximadamente 5 días en cada uno <strong>de</strong> losestadios que le faltan para llegar a clase 8.(Según esta <strong>de</strong>scripción <strong>de</strong><strong>de</strong>sarrollo folicular un folículo clase 1 tarda85 días en convertirse en clase 8)Durante este lapso se produce laselección <strong>de</strong> un folículo dominante queovularáEn los folículos <strong>de</strong> clase 5 en lafase folicular temprana, la FSH induce laactividad <strong>de</strong> aromatasa en las células <strong>de</strong> lagranulosa y esto hace que aumente laconcentración <strong>de</strong> estrógenos folicular, que asu vez estimula la captación <strong>de</strong> FSH <strong>de</strong>ntro<strong>de</strong>l líquido folicular y aumenta lasensibilidad <strong>de</strong>l folículo a esta hormona.Recor<strong>de</strong>mos que la biosíntesis <strong>de</strong>estrógenos en la fase folicular ováricarequiere <strong>de</strong> las células <strong>de</strong> la granulosa y<strong>de</strong> la teca y células intersticiales y sesintetizan los estrógenos según la teoría <strong>de</strong>dos células/dos gonadotrofinas, ya que lascélulas <strong>de</strong> la teca y las células intersticialesováricas producen andrógenos que sontransferidos a través <strong>de</strong> la membrana basal<strong>de</strong>l folículo a las células <strong>de</strong> la granulosadon<strong>de</strong> son aromatizados a estrógenos.En la parte media <strong>de</strong> la fasefolicular, un folículo, probablemente por
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 16haber producido por casualidad masestrógenos que el resto, aumenta laformación <strong>de</strong>l antro y la adquisición <strong>de</strong>receptores para LH. Esto lo convierte enfolículo dominante. Este folículo presentaalta capacidad para la biosíntesis y lasecreción <strong>de</strong> andrógenos, estrógenos,progestágenos e inhibina.En este folículo se produce unaumento importante <strong>de</strong> producción <strong>de</strong>estrógenos en la segunda parte <strong>de</strong> la fasefolicular, junto con una caída <strong>de</strong> FSH,elementos que serían centrales en lacapacidad <strong>de</strong> auto amplificación <strong>de</strong>l folículodominante. El resto <strong>de</strong> folículos que nolograron producir la suficiente cantidad <strong>de</strong>estrógenos, no se <strong>de</strong>sarrollan. (Fig. 14)En el folículo dominante, también seobservan cambios tecales. En el día 7 <strong>de</strong>lciclo, el folículo que va a ovular se ro<strong>de</strong>a <strong>de</strong>una teca que capta mas LH que la teca <strong>de</strong>lresto <strong>de</strong> folículos <strong>de</strong> la cohorte, <strong>de</strong> talmanera que al día 9, la irrigación <strong>de</strong> la teca<strong>de</strong>l folículo dominante es <strong>de</strong>l doble que la<strong>de</strong>l resto <strong>de</strong> folículos.Así, este folículo recibe mas LH, LDLy FSH que el resto <strong>de</strong> folículos en<strong>de</strong>sarrollo.Se ha postulado que el folículodominante sería capaz <strong>de</strong> secretarsustancias capaces <strong>de</strong> inhibir el resto <strong>de</strong>folículos <strong>de</strong>l mismo ovario y <strong>de</strong>l ovario <strong>de</strong>llado opuesto, pero otros investigadorescreen que esto no suce<strong>de</strong>ría .Fase <strong>de</strong> ovulaciónEsta fase es <strong>de</strong>pendiente <strong>de</strong> unaumento brusco <strong>de</strong> secreción <strong>de</strong> LH (oleada<strong>de</strong> LH y un aumento menor <strong>de</strong> FSH.Las oleadas <strong>de</strong> LH y FSH comienzanen forma brusca. La duración <strong>de</strong> la oleada<strong>de</strong> LH es <strong>de</strong> aproximadamente 48 hs y seacompaña <strong>de</strong> una rápida caída <strong>de</strong> losniveles circulantes <strong>de</strong> estradiol y <strong>de</strong> 17 OHprogesterona pero con un sostenidoaumento <strong>de</strong> inhibina.El tiempo preciso entre el comienzo<strong>de</strong> la oleada <strong>de</strong> LH y la ovulación aún se<strong>de</strong>sconoce con precisión, pero se estimaque la ovulación se produciría 35 a 44 hs<strong>de</strong>spués <strong>de</strong>l comienzo <strong>de</strong> la oleada <strong>de</strong> LH.Previo a la ovulación (cercano al día14) bajo la influencia <strong>de</strong> FSH y el pico <strong>de</strong> LH( que pue<strong>de</strong> prece<strong>de</strong>r hasta en 36 hs a laovulación), el folículo ovárico dominante(que aumentó su contenido en PGE2 , PGF2 α, activador <strong>de</strong> plasminógeno y colagenasa,activada por la plasmina formada por elactivador <strong>de</strong> plasminógeno ) presenta uncrecimiento súbito, produciéndose unatumefacción quística o abultamiento <strong>de</strong> lasuperficie <strong>de</strong>l ovario.Un pequeño punto avascular llamadoestigma aparece <strong>de</strong> pronto <strong>de</strong>ntro <strong>de</strong> eseabultamiento. Antes <strong>de</strong> que ocurra laovulación, el oocito y algunas células <strong>de</strong>lcúmulo prolígero se <strong>de</strong>spren<strong>de</strong>n <strong>de</strong>l interior<strong>de</strong>l folículo.El estigma se rompe, liberando aloocito junto con el líquido antral. La suaveexpulsión <strong>de</strong>l oocito es el resultado <strong>de</strong> unaumento <strong>de</strong> la presión intrafolicular.En el momento <strong>de</strong> la ovulación secompleta la división meiótica y se libera elprimer cuerpo polarEl oocito se expulsa ro<strong>de</strong>ado por la zonapelúcida y por una o mas capas <strong>de</strong> célulasfoliculares que rápidamente se colocan enforma <strong>de</strong> corona radiada.Fase lúteaEn las mujeres que han ovulado, loselementos residuales <strong>de</strong>l folículo forman elcuerpo lúteo que posee todas las enzimasesteroidogénicas <strong>de</strong> la serie P450.Luego <strong>de</strong> la salida <strong>de</strong>l óvulos, loscapilares y fibroblastos <strong>de</strong>l estromacircundante proliferan y penetran en lalámina basal aumentando su vascularizacióny las células murales <strong>de</strong> la granulosa sufrenel proceso <strong>de</strong> luteinización.
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 17La vascularización aumentada <strong>de</strong>esta estructura es muy importante ya queaporta abundante cantidad <strong>de</strong> LDL a lascélulas <strong>de</strong> la granulosa luteinizadas, y <strong>de</strong>allí se surten estas células para obtener elcolesterol para sintetizar progesterona.Las concentraciones pico <strong>de</strong>progesterona y estradiol logrados durante lafase lútea media constituyen el período <strong>de</strong> 3días, durante el cuál se produce laimplantación, si ha habido fertilización <strong>de</strong>lóvuloEl cuerpo lúteo duraaproximadamente 14 ± 2 días, sufriendoluego una regresión espontánea que loconvierte en la cicatriz avascular<strong>de</strong>nominada cuerpo albicans. La LH es lahormona central en la mantención <strong>de</strong>l cuerpolúteo.Los estrógenos y progesteronaproducidos por el cuerpo lúteo soninhibidores <strong>de</strong> la secreción e FSH, LH yGnRH.Si no ha habido fertilización <strong>de</strong>lóvulo, se produce la regresión <strong>de</strong>l cuerpolúteo, disminuyendo abruptamente lasecreción <strong>de</strong> estrógeno, progesterona einhibina e iniciándose un nuevo ciclo <strong>de</strong>secreción <strong>de</strong> FSH-LH (estimulados por GnRHhipotalámica) y por lo tanto un nuevo cicloovárico.Se consi<strong>de</strong>ra que la foliculogénesis,comienza durante la fase lútea tardía <strong>de</strong>lciclo previo y continúa durante la fasemenstrual, que se correspon<strong>de</strong> en el ovariocon la transición lúteo-folicular.En esta fase lútea tardía, <strong>de</strong>l cicloprevio, la lisis <strong>de</strong>l cuerpo lúteo asociado auna rápida disminución <strong>de</strong> los niveles <strong>de</strong>inhibina, estradiol y progesterona, permitenel aumento <strong>de</strong> la secreción <strong>de</strong> FSHaproximadamente 2 días antes <strong>de</strong>l comienzo<strong>de</strong> la menstruación, que ya comenzaría atener efecto para el reclutamiento <strong>de</strong>folículos para el nuevo cicloOtras sustancias que podrían jugarun papel modulador a nivel ovárico son:• Somatomedina o IGF-1, las células <strong>de</strong> lagranulosa ovárica expresan el gen <strong>de</strong>somatomedina y presentan receptorespara ella ,al igual que las células <strong>de</strong> lateca y las células intersticiales. Sepostula que esta hormona, regulada porSTH, podría tener importancia para el<strong>de</strong>sarrollo puberal.• Factor <strong>de</strong> crecimiento epidérmico (EGF) :ejerce efectos sobre la proliferación ydiferenciación <strong>de</strong> las células <strong>de</strong> lagranulosa ya que EGF y TGFα soninhibidores <strong>de</strong> la diferenciación <strong>de</strong> lascélulas <strong>de</strong> la granulosa porgonadotrofinas• TGFβ : sería producido a nivel <strong>de</strong> células<strong>de</strong> la teca e intersticiales, pero aún no sesabe su función.• hFGF: Factor básico <strong>de</strong>rivado <strong>de</strong>fibroblastos: : las células <strong>de</strong> la teca eintersticiales tienen receptores para estefactor que en <strong>trabajos</strong> experimentales ha<strong>de</strong>mostrado ser un potente atenuador <strong>de</strong>la síntesis <strong>de</strong> andrógenos-gonadotrofina<strong>de</strong>pendiente.• IL1: interleuquina 1: en formaexperimental se ha <strong>de</strong>mostrado que IL1produce inhibición <strong>de</strong> la luteinización <strong>de</strong>células <strong>de</strong> granulosa porcinaEn la menopausia, a pesar <strong>de</strong> lasaltas concentraciones <strong>de</strong> gonadotrofinascirculantes , el ovario se presenta como unaestructura atrófica, <strong>de</strong> superficie arrugada,con un peso <strong>de</strong> menos <strong>de</strong> 10 g.Presenta una médula <strong>de</strong> mayortamaño que la corteza y presenta loscuerpos albicans atravesados por vasosesclerosados.Esta estructura aún conservacarácter hormonal y secretaandrostenediona y testosterona.Los estrógenos (fundamentalmenteestrona) <strong>de</strong>tectables en la mujerpostmenopáusica <strong>de</strong>rivan <strong>de</strong> unaaromatización extraovárica <strong>de</strong>, sobre todo laandrostenediona suprarrenalUTEROTiene como función alojar y nutrir alfeto en <strong>de</strong>sarrollo y evacuarlo en el momentoa<strong>de</strong>cuado.Este órgano es blanco <strong>de</strong> lashormonas esteroi<strong>de</strong>a ováricas.
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 18Al inicio <strong>de</strong>l ciclo sexual el úterosufre el <strong>de</strong>sprendimiento <strong>de</strong> la capafuncional endometrial conservando su capabasal ( Fase menstrual).El aumento <strong>de</strong> estrógenos durante laprimera parte <strong>de</strong>l ciclo ovárico, produce unintenso efecto proliferativo sobre elendometrio, aumentando sus células yglándulas.(Fase proliferativa)El efecto estrogénico es intensosobre la secreción <strong>de</strong> moco cervical queadquiere una característica acuosa, elásticay filante.Los estrógenos estimulan lahidratación, con un gran aumento <strong>de</strong> laconcentración <strong>de</strong> sodio en el moco cervical.También aumentan su contenido enácido siálico ( que juega un papelimportante en la capacidad <strong>de</strong> receptividad<strong>de</strong>l moco cervical para el ingreso <strong>de</strong> losespermatozoi<strong>de</strong>s) ,por medio <strong>de</strong> laregulación <strong>de</strong> las enzimas fucosidasa ysialiltransferasa.Los filamentos <strong>de</strong> moco se alinean alo largo <strong>de</strong>l cuello uterino formandoconductos que ayudan a guiar a losespermatozoi<strong>de</strong>s en dirección apropiadahacia el úteroDespués <strong>de</strong> la ovulación, el efectopredominante sobre el útero es ejercido porla progesterona.Esta hormona produce e<strong>de</strong>matización<strong>de</strong> las células endometriales e induce enellas el <strong>de</strong>pósito <strong>de</strong> moco, lípidos yglucógeno.También estimula la irrigaciónsanguínea y disminuye el moco cervical quese hace grueso y no elástico ( Fasesecretoria o progestacional ó luteínica).Si no hubo fecundación <strong>de</strong>l óvulo, laatresia <strong>de</strong>l cuerpo amarillo ovárico, produceuna drástica disminución <strong>de</strong> la secreción <strong>de</strong>estrógenos y progesterona.La falta <strong>de</strong> estas hormonas quita elsustento trófico al <strong>de</strong>sarrollo endometrialuterino y esta capa es eliminada .Se estima que los vasos uterinos secontraen por acción <strong>de</strong> prostaglandinasproducidas en este período, quitándoleirrigación al endometrio. Se elimina la capafuncional endometrial durante lamenstruación y con ello se da inicio a unnuevo ciclo.En mujeres don<strong>de</strong> la ovulación esproducida por administración <strong>de</strong>gonadotrofinas , hay aparentemente unapérdida <strong>de</strong>l control fino <strong>de</strong> la liberación <strong>de</strong>FSH con la producción <strong>de</strong> ovulacionesmúltiplesPREGUNTAS DE AUTOEVALUACIONPregunta Nº 1 : (Tache lo que correspon<strong>de</strong>)Se producen cambios cíclicos en el ovariocomo reacción a las variaciones cíclicas <strong>de</strong> lasgonadotrofinas <strong>de</strong> la hipófisis ANTERIOR - POSTERIOR.Dichas gonadotrofinas son : ESTROGENOSLUTEINIZANTE - PROGESTERONA - HORMONAFOLÍCULO ESTIMULANTE - PROLACTINA.Pregunta Nº 2 : “La hormona luteinizante LO MISMOQUE - PERO NO la hormona estimulante <strong>de</strong> losfolículos ; es una glucoproteina pequeña con efectosespecíficos importantes sobre los ovarios LO MISMOQUE - PERO NO sobre los testículos.”Pregunta Nº 3 : La ovulación se acompaña <strong>de</strong> expulsión<strong>de</strong> un óvulo hacia UTERO - CAVIDAD ABDOMINALaproximadamente 14 ; 28 DÍAS <strong>de</strong>spués <strong>de</strong> la iniciación<strong>de</strong> la menstruación en el ciclo femenino <strong>de</strong> 14 -28 DÍASen promedio.Pregunta Nº 4 : Se produce una secreción máxima, enpico, <strong>de</strong> LH DOS DÍAS ANTES - DESPUÉS - 18HORAS ANTES - DESPUÉS <strong>de</strong> la ovulación.Pregunta Nº 5 : El incremento <strong>de</strong> la secreción endocrina<strong>de</strong>l cuerpo lúteo AUMENTA - INHIBE la secreción <strong>de</strong>LH lo mismo que - pero no la <strong>de</strong> FSH.Pregunta Nº 6 : Las concentraciones CRECIENTES -DECRECIENTES <strong>de</strong> ESTROGENOS -PROGESTERONA - GONADOTROFINAS durante lapubertad son la causa más directa <strong>de</strong>l crecimiento <strong>de</strong> lostejidos <strong>de</strong> los órganos sexuales femeninos.Pregunta Nº 7 : El efecto más importante <strong>de</strong> laprogesterona consiste en fomentar EL DEPOSITO DEGRASA - LA MITOSIS - LOS CAMBIOSSECRETORIOS en el endometrio.
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 19Pregunta Nº 8 : La fase proliferativa o FASEESTROGENICA - FASE PROGESTACIONAL <strong>de</strong>l cicloendometrial se produce en relación con LA PRIMERAPARTE - LA PARTE MEDIA - LA PARTE FINAL <strong>de</strong>lciclo ovárico.Pregunta Nº 9 : Haga un esquema relacionando lossiguientes parámetros:1. curva <strong>de</strong> secreción mensual <strong>de</strong> FSH y LH.2. curva <strong>de</strong> secreción mensual <strong>de</strong> estrógenos yprogesterona.3. ciclo ovárico.4. ciclo uterino.Utilice un esquema <strong>de</strong> ciclo sexual <strong>de</strong> 28 días<strong>de</strong> duración .PREGUNTAS A CONTESTAR DURANTE LAAUTOINSTRUCCIONDefina a que se <strong>de</strong>nomina ciclo sexualfemenino y analice las modificaciones que seproducen en los órganos <strong>de</strong>l aparato genitalfemenino, mamario y gónadas durante elmismo.Haga un esquema que represente elmecanismo <strong>de</strong> integración hipotálamo -hipófiso - gonadal y su mecanismoregulatorioRepresente en un sistema <strong>de</strong> coor<strong>de</strong>nadas lassecreciones hormonales implicadas en laproducción <strong>de</strong> la ovulación y <strong>de</strong>scriba loscambios que producen dichas hormonas en elovario. Tome <strong>de</strong> mo<strong>de</strong>lo un ciclo sexual <strong>de</strong>28 días.Represente es un esquema <strong>de</strong> coor<strong>de</strong>nadaslas secreciones hormonales que continúan alproceso <strong>de</strong> ovulación , hasta el momento <strong>de</strong>la menstruación. Analice que funcióncumplen.Defina a que se <strong>de</strong>nominan hormonassexuales femeninas, su lugar <strong>de</strong>producción y su función en las distintasetapas <strong>de</strong> la vida <strong>de</strong> la mujer.
CATEDRA Nº 1 DE FISIOLOGIA 2006 20CORRELACION ENDOCRINA EN ELHOMBRE(Dra. Susana Leyes <strong>de</strong> Lagraña)OBJETIVOS:Que el alumno explique los componentes <strong>de</strong>l sistemareproductor masculino y su funcionamiento.Que el alumno refiera la función interrelacionada <strong>de</strong>los distintos órganos que lo integran.Que el alumno i<strong>de</strong>ntifique la repercusión somática ypsíquica <strong>de</strong> la función sexual en el hombre.Que el alumno <strong>de</strong>talle los servomecanismos <strong>de</strong>regulación entre los distintos niveles <strong>de</strong> control <strong>de</strong> lafunción <strong>de</strong>l sistema reproductor.También se postula la existencia <strong>de</strong> unainhibina que sería producida por las células<strong>de</strong> Sértoli y que sería capaz <strong>de</strong> inhibir laliberación hipofisaria <strong>de</strong> FSH.RESEÑA EMBRIOLÓGICA:El aparato genital y el aparato urinarioprovienen <strong>de</strong> un pliegue mesodérmicocomún, e inicialmente <strong>de</strong>sembocan en unacavidad común: la cloaca.Con el <strong>de</strong>sarrollo y diferenciación anterior,sus productos finales, el semen y la orina,se eliminan a través <strong>de</strong> un conducto común:la uretra peniana. (Fig. Nº 1)Las gónadas son las estructurasencargadas <strong>de</strong> producir hormonas sexuales y<strong>de</strong> formar las células <strong>de</strong>stinadas a lareproducción.En el hombre se <strong>de</strong>nominan testículos yse encuentran localizadas en la bolsaescrotal.Si bien el sexo genético se <strong>de</strong>termina enel momento <strong>de</strong> la concepción y el sexogonadal esta <strong>de</strong>finido ya a nivel fetal, laplenitud morfológica y funcional <strong>de</strong> lasgónadas no se alcanza hasta la pubertad,momento a partir <strong>de</strong>l cual se expresan susfunciones reproductora y hormonal.Ambas funciones están bajo el control <strong>de</strong> lasgonatrofinas: FSH y LH hipofisarias,predominando la FSH sobre la funciónespermatogénica, y la LH sobre la funciónhormonal <strong>de</strong> producción <strong>de</strong> testosterona yuna pequeña cantidad <strong>de</strong> estrógenosAdicionalmente, existe un nivel <strong>de</strong> controlhipotalámico, hipofisario y gonadal estarelacionado e interregulado por mecanismos<strong>de</strong> retroalimentación positivos y negativosque se realizan <strong>de</strong> modo continuo en elhombre, <strong>de</strong>s<strong>de</strong> la pubertad.La testosterona mediante un mecanismo <strong>de</strong>retroacción larga, <strong>de</strong> tipo negativo, regula lasecreción hipofisaria <strong>de</strong> LH y <strong>de</strong> GnRH.Se postula existencia <strong>de</strong> una retroaccióncorta negativa hipofisaria – hipotalámica.. Fig. Nº 1: <strong>de</strong>terminación ydiferenciación sexual masculinaGÓNADAS:El sexo genético <strong>de</strong>l embrión es <strong>de</strong>terminadoen el momento <strong>de</strong> la fecundación por launión <strong>de</strong> un cromosoma X con un cromosomaY. (Fig. Nº 1)Pero las gónadas adquieren caracteresmorfológicos masculinos en la séptimasemana <strong>de</strong> <strong>de</strong>sarrollo intrauterino.Su primera manifestación la constituyen lospliegues o crestas genitales, las cuales sonpobladas por las células germinalesprimordiales provenientes <strong>de</strong>l saco vitelino.Esta estructura constituye los cordonessexuales primitivos, indiferenciados en lasprimeras semanas.Si el cariotipo <strong>de</strong>l embrión es XY, porinfluencia <strong>de</strong>l cromosoma Y, los cordones
CATEDRA Nº 1 DE FISIOLOGIA 2006 21sexuales proliferan y se introducen en lamédula gonadal diferenciándose en cordonesmedulares, (que en la pubertad se canalizandando origen a las testículos seminíferos).Hacia la 8ª semana <strong>de</strong>l <strong>de</strong>sarrollointrauterino, las células <strong>de</strong> Leydig fetalesestimuladas por la GCH comienzan aproducir testosterona que:1. Estimula el <strong>de</strong>sarrollo <strong>de</strong>l epídídimo,vasos <strong>de</strong>ferentes, vesículas seminales yconductos eyaculadores (a partir <strong>de</strong>lconducto <strong>de</strong> Wolff).introducen y forman un cordón epitelial cortoque luego se canaliza.ETAPA ADULTA:En el adulto el sistema reproductormasculino consta <strong>de</strong> dos gónadas: lostestículos, alojados en el escroto; losconductos excretores (vasos rectos, retetestis, conductos <strong>de</strong>ferentes, ampolla,conductor eyaculador y uretra) yglándulas accesorias: próstata, vesículasseminales y glándulas bulbouretrales.(Fig. N° 2)2. Promueve la virilización <strong>de</strong>l senourogenital y <strong>de</strong>l tubérculo genital, dandolugar a la formación <strong>de</strong> la próstata,uretra masculina, pene y escroto.Hacia 10ª semana se pue<strong>de</strong> reconocer elfeto masculino por la apariencia <strong>de</strong> susgenitales externos.3. Estimula el <strong>de</strong>scenso <strong>de</strong> los testículoshacia el 7º mes.El número y actividad <strong>de</strong> las células <strong>de</strong>Leydig disminuye hacia el final <strong>de</strong>l tercertrimestre <strong>de</strong>l embarazo, <strong>de</strong>clinandobruscamente hasta la edad puberal.Se reconoce dos andrógenos principales quecontrolan la diferenciación y <strong>de</strong>sarrollo <strong>de</strong>los órganos sexuales accesorios:a) La testosterona: que viriliza losconductos <strong>de</strong> Wolff.b) La dihidrotestosterona: que controla ladivisión <strong>de</strong>l seno urogenital y <strong>de</strong>ltubérculo genital para dar lugar a laformación <strong>de</strong>: próstata, glándulas <strong>de</strong>Cowper y genitales externos.GENITALES EXTERNOSPor influencia <strong>de</strong> los andrógenos fetales seproduce alargamiento <strong>de</strong>l tubérculo genital.Simultáneamente los pliegues uretralesforman la uretra peneana al final <strong>de</strong>l 3ºmes.El meato urinario se forma en el 4º mes, porproliferación <strong>de</strong> células <strong>de</strong>l glan<strong>de</strong> que seFig. N° 2: sistema reproductormasculinoLos testículos están alojados en el escroto,bolsa dérmica que contiene receptores <strong>de</strong>calor y fibras nerviosas vasoconstrictoras yvasodilatadoras.El escroto contribuye al mantenimiento <strong>de</strong> latemperatura testicular por <strong>de</strong>bajo <strong>de</strong> lacorporal, condición necesaria para unaespermatogénesis normal.La función testicular está implicada en:espermatogénesisproducción hormonalESPERMATOGENESIS:Es el proceso <strong>de</strong> formación <strong>de</strong> los gametosmasculinos o espermatozoi<strong>de</strong>s, a partir <strong>de</strong>precursores germinales llamadosespermatogonias.Estas se encuentran en la base <strong>de</strong>l túbuloseminífero y permanecen en reposo hasta lapubertad.
CATEDRA Nº 1 DE FISIOLOGIA 2006 22A partir <strong>de</strong> esa etapa se divi<strong>de</strong>nmitoticamente para reponer su número yformar células llamadas espermatogonias Bque se diferencian a espermatozoi<strong>de</strong>s.Esto se logra por medio <strong>de</strong> un proceso queincluye 2 formas <strong>de</strong> diferenciación:meiosis, en la que se reduce el númerocromosómico y se reor<strong>de</strong>na (recombina) elmaterial genético, y espermiogénesis,proceso regulado por testosterona don<strong>de</strong> seproducen cambios estructurales:compactación <strong>de</strong> la cromatina, aparición <strong>de</strong>lacrosoma (aparato <strong>de</strong>stinado a lapenetración <strong>de</strong>l espermatozoi<strong>de</strong> en el óvulo),formación <strong>de</strong> la pieza intermediaconteniendo mitocondrias, cambio <strong>de</strong> forma,con<strong>de</strong>nsación <strong>de</strong>l citoplasma y <strong>de</strong>sarrollo <strong>de</strong>lflagelo, estructura <strong>de</strong>stinada a dotar <strong>de</strong>movilidad al espermatozoi<strong>de</strong> maduro. (Fig.N° 3)Fig. N° 3: espermatogénesisFinalizada la fase <strong>de</strong> diferenciación, losespermatozoi<strong>de</strong>s salen <strong>de</strong>l epitelioseminífero hacia el epidídimo, don<strong>de</strong>adquieren movilidad. Este paso esfundamental ya que <strong>de</strong>termina su capacidadpara alcanzar e interaccionar con el oocito.En el espermatozoi<strong>de</strong> <strong>de</strong>bemos distinguirtres partes: cabeza; cuello y cola. (Fig.N° 4)La cabeza esta formada por el núcleo <strong>de</strong>esta célula, ro<strong>de</strong>ado <strong>de</strong> la membranacitoplasmática.En su parte anterior presente una cubiertagruesa: el acrosoma que tiene un alto contenido <strong>de</strong>enzimas proteolíticas; entre las cuales mencionamos a lahialuronidasa porque juega un papel trascen<strong>de</strong>nte en elproceso <strong>de</strong> la fecundación.La cola presenta:a) Un esqueleto central, el axonema, cubierto por unamembrana.b) El cuerpo <strong>de</strong> la cola, que contiene una cantidad <strong>de</strong>mitocondrias que proveen energía para elmovimiento <strong>de</strong>l espermatozoi<strong>de</strong>, lo cual le permitea este <strong>de</strong>splazarse a una velocidad <strong>de</strong> 1 a 4mm/minuto en el tracto genital femenino.Regulación <strong>de</strong> la espermatogénesis:Son fundamentales las uniones estrechasentre células <strong>de</strong> Sertoli y la membrana basal<strong>de</strong>l túbulo seminífero, que forman la barrerahemato- testicular. Esta crea unmicroambiente aislado <strong>de</strong>l medio externo quetiene 2 partes: el compartimiento basal quecontiene las espermatogonias y elcompartimiento luminal que contiene lasespermáti<strong>de</strong>s y espermatozoi<strong>de</strong>s.Las células <strong>de</strong> Sertoli, bajo influencia <strong>de</strong>FSH, aumentan la síntesis proteica,aromatizan la testosterona proveniente <strong>de</strong>las células <strong>de</strong> Leydig para formar estradiol.También producen ABP (proteína fijadora <strong>de</strong>andrógenos) que se secreta a la luz tubular,don<strong>de</strong> asegura alta concentración <strong>de</strong>testosterona para la espermatogénesis, einhibina, hormona glucoproteica que actúasobre la hipófisis inhibiendo la FSH.CELULAS DE SERTOLI:Constituyen junto con las espermatogonias,los elementos epiteliales <strong>de</strong>l túbuloseminífero.Se hallan en contacto con la membranabasal <strong>de</strong>l túbulo y emiten prolongacioneshacia la luz.
CATEDRA Nº 1 DE FISIOLOGIA 2006 23Estas células proliferan durante lavida fetal y perinatal perdiendo sucapacidad <strong>de</strong> división en el adulto.Son células blanco <strong>de</strong> la FSH y tienenfunción secretora produciendo: factor <strong>de</strong>inhibición mullenariana, inhibina, proteínatransportadora <strong>de</strong> andrógenos y sistemas <strong>de</strong>conversión <strong>de</strong> pregnenolona a testosteronay <strong>de</strong> testosterona a dihidrotestosterona.TEJIDO INTERSTICIAL:Los túbulos seminíferos están incluidos enuna matriz <strong>de</strong> células intersticiales, vasossanguíneos y linfáticos.La célula predominante es la célula <strong>de</strong>Leydig, responsable <strong>de</strong> la esteroidogénesis.Esta se produce a partir <strong>de</strong>l colesterolproviene <strong>de</strong> la circulación o por síntesiscelular.La esteroidogénesis está regulada por la LHen el adulto y por la HGC en el feto.Los niveles <strong>de</strong> gonadotrofinas hipofisariasson muy bajos hasta los 10 años <strong>de</strong> edad,etapa en que comienza a aumentar, con elconsecuente aumento <strong>de</strong> andrógenos.Esto indica el inicio <strong>de</strong> la pubertadmasculina, hacia los 13 años.El inicio <strong>de</strong> la pubertad se atribuye a unproceso <strong>de</strong> maduración hipotalámica,postulándose que el hipotálamo prepuberalse inhibe ante las bajas concentraciones <strong>de</strong>andrógenos circulantes en ese período, loque bloquea la producción <strong>de</strong> GnRH.Al llegar a la pubertad, la maduraciónhipotalámica produce una disminución <strong>de</strong> lasensibilidad <strong>de</strong> las células productoras <strong>de</strong>GnRH a la testosterona circulante y se iniciala producción <strong>de</strong> la liberina hipotalámica <strong>de</strong>hormona luteinizante con liberación <strong>de</strong> LHa<strong>de</strong>nohipofisaria.Se estimula así la espermatogénesis ysecreción <strong>de</strong> hormonas sexuales,permitiendo el <strong>de</strong>sarrollo <strong>de</strong> la capacidadcompleta <strong>de</strong>l hombre para la reproducción.La función reproductora y hormonal semantiene durante toda la vida <strong>de</strong>l hombre,aunque habría una disminución hacia laquinta o sexta década <strong>de</strong> la vida, atribuible afactores psicosomáticos o a la repercusión<strong>de</strong> patologías propias <strong>de</strong> esa edad sobre elaparato sexual.HORMONAS SEXUALES:Andrógenos: la testosterona producida porlas células <strong>de</strong> Leydig bajo la influencia <strong>de</strong> laLH, es el andrógeno principal tanto en elhombre como en la mujer.En algunos órganos blanco como próstata,genitales externos y folículos pilosos lo hacemediante transformación a 5-DHT, <strong>de</strong>rivadomás potente que se une al receptor conmayor afinidad.Sobre el SNC su acción se produce poraromatización. Sobre otros tejidos comohipófisis, células <strong>de</strong> Sertoli, vesículasseminales, epidídimo y conductos <strong>de</strong>ferente,actúa sin modificaciones.Sus acciones se verifican <strong>de</strong>s<strong>de</strong> la vidaembrionaria, don<strong>de</strong> estimulan el <strong>de</strong>sarrollo<strong>de</strong> los conductos <strong>de</strong> Wolf y <strong>de</strong> los genitalesexternos.A partir <strong>de</strong> la pubertad estimulan elcrecimiento <strong>de</strong>l pene, escroto y próstata, lapigmentación escrotal, el crecimiento <strong>de</strong>lvello facial y corporal, el crecimientosomático, <strong>de</strong> la laringe y el aumento <strong>de</strong>masa muscular.También contribuye a mantener laespermatogénesis, y tiene un fuerte efectoanabólico que se manifiesta en el hueso, enlos glóbulos rojos y en la síntesis proteica.Sobre el sistema nervioso, actúa <strong>de</strong>finiendolas características conductuales <strong>de</strong>l varón, ysobre la hipófisis actúa sobre la LHEn el plasma, la testosterona circula unida auna beta globulina fijadora <strong>de</strong> andrógenos.La testosterona que no se fija a célulasblanco <strong>de</strong> metaboliza a nivel hepático aandrosterona que es conjugada y eliminadapor bilis u orina.EPIDIDIMO
CATEDRA Nº 1 DE FISIOLOGIA 2006 24Es un conducto espiralado don<strong>de</strong> seproducen las últimas etapas <strong>de</strong> maduración y<strong>de</strong>sarrollo <strong>de</strong>l espermatozoi<strong>de</strong>.El epitelio epididimario se caracteriza porser muy activo metabólicamente, realizandofunciones <strong>de</strong> absorción y secreción.Sintetiza y secreta hacia la luz:proteínascarnitinalípidosglicerolfosforilcolinacarbohidratosesteroi<strong>de</strong>s.Se <strong>de</strong>mostró la presencia <strong>de</strong> dos enzimasespecíficas en el epidídimo: lagalactosiltransferasa y la alfa lactalbúmina,las cuales producen glucosilación <strong>de</strong> lasuperficie <strong>de</strong> los espermatozoi<strong>de</strong>s, a medidaque progresan por el epidídimo.Este proceso es fundamental para elreconocimiento por parte <strong>de</strong>lespermatozoi<strong>de</strong>, <strong>de</strong>l tracto genital femeninoy su posibilidad <strong>de</strong> unirse al óvulo.La carnitina no se sintetiza en el epidídimo,no se acumula allí luego <strong>de</strong> su absorción<strong>de</strong>s<strong>de</strong> la sangre. Esta molécula interviene enel metabolismo <strong>de</strong> los ácidos grasos ytendría una función vital para la mantención<strong>de</strong> la integridad <strong>de</strong> la cubierta <strong>de</strong>espermatozoi<strong>de</strong>.La función epididimaria <strong>de</strong>pen<strong>de</strong> <strong>de</strong> lashormonas testiculares y dihidrotestosterona;también <strong>de</strong> la prolactina que actuaríaaumentando la captación <strong>de</strong> testosterona.VASOS DEFERENTES:Son órganos tubulares <strong>de</strong> 35 a 45 cm <strong>de</strong>longitud. Se extien<strong>de</strong>n <strong>de</strong>s<strong>de</strong> la cola <strong>de</strong>lepidídimo hasta la <strong>de</strong>sembocadura <strong>de</strong> lasvesículas seminales, don<strong>de</strong> se dilatanformando una ampolla.A partir <strong>de</strong> allí se llama conductoeyaculador, pasa a través <strong>de</strong> la próstata ytermina en a uretra peneana. Su funciónprincipal es transportar y almacenar a losespermatozoi<strong>de</strong>s en la ampolla.PROSTATA:Es la glándula sexual accesoria más gran<strong>de</strong>.En el adulto joven pesa unos 20 gramos.Su parénquima está formado por 30 a 40glándulas túbulo – alveolares cuyosconductos excretorios se abren en el piso ysuperficie lateral <strong>de</strong> la uretra. Estasglándulas están construidas por célulassecretorias y células <strong>de</strong> reserva.Tiene dos funciones principales:1. Función esfinteriana: la base prostática selocaliza en íntima relación con el cuello vesical,entre los dos esfínteres <strong>de</strong> la uretra. El músculo lisoque ro<strong>de</strong>a a la próstata se fun<strong>de</strong> con el esfíntervesical externo, tiene relación con lacontinencia urinaria y probablemente estaimplicado en la emisión <strong>de</strong> orina.2. Función secretoria: 15 a 30 % <strong>de</strong>l volumeneyaculado, constituido por 0,2 a 1,5 ml <strong>de</strong> líquidolevemente alcalino, <strong>de</strong> aspecto lechoso, esproducido por la próstata. Esta secreción es rica enfosfatasa ácida, poliaminas e incluye espermina,espermidina y putrescina. Por oxidación producen eltípico olor a almizcle <strong>de</strong>l semen.La actividad secretoria esta bajo el control <strong>de</strong> latestosterona que ejerce su acción al convertirse enla células blanco en dihidrotestosterona.VESICULAS SEMINALES:Son glándulas pares <strong>de</strong> forma filiforme yespiralada que <strong>de</strong>sembocan en la ampolla<strong>de</strong>l conducto <strong>de</strong>ferente homolateral,producen un líquido viscoso, amarillento <strong>de</strong>pH alcalino, que constituye el 70% <strong>de</strong>lEyaculado.Durante la excitación sexual, las vesículasseminales se vuelven tensas y se contrae sucopa <strong>de</strong> músculo liso en forma peristáltica,<strong>de</strong>scargando su secreción a los conductoseyaculadores y a la uretra.El líquido seminal está constituido por:FructosaSorbitolCitratosInositolColinesterasaFibrinógeno
CATEDRA Nº 1 DE FISIOLOGIA 2006 25Prostaglandinas (que cumplirían la función <strong>de</strong>permitir el paso <strong>de</strong>l espermatozoi<strong>de</strong>).RESPUESTA SEXUAL MASCULINA:Consta <strong>de</strong> cuatro fases:1. Excitación2. Meseta3. Orgasmo4. ResoluciónSe inicia con la erección, fenómenoinvoluntario que obe<strong>de</strong>ce a reflejospsicógenos o estimulación <strong>de</strong>l glan<strong>de</strong>,escroto, cuerpo <strong>de</strong>l pene o región anal,producidos por impulsos parasimpáticos queviajan por vía nervios erectores <strong>de</strong>l plexosacro.La erección se produce por:- Cierre <strong>de</strong> anastomosis arteriovenosas entre laarteria profunda <strong>de</strong>l pene y el retorno venosoperiférico.- Constricción <strong>de</strong> las arteriolas peneanas que facilitala entrada <strong>de</strong> sangre a los cuerpos cavemosos.- Bloqueo <strong>de</strong>l drenaje sanguíneo por constricción <strong>de</strong>las venas peneanas.Cuando el pene se encuentra en estado <strong>de</strong>relajación, los músculos circulares ylongitudinales <strong>de</strong> las almohadillas arterialesestán contraídos y disminuyen el volumen <strong>de</strong>su luz: esto impi<strong>de</strong> la entrada <strong>de</strong> sangre enlos cuerpos cavernosos.Durante la erección, las fibras se relajan eingresan 20 a 25 ml <strong>de</strong> sangre en loscuerpos cavernosos, que adquieren rigi<strong>de</strong>zpor estar ro<strong>de</strong>ados por una cápsulainextensible.Durante la fase <strong>de</strong> meseta y orgasmo, seproduce los fenómenos <strong>de</strong> emisión yeyaculación.La emisión resulta <strong>de</strong> la acumulación <strong>de</strong>semen en la uretra prostática.La actividad refleja simpática producecontracción <strong>de</strong>l músculo liso <strong>de</strong> las vesículasseminales, próstata y vasos <strong>de</strong>ferentes y elingreso <strong>de</strong> sus secreciones en la uretra.En ese momento se produce el cierre <strong>de</strong>lcuello vesical para evitar el flujo retrógrado<strong>de</strong>l semen hacia el interior <strong>de</strong> la vejiga.Luego <strong>de</strong> la emisión <strong>de</strong> semen se produce laeyaculación, por contracción rítmica <strong>de</strong> losmúsculos bulbocavernosos oisquicavernosos, y <strong>de</strong>l esfínter uretralexterno.La eyaculación se producen en fracciones,en forma bien <strong>de</strong>finida, que permite que laemisión <strong>de</strong> espermatozoi<strong>de</strong>s a partir <strong>de</strong>lconducto <strong>de</strong>ferente y ampolla, se mezcle conlas secreciones <strong>de</strong> las glándulas <strong>de</strong> Cowper,uretrales y prostática.ESTUDIO FUNCIONAL:Espermograma:En el estudio <strong>de</strong> la pareja estéril, el análisis<strong>de</strong> semen es una <strong>de</strong> las pruebas básicaspara la elaboración <strong>de</strong> un diagnóstico enreproducción.Para el estudio <strong>de</strong>l semen se empleantécnicas citológicas, bioquímicas einmunológicas entre otras. Cada una analizadiferentes parámetros que pue<strong>de</strong>n afectar ala capacidad potencial <strong>de</strong> fertilización <strong>de</strong>lvarón.Por ejemplo, la medida <strong>de</strong>l volumen seminalevalúa la capacidad <strong>de</strong> las glándulassexuales accesorias para proporcionarsuficiente cantidad <strong>de</strong> fluido seminal,mientras que el recuento espermáticocuantifica la capacidad <strong>de</strong>l testículo paraproducir espermatozoi<strong>de</strong>s.COMPONENTES DEL EYACULADO:El eyaculado es una mezcla <strong>de</strong> distintoscomponentes proce<strong>de</strong>ntes <strong>de</strong>l testículo, <strong>de</strong>los segmentos <strong>de</strong> la vía seminal y <strong>de</strong> lasglándulas anejas:espermatozoi<strong>de</strong>s,secreción <strong>de</strong> glándulas bulbouretrales (10 %),secreción prostática (30 %)y <strong>de</strong> vesículas seminales (60 %)En general es expulsado en cuatrofracciones diferentes:
CATEDRA Nº 1 DE FISIOLOGIA 2006 261. La fracción preeyaculatoria,proce<strong>de</strong>nte <strong>de</strong> las glándulas <strong>de</strong>Cowper y Litré, es <strong>de</strong> consistenciamucosa y está exenta <strong>de</strong>espermatozoi<strong>de</strong>s.2. Fracción previa, completamentefluida, proviene <strong>de</strong> la próstata, ytampoco presenta espermatozoi<strong>de</strong>s.Supone <strong>de</strong>l 13 al 33% <strong>de</strong>l total <strong>de</strong>leyaculado, posee un pH ácido y secaracteriza por su elevadaconcentración <strong>de</strong> fosfatasa ácida yácido cítrico.3. Fracción principal, que consta tanto<strong>de</strong> elementos líquidos comogelatinosos, es la más rica enespermatozoi<strong>de</strong>s y proce<strong>de</strong> <strong>de</strong>lepidídimo, <strong>de</strong>ferente y ampolla<strong>de</strong>ferencial.4. Fracción terminal, gelatinosa, es lamás abundante <strong>de</strong>l eyaculado, 50-80%. Proce<strong>de</strong> <strong>de</strong> las vesículasseminales, por lo que es rica enfructosa. Posee pH alcalino yespermatozoi<strong>de</strong>s, aunque la mayoríainmóviles.CONDICIONES INICIALES DELESTUDIOLa muestra <strong>de</strong> semen <strong>de</strong>be recogerse tras unmínimo <strong>de</strong> 48 horas <strong>de</strong> abstinencia sexual,pero no <strong>de</strong>spués <strong>de</strong> 8 días, preferiblementepor masturbación.El intervalo entre el momento <strong>de</strong> laeyaculación y el análisis, que no <strong>de</strong>be sernunca mayor <strong>de</strong> 45 minutos.Licuación: Inmediatamente tras laeyaculación, el líquido seminal coagula yposteriormente se licua <strong>de</strong>ntro <strong>de</strong> los 5-40minutos a temperatura ambiente. La enzimaresponsable <strong>de</strong> la licuación proviene <strong>de</strong> lapróstata, mientras que la <strong>de</strong> la coagulaciónproce<strong>de</strong> <strong>de</strong> la vesícula seminal.Volumen: El volumen <strong>de</strong> líquido seminaltiene un promedio <strong>de</strong> 2 a 5,5 ml. Laabstinencia sexual prolongada pue<strong>de</strong> darlugar a volúmenes seminales mayores. Lacontribución prostática y <strong>de</strong>l epidídimo allíquido seminal no exce<strong>de</strong> por lo general <strong>de</strong>1 ml. En consecuencia, el volumen seminales fundamentalmente función <strong>de</strong> la actividad<strong>de</strong> las vesículas seminales.Ph: en condiciones normales, oscila entre7,2 y 8,0Número: entre 20 y 250 millones/ml. Valoresmenores <strong>de</strong> 20 mill/ml se <strong>de</strong>nominanoligozoospermia.Otros elementos celulares: El eyaculadocontiene otras células que no sonespermatozoi<strong>de</strong>s. Entre éstas se incluyen lascélulas epiteliales proce<strong>de</strong>ntes <strong>de</strong>l tractoreproductivo, células <strong>de</strong> espermiogénesis yleucocitosLa movilidad espermática constituye uno<strong>de</strong> los parámetros fundamentales paravalorar la calidad <strong>de</strong>l eyaculado.Los espermatozoi<strong>de</strong>s son capaces <strong>de</strong> exhibirdiferentes patrones <strong>de</strong> movimientoadaptados a sus necesida<strong>de</strong>s funcionales:<strong>de</strong>ntro <strong>de</strong>l plasma seminal muestrantrayectorias lineales que son las quefisiológicamente mostrarían al penetrar en elmoco cervical.Al iniciar su ascenso por el tractoreproductivo femenino, sus características<strong>de</strong> movilidad cambian.Cuando se produce la capacitación, o a lolargo <strong>de</strong> este proceso, aumenta la amplitud<strong>de</strong> la onda flagelar, originándosetrayectorias que se muestran con un mayor<strong>de</strong>splazamiento lateral <strong>de</strong> la cabeza.En estas condiciones, el espermatozoi<strong>de</strong> sedice que se encuentra en un estado"transicional"Cuando tiene lugar verda<strong>de</strong>ramente lacapacitación, la amplitud <strong>de</strong> la onda flagelarcomienza a hacerse asimétrica, dando lugara un movimiento no progresivo y vigorosoconocido como hiperactivación.Esta forma <strong>de</strong> movilidad es típica <strong>de</strong> losespermatozoi<strong>de</strong>s totalmente capacitados, yse piensa que proporciona las fuerzasnecesarias para que el espermatozoi<strong>de</strong>pueda atravesar la zona pelúcida.
CATEDRA Nº 1 DE FISIOLOGIA 2006 27La movilidad <strong>de</strong>pen<strong>de</strong> tanto <strong>de</strong> factoresintrínsecos (estructura <strong>de</strong>l flagelo, activida<strong>de</strong>nzimática), como <strong>de</strong> factores extrínsecos(composición bioquímica <strong>de</strong>l medioextracelular en el que se encuentra elespermatozoi<strong>de</strong>, plasma seminal, mococervical, etc.).2. Que cambios ocurren en el varón al llegar a lapubertad? (generales y sobre el aparato genital ygonadal).3. Describa las acciones <strong>de</strong> la testosterona.4. ¿A que se <strong>de</strong>nomina espermatogénesis?5. ¿Cuál es su producto final?; ¿Cuales son lascaracterísticas <strong>de</strong> un espermograma normal?La OMS aconseja aceptar una muestra<strong>de</strong> semen como normal en lo que serefiere a movilidad espermática,cuando al menos el 50% <strong>de</strong> losespermatozoi<strong>de</strong>s están dotados <strong>de</strong>movilidad progresiva, y al menos 30 %tienen movimientos rápidosMorfología espermática: su estudioadquiere importancia <strong>de</strong>s<strong>de</strong> la utilización <strong>de</strong>la fertilización in vitro, ya que el porcentaje<strong>de</strong> espermatozoi<strong>de</strong>s morfológicamentenormales es una <strong>de</strong> las variables que mejorse relaciona con el éxito <strong>de</strong> la fertilización<strong>de</strong> ovocitos.La cabeza <strong>de</strong> un espermatozoi<strong>de</strong> normal<strong>de</strong>be tener forma oval. La longitud es <strong>de</strong>entre 4 y 5,5 u, y la anchura <strong>de</strong> entre 2,5 y3,5 u. A<strong>de</strong>más <strong>de</strong>be poseer una regiónacrosomal bien <strong>de</strong>finida que compren<strong>de</strong>entre el 40 y 70% <strong>de</strong>l área <strong>de</strong> la cabeza y no<strong>de</strong>ben existir <strong>de</strong>fectos en el cuello,segmento intermedio o flagelo.Estudio bioquímico <strong>de</strong>l eyaculado.:a) Capacidad secretora <strong>de</strong> la próstata: Elcontenido <strong>de</strong> zinc, ácido cítrico y fosfatasaácida en semen nos da una buena medida <strong>de</strong>la secreción <strong>de</strong> esta glándula.b) Capacidad secretora <strong>de</strong> las vesículasseminales: La fructosa en semen refleja lafunción secretora <strong>de</strong> las vesículas seminalesc) Capacidad secretora <strong>de</strong>l epidídimo: losmarcadores epididimarios son L-carnitina yalfa-glucosidasa, que es específica <strong>de</strong>lepidídimo y más sensible.PREGUNTAS A CONTESTAR DURANTE LAAUTOINTRUCCION1. ¿Qué funciones gonadales regulan las hormonasFSH y LH?
CATEDRA Nº 1 DE FISIOLOGIA 2006 28FISIOLOGIA DE LOS REFLEJOS(Dra. Lilian Barrios y Dr. O. Poletti)La actividad refleja <strong>de</strong>l sistema nerviosointerviene en una gran cantidad <strong>de</strong>funciones <strong>de</strong>l individuo.Por ejemplo: en la regulación <strong>de</strong> la presiónarterial, regulación <strong>de</strong> la motilidad <strong>de</strong>l tubodigestivo, mantención <strong>de</strong> la postura,mantención <strong>de</strong>l tono muscular, mantención<strong>de</strong>l equilibrio, marcha, producción <strong>de</strong> tos,entre otros.Dicha actividad se constituye sobre unaestructura <strong>de</strong>nominada arco reflejo.DEFINICION DE ARCO REFLEJO:Se consi<strong>de</strong>ra al arco reflejo como la unidadfuncional <strong>de</strong> la actividad integrada <strong>de</strong>lsistema nervioso.Es una actividad involuntaria y pue<strong>de</strong> ser consciente ono.Dicho arco consta <strong>de</strong> los siguientescomponentes: (Fig. 1)Fig. 1: componentes <strong>de</strong> un arcoreflejo simple1. Receptor sensorial;2. Neurona aferente (<strong>de</strong> localización enganglio raquí<strong>de</strong>o o en estación centralintegradora);3. Neuronas intercalares ó interneuronas;4. Neurona eferente;5. Efector.1. Receptor Sensorial : pue<strong>de</strong> estarubicado en piel, músculo, tendones,articulaciones o vísceras; <strong>de</strong> acuerdo asu ubicación se clasifica en :• Receptores exteroceptivos: los queestán ubicados en piel o mucosas.• Receptores propioceptivos: los queestán en músculo, tendones oarticulaciones.• Receptores interoceptivos: los queestán en las vísceras.2. Neurona aferente: Por su prolongación<strong>de</strong>ntrítica se conecta con el receptor(Ver figuras Nº 1 y 2 en hojaposterior); su soma está ubicado en unganglio raquí<strong>de</strong>o o en un ganglio anivel craneal (<strong>de</strong>pendiendo <strong>de</strong> sulocalización) y su prolongación axonalpenetra en el sistema nervioso centralconectándose con interneuronas y / oneuronas motoras α , γ o vegetativas.3. Neuronas intercalares ó interneuronas:son neuronas cuya estructura totalestá ubicada en el sistema nerviosocentral, son muy numerosas y seestima que son las principalesencargadas <strong>de</strong> la función integradora<strong>de</strong> este sistema.4. Neurona eferente: es la neurona cuyaprolongación axónica se pone endirecto contacto con el efector paraproducir la respuesta refleja (neuronasmotoras α (alfa), γ (gamma) ovegetativas.5. Efectores: Producen las respuestasreflejas y pue<strong>de</strong>n ser:músculo esqueléticomúsculo lisoglándulasCLASIFICACION DE LOS REFLEJOS:La clasificación más usada enclínica divi<strong>de</strong> a los reflejos en:Reflejos Superficiales: Son aquellosque se inician a partir <strong>de</strong> fenómenosexteriores al organismo incidiendo sobrereceptores ubicados en piel y mucosas(calor, frío, presión).Compren<strong>de</strong>n reflejos tales como:abdominales,<strong>de</strong> succión,plantar,
CATEDRA Nº 1 DE FISIOLOGIA 2006 29cremasteriano, etc...Reflejos Profundos: Que compren<strong>de</strong>n aaquellos que inci<strong>de</strong>n sobre estructurasinternas <strong>de</strong>l cuerpo.Compren<strong>de</strong>n alos reflejos propioceptivosy reflejos visceroceptivos.Los reflejos propioceptivos se inician enreceptores ubicados en músculoesquelético, tendones, articulaciones yaparato vestibular.Compren<strong>de</strong>n reflejos tales como:reflejos <strong>de</strong> postura,tono muscular,equilibrio,marcha,risa,grito,llanto.Los reflejos visceroceptivos se originan enreceptores ubicados en las vísceras(reflejo <strong>de</strong>l seno carotí<strong>de</strong>o, reflejos <strong>de</strong>insuflación - <strong>de</strong>flación, etc.).REFLEJOS INVESTIGADOSUSUALMENTE EN SEMIOLOGIADEL SISTEMA NERVIOSO:Reflejo corneo: Se obtiene excitando oirritando la córnea con un algodoncillo,produce cierre <strong>de</strong>l ojo y su centro se hallaen la protuberancia.Reflejo maseterino: Se obtiene percutiendocon el martillo <strong>de</strong> reflejos sobre el mentón,con la boca abierta; la respuesta consisteen cierre <strong>de</strong> la boca y su centro se hallaen la protuberancia.Reflejo olecraneano: Se obtienepercutiendo con el martillo <strong>de</strong> reflejossobre el olécranon; la respuesta consisteen la flexión <strong>de</strong>l antebrazo sobre el brazoy su centro se halla en 5º y 6º segmentoscervicales.Reflejo tricipital: Se obtiene percutiendocon el martillo <strong>de</strong> reflejos sobre el tendón<strong>de</strong>l tríceps, su respuesta es una extensión<strong>de</strong>l antebrazo sobre el brazo y su centro sehalla en 6º y 7º segmentos cervicales.Reflejo radial: Se obtiene percutiendo conel martillo <strong>de</strong> reflejos sobre la apófisisestiloi<strong>de</strong>s <strong>de</strong>l radio, su respuesta es unaflexión y supinación <strong>de</strong>l antebrazo y sucentro se halla en 5º y 6º segmentoscervicales.Reflejo cubital: Se obtiene percutiendo conel martillo <strong>de</strong> reflejos sobre la apófisisestiloi<strong>de</strong>s <strong>de</strong>l cúbito; su respuesta consisteen pronación <strong>de</strong>l antebrazo con flexión <strong>de</strong>los <strong>de</strong>dos y su centro correspon<strong>de</strong> al 7º y8º segmentos cervicales.Reflejo cutáneo - abdominales superior einferior: Se obtienen produciendoestimulación <strong>de</strong> la pared abdominal en suparte superior e inferior con la punta <strong>de</strong> unalfiler o con la yema <strong>de</strong>l <strong>de</strong>do o con elmango <strong>de</strong>l martillo <strong>de</strong> reflejos pasándoloen forma perpendicular a la línea media;su respuesta consiste en una <strong>de</strong>sviación<strong>de</strong>l ombligo hacia el lado estimulado.El reflejo cutáneo - abdominal superiortiene su centro en 6º y 7º segmentosdorsales y el inferior en X, XI y XIIsegmentos dorsales.También se pue<strong>de</strong> tomar el reflejo en lazona abdominal media investigándose enese caso los segmentos dorsales 8º y 9º.Reflejo mediopubiano: Se investigapercutiendo con el martillo <strong>de</strong> reflejos enla sínfisis púbica; el enfermo <strong>de</strong>be estarcon los músculos separados y las piernassemiflexionadas.Da una respuesta doble:1. Contracción <strong>de</strong> los músculos abdominales;2. Aproximación <strong>de</strong> los muslos por contracción <strong>de</strong>los aductores. Su centro se halla entre la 10adorsal y L2.Reflejo cremasteriano: Se investiga porexcitación en la cara interna <strong>de</strong>l muslo enparte superior.Da como respuesta elevación <strong>de</strong>l testículo ycontracción <strong>de</strong>l oblicuo mayor en el hombre ycontracción <strong>de</strong>l oblicuo mayor en la mujer.Su centro se halla en los segmentos L1 y L2.
CATEDRA Nº 1 DE FISIOLOGIA 2006 30Reflejo patelar: Se pue<strong>de</strong> tomar con elindividuo acostado o sentado. (Fig. 2)En el individuo acostado se proce<strong>de</strong> alevantar ligeramente el miembro don<strong>de</strong> seva tomar el reflejo colocando una mano anivel <strong>de</strong>l hueco poplíteo, <strong>de</strong> manera que lapierna que<strong>de</strong> ligeramente flexionada.Conseguida dicha posición se percute conel martillo <strong>de</strong> reflejos sobre el tendónrotuliano.En el individuo acostado: se coloca el pie<strong>de</strong>l miembro a investigar sobre el maléolo<strong>de</strong> la pierna opuesta; se toma la planta <strong>de</strong>lpie don<strong>de</strong> se va a investigar el reflejoflexionándolo ligeramente y se percute eltendón <strong>de</strong> Aquiles.También pue<strong>de</strong> tomarse con el individuoarrodillado sobre una silla o sobre lacama.Se reflexiona ligeramente el pié don<strong>de</strong> seva a investigar el reflejo y se percute eltendón.Reflejo bulbocavernoso: la excitación <strong>de</strong> lamucosa <strong>de</strong> glan<strong>de</strong> da como respuesta lacontracción <strong>de</strong>l músculo bulbocavernoso.Su centro se halla en el S3 y S4.REFLEJOS PUPILARES:Reflejo fotomotor: Se investiga usando laluz natural o por medio <strong>de</strong> una linterna.Fig. 2: Reflejo patelar: componentes<strong>de</strong>l arco reflejoSu respuesta es la extensión <strong>de</strong> la pierna y su centrolos segmentos L2; L3 y L4.En enfermos sentados: se solicita que lapersona a investigar <strong>de</strong>je sus piernascolgando <strong>de</strong>l bor<strong>de</strong> <strong>de</strong> la silla y se proce<strong>de</strong>a golpear el tendón rotuliano con elmartillo <strong>de</strong> reflejos. La repuesta es igual ala <strong>de</strong>l individuo acostado.Reflejo plantar: Se investiga pasando layema <strong>de</strong>l <strong>de</strong>do; la punta <strong>de</strong> un alfiler o unlápiz sobre la planta <strong>de</strong>l pié.Produce como respuesta la flexión <strong>de</strong> los <strong>de</strong>dos y sucentro se halla en S1 y S2.Reflejo aquiliano: Se pue<strong>de</strong> tomar con elindividuo sentado o acostado.En el individuo sentado se solicita que se coloque<strong>de</strong>jando colgar sus piernas al bor<strong>de</strong> <strong>de</strong> la silla y sepercute el tendón <strong>de</strong> Aquiles con el martillo <strong>de</strong> reflejos.La repuesta es la extensión <strong>de</strong>l pié y sucentro son L5 y S1 - S2.Se solicita a la persona en quien se va a investigar elreflejo que mire a lo lejos.El investigador <strong>de</strong>l reflejo <strong>de</strong>be tapar consus manos los ojos <strong>de</strong>l individuo ainvestigar.Luego retira la mano que cubre uno <strong>de</strong> losojos, y a continuación tapa éste y retira laotra mano (<strong>de</strong>jando siempre cubierto el ojoque no se está investigando).Bajo la acción <strong>de</strong> la luz la pupila secontrae. Su centro se halla en lostubérculos cuadrigéminos anteriores.Reflejo <strong>de</strong> acomodación: Se pi<strong>de</strong> alindividuo a quien se va investigar el reflejoque mire a lo lejos y luego que dirija sumirada al <strong>de</strong>do índice <strong>de</strong>l investigador que<strong>de</strong>be colocarse a unos 30 cm <strong>de</strong>distancia <strong>de</strong>l ojo: la respuesta es unacontracción <strong>de</strong> la pupila.Este reflejo investiga la vía: retina - nervio óptico;corteza occipital y vías <strong>de</strong> asociación con los núcleos<strong>de</strong>l nervio motor ocular común.Los reflejos <strong>de</strong>tallados en esta guía correspon<strong>de</strong>n alos estudiados en exploraciones clínicas.
CATEDRA Nº 1 DE FISIOLOGIA 2006 31Ellos tienen mucho valor como exploraciónfuncional <strong>de</strong>l sistema nervioso por el hecho<strong>de</strong> que tienen vías muy precisas y<strong>de</strong>terminan localizaciones exactas <strong>de</strong>lesiones o patologías, pero <strong>de</strong> ningunamanera cubren el amplísimo espectro <strong>de</strong>activida<strong>de</strong>s reflejadas que permiten unaa<strong>de</strong>cuada función <strong>de</strong>l organismo humano.d) Con una extensión muscular fuerteexiste un incremento en la actividad<strong>de</strong> las fibras nerviosas aferentes <strong>de</strong>ltendón (Golgi).Temario a <strong>de</strong>sarrollar durante laautoinstrucción1. Médula espinal como centro reflejo.2. Arco reflejo simple.3. Reflejos mono y multisegmentarios.4. Reflejos superficiales y profundos.PREGUNTAS DE AUTOEVALUACION1. Indique cuales <strong>de</strong> los reflejos arribamencionados son superficiales ycuales son profundos.2. Señale si entre los reflejos arribamencionados hay reflejos miotáticos,miotáticos inversos y <strong>de</strong> <strong>de</strong>fensa. Encaso afirmativo indique cuales reflejoscorrespon<strong>de</strong>n a cada uno <strong>de</strong> los tipos<strong>de</strong> reflejos mencionados.3. Mencione por lo menos 5 reflejosvisceroceptivos.4. Cuando un músculo esqueléticonormalmente inervado es extendido, larepuesta inicial es contracción, perocon una extensión creciente el músculorepentinamente se relaja.Esta relajación se produce porque:a) Con una extensión fuerte se producela <strong>de</strong>scarga eferente gamma.b) Con una extensión fuerte se inhibe la<strong>de</strong>scarga <strong>de</strong> las terminacionesespirales <strong>de</strong> las fibras nerviosasaferentes.c) Con una extensión muscular fuerteexiste reducción <strong>de</strong> la actividad <strong>de</strong> lasfibras nerviosas aferentes <strong>de</strong> losórganos tendinosos <strong>de</strong> Golgi.
CATEDRA Nº 1 DE FISIOLOGIA 2006 32Fig. 3: Distintos reflejos y porción <strong>de</strong>l sistema nervioso involucrada
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 33FISIOLOGIA DEL CEREBELO(Dra. Lilian Barrios y Dr. OscarPoletti)El cerebelo es consi<strong>de</strong>rado tradicionalmentecomo un sistema regulador motor <strong>de</strong>bido aque la estimulación eléctrica <strong>de</strong> suestructura no provoca sensaciones y solorara vez, algún movimiento.Sin embargo, la extirpación <strong>de</strong> estaestructura nerviosa produce una profundaalteración <strong>de</strong> los movimientos.y mantenimiento <strong>de</strong>l movimiento sonregulados por el neocerebelo, zonafilogenéticamente nueva que funciona enestrecha relación con la cortezacerebral.MORFOLOGIA DEL CEREBELOEste órgano presenta numerosascircunvoluciones transversales en susuperficie y cisuras que lo divi<strong>de</strong>n enlóbulos y lobulillos (Fig. Nº 1).El cerebelo está situado por encima <strong>de</strong>l tallocerebral, en la fosa cerebral posterior yrecibe sus aferencias y envía sus eferenciaspor medio <strong>de</strong> 3 pares <strong>de</strong> haces: lospedúnculos cerebelosos superior, medio einferior.Sus aferencias sensoriales, sobre todo <strong>de</strong>las que informan <strong>de</strong> la posición <strong>de</strong>l cuerpo,el largo muscular y su tensión, se integran anivel <strong>de</strong> la corteza cerebelosa y laseferencias restantes se transmiten a las víasmotoras <strong>de</strong>scen<strong>de</strong>ntes.Las aferencias y eferencias establecen trestipos <strong>de</strong> interrelaciones, hallándose estasvinculadas entre sí y en última instancia,controlan los músculos esqueléticos, por loque no pue<strong>de</strong>n in<strong>de</strong>pendizarse una <strong>de</strong> otra.1. La posición <strong>de</strong>l cuerpo: la posición<strong>de</strong>l cuerpo en el espacio es informadapor el tracto vestíbulocerebeloso alarchicerebelo. Las eferencias quevuelven a los núcleos vestibulares yotros núcleos <strong>de</strong>l tallo cerebralcontribuyen a reflejos vestibulares queregulan músculos <strong>de</strong>l ojo, cuello, troncoy extremida<strong>de</strong>s comprometidos en lafunción <strong>de</strong>l equilibrio.2. La tensión: la información <strong>de</strong> tensiónen los músculos esqueléticos,articulaciones y tendones, junto a lainformación exteroceptiva, se dirigen alpaleocerebelo por las víasespinocerebelosas y las víascuneocerebelosas. Esta porción <strong>de</strong>lcerebelo interviene en la regulación <strong>de</strong>ltono muscular.Fig. Nº 1: morfología cerebelosaSu cara posteroinferior se aloja en la fosacraneal posterior, su cara superior está encontacto con la tienda <strong>de</strong>l cerebelo y caraanterior contacta con la cara posterior <strong>de</strong>lfallo cerebral, al cual está conectado por lospedúnculos cerebelosos superior, medio einferior.Presenta una estructura central: el vermis ydos hemisferios laterales.Las regiones cerebelosas han sidoclasificadas <strong>de</strong> diversas formas:Una clasificación filogénica, fisiológica yclínica divi<strong>de</strong> al cerebelo en:ARCHICEREBELOPALEOCEREBELONEOCEREBELO (Fig. Nº 2)3. Los patrones <strong>de</strong> movimientosvoluntarios y automáticos y el ajuste <strong>de</strong>ltono que acompañan al inicio, <strong>de</strong>sarrollo
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 34Estos autores divi<strong>de</strong>n al cerebelo en unaparte media, en relación con los Núcleos <strong>de</strong>ltecho (Fastigii); una zona intermedia que serelaciona con los Núcleos Globoso yEmboliforme (Núcleos Intermedios oInterpósitos) y una zona lateral que seproyecta hacia el Núcleo <strong>de</strong>ntado. (Fig. Nº3)Fig. Nº 2: división funcional <strong>de</strong>l cerebelo.Una clasificación anatomofisiológicatransversal ha sido hecha por Larsell, quiendivi<strong>de</strong> al cerebelo en lóbulos superpuestos<strong>de</strong> a<strong>de</strong>lante a atrás, separados entre sí porsurcos transversales que se presentan tantoen los lóbulos laterales como en el vermis:LOBULO FLOCULONODULAR: que compren<strong>de</strong> elnódulo en el plano medio y el flóculo a loslados.LOBULO ANTERIOR: constituido por la casitotalidad <strong>de</strong> la cara anterior <strong>de</strong>l cerebelo eintegrado por la língula, el lobulillo central yel culmen en el vermis y por los lobulillosanterior y amígdala en los hemisferioslaterales.LOBULO POSTERIOR: constituido por el lóbuloansiforme y el lobulillo digástrico.Una clasificación anatómica <strong>de</strong> Brodal yJansen, longitudinal.•La clasificación <strong>de</strong> Brodal y Jansen se basaen las conexiones entre la cortezacerebelosa y sus núcleos profundos.Fig. Nº 3: Núcleos cerebelososDenominan a las zonas como:VERMIANA O MEDIA: que se conecta con elnúcleo Fastigii.LATERAL o HEMISFERICA: que se conectacon los núcleos <strong>de</strong>ntados.INTERMEDIA: que se conecta con los núcleosgloboso y emboliforme.A esta segmentación se agrega una parte<strong>de</strong>l lóbulo flóculo – nodular que en suporción nodular se conecta con los núcleosvestibulares (equivalentes a los núcleoscerebelosos).Un aspecto interesante <strong>de</strong> la clasificación <strong>de</strong>Brodal y Jansen se <strong>de</strong>riva <strong>de</strong>l hecho <strong>de</strong>señalar las vías motoras (vías piramidal yrubroespinal) con que se conectan lasdiversas regiones cerebelosas.La zona <strong>de</strong>l vermis, por medio <strong>de</strong> axonesque nacen en el núcleo Fastigii, se conectacon la formación reticular y <strong>de</strong>scien<strong>de</strong> por elhaz reticuloespinal hacia las motoneuronas(recor<strong>de</strong>mos que estas vías reguladoras
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 35terminan en interneuronas propioespinalescortas).Una porción <strong>de</strong> axones <strong>de</strong> las células <strong>de</strong>Purkinje cerebelosas <strong>de</strong>l vermis no pasanpor el núcleo Fastigii, sino que hacenestación directamente en los núcleosvestibulares (involucrados en la regulación<strong>de</strong>l equilibrio), que por el hazvestibuloespinal también terminan eninterneuronas reguladoras <strong>de</strong> las αmotoneuronas.La zona intermedia, por medio <strong>de</strong> losnúcleos intermedios o interpósitos, seconectan con el núcleo rojo en la porción <strong>de</strong>origen <strong>de</strong>l haz lateral o rubroespinal.La zona lateral, previa estación en el núcleo<strong>de</strong>ntado se proyecta al núcleo ventrolateral<strong>de</strong>l tálamo cuyos axones terminan en el áreaIV motora, inicio <strong>de</strong> la vía piramidal. Unacantidad menor <strong>de</strong> axones termina en laporción parvocelular <strong>de</strong>l núcleo rojo.Estas tres clasificaciones para lasegmentación <strong>de</strong>l cerebelo son importantespues cada una aborda la organización <strong>de</strong>lmismo <strong>de</strong>s<strong>de</strong> una perspectiva diferente.ORGANIZACIÓN DE LA CORTEZACEREBELOSAA diferencia <strong>de</strong> la corteza cerebral quepresenta distintas capas <strong>de</strong> células según lazona estudiada, el cerebelo es uniforme entoda su extensión significando estacircunstancia que, probablemente, elprocesamiento y respuesta que da es similaren toda su superficie.Esto significa que el archi; palio yneocerebelo tienen exactamente las mismascapas celulares.Para recibir sus aferencias, procesar lainformación y enviar su respuesta elcerebelo consta <strong>de</strong>:Fibras aferentesCorteza cerebelosaNúcleos profundos cerebelososLa corteza cerebelosa humana es en erealidad una gran masa plegada, con lospliegues colocados en forma transversal.Cada doblez se llama folio. En laprofundidad <strong>de</strong> la masa plegada seencuentran los núcleos profundoscerebelosos.FIBRAS AFERENTES: las aferencias a lacorteza cerebelosa entran por medio <strong>de</strong> dossistemas <strong>de</strong> fibras <strong>de</strong> tipo excitatorio:FIBRAS MUSGOSAS: <strong>de</strong> médula: <strong>de</strong> tronco cerebral: <strong>de</strong> neocortezaFIBRAS TREPADORAS: <strong>de</strong> las olivasbulbaresLas fibras musgosas <strong>de</strong> la médula provienen<strong>de</strong> los husos musculares y <strong>de</strong> los órganostendinosos <strong>de</strong> Golgi (víasespinocerebelosas), <strong>de</strong> los receptoresarticulares y en menor medida <strong>de</strong> lasensibilidad profunda consciente.Las <strong>de</strong>l tallo cerebral provienen <strong>de</strong> losnúcleos <strong>de</strong> la sustancia reticular y <strong>de</strong> losnúcleos vestibulares.Las <strong>de</strong> neocorteza proce<strong>de</strong>n <strong>de</strong> los lóbulosfronto-parieto-témporo-occipitales.Toda la información que entra al cerebelo(excepto la que va al nódulo flóculonodular)se organiza somatotópicamente.Antes <strong>de</strong> <strong>de</strong>scribir las conexiones <strong>de</strong> lasfibras aferentes cerebelosas<strong>de</strong>sarrollaremos la organización celular <strong>de</strong> lacorteza cerebelosa.CORTEZA CEREBELOSA: consta <strong>de</strong> trescapas histológicas (Fig. Nº 4).Fig. Nº 4: organización histológica <strong>de</strong>lcerebelo.La capa más profunda se <strong>de</strong>nomina capagranular y contiene células grano en alta<strong>de</strong>nsidad. Las células grano son lasneuronas que reciben la mayor parte <strong>de</strong> lasaferencias a la corteza.
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 36En esta etapa también se ven células <strong>de</strong>Golgi, interneuronas cerebelosas.La capa media presenta células <strong>de</strong> Purkinje.Esta capa presenta una sola hilera <strong>de</strong> dichascélulas, que constituyen las eferenciascerebelosas.La capa más superficial es la molecular; enella se encuentran células en canasto ycélulas estrelladas. A<strong>de</strong>más, la capamolecular contiene las <strong>de</strong>ndritas <strong>de</strong> lascélulas <strong>de</strong> Golgi y <strong>de</strong> las células <strong>de</strong>Purkinje. Las fibras presentes en esta capason los axones <strong>de</strong> las células en grano.Estos axones ascien<strong>de</strong>n <strong>de</strong>s<strong>de</strong> la capagranulosa y al llegar a la capa molecular sebifurcan en ramas paralelas a lasbifurcaciones cerebelosas (sentidotransversal), dando un sistema <strong>de</strong> fibrasparalelas.Las fibras musgosas hacen sinapsis con las<strong>de</strong>ndritas <strong>de</strong> las células en grano. Se hacalculado que una sola fibra musgosa pue<strong>de</strong>hacer sinapsis con unas 3.000 células engrano y que, varias fibras trepadoras pue<strong>de</strong>nterminar en <strong>de</strong>ndritas <strong>de</strong> una misma célulagranular.Las células granulares emiten axones quevan a la superficie <strong>de</strong> la corteza cerebelosay se divi<strong>de</strong>n en dos ramas opuestasparalelas: en el curso <strong>de</strong> su trayecto hacensinapsis con las arborizaciones <strong>de</strong>ndríticas<strong>de</strong> las células <strong>de</strong> Purkinje.El número <strong>de</strong> células granulosas se estimaen varios miles; y en unas 200.000 elnúmero <strong>de</strong> fibras paralelas que atraviesanlas <strong>de</strong>ndritas <strong>de</strong> las células <strong>de</strong> Purkinje.Las fibras paralelas a su vez, emiten trestipos <strong>de</strong> colaterales que se conectan contres interneuronas inhibidoras:1. Las células <strong>de</strong> Golgi: que aplican suaxón sobre regiones pre y postsinápticas<strong>de</strong> la conexión entre las fibras musgosasy las células granulares.2. Las células estrelladas, que ejercensu acción sobre las <strong>de</strong>ndritas <strong>de</strong> lascélulas <strong>de</strong> Purkinje, inhibiéndola.3. Las células en cesto, cuyos axonesse aplican directamente sobre lascélulas <strong>de</strong> Purkinje a través <strong>de</strong> lassinapsis axosomáticas, inhibiéndolas.Estas células terminan en una <strong>de</strong>cena <strong>de</strong>células <strong>de</strong> Purkinje.El sistema <strong>de</strong> fibras musgosas poseeconvergencia y divergencia masivas. Cuandose activan las fibras paralelas <strong>de</strong> las célulasen grano, en las que estas fibras terminan,se excitan alre<strong>de</strong>dor <strong>de</strong> 50 células <strong>de</strong>Purkinje. Hacen conexión sináptica con lascélulas <strong>de</strong> Purkinje a través <strong>de</strong> las célulasen grano.Las fibras trepadoras poseen convergencia ydivergencia, su axón sube y se ramifica enfinas ramas alre<strong>de</strong>dor <strong>de</strong> las <strong>de</strong>ndritas <strong>de</strong>las células <strong>de</strong> Purkinje. Cada fibra trepadoratermina en una sola célula <strong>de</strong> Purkinje, enforma monosináptica.La disposición <strong>de</strong> neuronas e interneuronasen el cerebelo, <strong>de</strong>termina que las fibrasmusgosas produzcan una excitación seguidapor un período <strong>de</strong> inhibición. La excitaciónse <strong>de</strong>be a la acción <strong>de</strong> las células en granoy la inhibición a la conexión con las célulasestrelladas y al efecto <strong>de</strong> las células encanasto.Las fibras trepadoras tienen efectoexcitatorio sobre las células <strong>de</strong> Purkinje.Las células <strong>de</strong> Purkinje tienen efectoinhibitorio y su neurotransmisor es el GABA.Sus axones hacen sinapsis en losrespectivos núcleos profundos cerebelosos ysobre el núcleo <strong>de</strong> Deiters. Este efectoinhibidor modula el efecto excitatorio <strong>de</strong> lasneuronas <strong>de</strong> dichos núcleos, según lasnecesida<strong>de</strong>s <strong>de</strong> regulación <strong>de</strong> la actividadmotora.Hay colaterales <strong>de</strong> las células <strong>de</strong> Purkinjesobre las neuronas internuncialesinhibidoras <strong>de</strong> las fibras musgosas, lo cualconstituiría un circuito <strong>de</strong> <strong>de</strong>sinhibición.Por lo tanto, las células <strong>de</strong> Purkinje sonasiento <strong>de</strong> procesos sucesivos o simultáneos<strong>de</strong> excitación e inhibición.ARCHICEREBELOEsta porción <strong>de</strong>l cerebelo está involucradaen la regulación <strong>de</strong>l equilibrio. Susaferencias provienen <strong>de</strong>l aparato vestibularque consta <strong>de</strong> un sistema <strong>de</strong> controles queactúa <strong>de</strong> manera refleja para estabilizar lavisión y coordinar los movimientos <strong>de</strong>lcuerpo, la cabeza y los miembros.
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 37El aparato vestibular incluye: (Fig. 5)Fisiología <strong>de</strong> los conductos semicirculares:(Fig. Nº 6).Fig. 5: aparato vestibular√ Tres canales semicirculares provistoscada uno, <strong>de</strong> una ampolla que contieneuna placa pequeña llamada cresta. Estoscanales llenos <strong>de</strong> endolinfa, estándispuestos entre sí en ángulo recto yrepresentan los tres planos <strong>de</strong>l espacio.Estas estructuras <strong>de</strong>tectan y mi<strong>de</strong>n lasaceleraciones angulares en cualquierdirección.√ El utrículo y el sáculo. En estasestructuras hay una placa <strong>de</strong>neuroepitelio con células pilosasorientadas sistemáticamente para quesean sensibles a los <strong>de</strong>splazamientos enaceleraciones lineales y en ciertamedida, vibraciones. Dichas célulasconstituyen el receptor sensorial llamadomácula.Fig. Nº 6: órgano sensorial <strong>de</strong> losconductos semicirculares.Cada ampolla contiene una cresta conepitelio sensorial cubierto con una cúpulagelatinosa, en n la cual penetran lasestereocilias <strong>de</strong> las células pilosas.Cada célula ciliada tiene un promedio <strong>de</strong> 50a 100 pequeños cilios <strong>de</strong>nominadosestereocilios y un cilio muy gran<strong>de</strong> llamadocinocilio que se encuentra siempre <strong>de</strong>lmismo lado respecto <strong>de</strong> la orientación <strong>de</strong> lascélulas sobre la cresta.Cuando hay <strong>de</strong>splazamiento <strong>de</strong> losestereocilios <strong>de</strong> unos 100 pm, la célularespon<strong>de</strong> con una <strong>de</strong>spolarización <strong>de</strong> sufibra sensorial.Los estereocilios tienen tamaño graduado encuanto a su largo, con respecto a lo máscercano al cinocilio.La inclinación hacia un lado <strong>de</strong>l estereocilioproduce su <strong>de</strong>spolarización, y hacia el ladoopuesto, hiperpolarización.Como el conducto está lleno <strong>de</strong> endolinfa, almoverse la cabeza se mueve la endolinfa yesta moviliza a los cilios.
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 38Fisiología <strong>de</strong>l utrículo y <strong>de</strong>l sáculo: (Fig. Nº7).Los núcleos vestibulares dan ramas<strong>de</strong>scen<strong>de</strong>ntes que van a la rama espinal.También envían colaterales a:√ Archicerebelo√ Núcleos motores III, IV y VI√ Tubérculo cuadrigémino superior√ Corteza cerebralPor medio <strong>de</strong> estas fibras producen elcontrol reflejo <strong>de</strong> los movimientos ocularesen relación con el movimiento <strong>de</strong> la cabeza;y el control <strong>de</strong> los movimientos <strong>de</strong> la cabezay miembros en movimiento, permitiendomantener la situación <strong>de</strong> equilibrio en lapersona.PALEOCEREBELOFig. Nº 7.utrículo y sáculo.Mientras los conductos semicircularesactúan en virtud <strong>de</strong> las propieda<strong>de</strong>sinerciales <strong>de</strong> la endolinfa, la mácula actúapor la interacción <strong>de</strong> sus células epitelialesciliadas con los otolitos y la capa gelatinosa<strong>de</strong> soporte don<strong>de</strong> están los cilios. (Fig. Nº8)Correspon<strong>de</strong> a un <strong>de</strong>sarrollo filogenéticomás tardío que el <strong>de</strong>l archicerebelo.Compren<strong>de</strong> el lóbulo anterior y parteposterior <strong>de</strong>l vermis. Recibe como aferentesimportantes a las vías espinocerebelosasque dan información <strong>de</strong> los segmentoslumbar y dorsal y las vías espinocerebelosasrostral y cúneocerebelosas que traeninformación propioceptiva cervical.También llegan al paleocerebelo informacióncutánea, visual y auditiva.Esta zona cerebelosa presenta somatotopía<strong>de</strong> todo el cuerpo con la zona medial <strong>de</strong>lmismo representada a nivel <strong>de</strong>l vermis y la<strong>de</strong> los miembros a nivel <strong>de</strong> los hemisferioslaterales.Las vías eferentes <strong>de</strong>l paleocerebelo<strong>de</strong>scien<strong>de</strong>n por el haz rubroespinal.Fig. Nº: 8: órgano sensorial <strong>de</strong>lutrículo y sáculoLos movimientos <strong>de</strong> aceleración lineal,mueven más fácilmente a la capa gelatinosaque a los otolitos, y esto <strong>de</strong>forma los cilios y<strong>de</strong>scarga la terminal sensitiva.La vía aferente <strong>de</strong>l aparato vestibular tienesu neurona en el ganglio vestibular.Los axones <strong>de</strong> las neuronas <strong>de</strong>l gangliovestibular terminan en parte en los núcleosbulboprotuberanciales, y en parte (sin hacersinapsis) en el archicerebelo.Esta porción <strong>de</strong>l cerebelo interviene en laregulación central <strong>de</strong>l tono muscular y por lotanto en la regulación <strong>de</strong> la postura.NEOCEREBELOEs la adquisición filogenéticamente másnueva <strong>de</strong>l órgano.Sus fibras aferentes provienen <strong>de</strong> losdistintos lóbulos cerebrales, que se dirigena hacer sinapsis con los núcleos <strong>de</strong>l puente,formando las fibras fronto-ponto-parietopónticasque ingresan al neocerebelo por lospedúnculos cerebelosos superiores. Laseferencias cerebelosas se dirigen al núcleoventrolateral <strong>de</strong>l tálamo y se proyectanluego a la zona motora cortical.
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 39Se consi<strong>de</strong>ra que estos circuitos ejercen suacción reguladora motora sobre la zona <strong>de</strong>comando <strong>de</strong> las motoneuronas.Todas las lesiones cerebelosa traen comoconsecuencia la producción <strong>de</strong> movimientosmotores anormales. En general las lesiones<strong>de</strong>ben ser <strong>de</strong> magnitud para que dichasanormalida<strong>de</strong>s aparezcan.Si los núcleos cerebelosos profundos estánin<strong>de</strong>mnes, pareciera que hay capacidad <strong>de</strong>compensación por parte <strong>de</strong> las estructurascerebelosas in<strong>de</strong>mnes, lo que permitemantener las funciones motorasprácticamente normales.Dentro <strong>de</strong> los síntomas <strong>de</strong> lesión cerebelosase pue<strong>de</strong>n mencionar:– ATAXIA: caracterizada por la incoordinación en lafrecuencia, fuerza y dirección <strong>de</strong> los movimientos.Da la “marcha <strong>de</strong>l ebrio” <strong>de</strong>l cerebeloso.– DISMETRIA: caracterizada por movimientos quesuelen ir más allá <strong>de</strong>l fin propuesto.– TEMBLOR INTENCIONAL: temblor producido porla contracción en sacudidas <strong>de</strong> los músculoscuando se va a realizar un movimiento.– DISARTRIA: producida por los movimientos conprogresividad irregular <strong>de</strong> los músculosinvolucrados en la formación <strong>de</strong> las palabras.Recor<strong>de</strong>mos que la habilidad <strong>de</strong> hablar exige unafunción muscular muy perfeccionada.– ADIADOCOCINESIA: producida por la incapacidad<strong>de</strong> estos enfermos <strong>de</strong> ejecutar movimientosopuestos alternantes en forma rápida (por ejemplo:pronación y supinación <strong>de</strong> un miembro).c) e Purkinjed) <strong>de</strong> Golgi2: El archicerebelo con su núcleoFastigii regulan:a) La precisión y elasticidad <strong>de</strong> losmovimientosb) El tono muscularc) El equilibrio corporald) La predicción <strong>de</strong> los movimientos3: Las fibras eferentes <strong>de</strong>l cerebeloson <strong>de</strong> carácter EXCITATORIO –INHIBITORIO y tiene su origen en lasneuronas <strong>de</strong> LOS NUCLEOSPROFUNDOS DEL CEREBELO – LACORTEZA CEREBELOSA (tache lo queno corresponda).4: Por lo general la estimulación <strong>de</strong>las áreas <strong>de</strong>l cerebelo SI – NO producemovimientos <strong>de</strong>l músculo esquelético;SI – NO produce movimiento <strong>de</strong>músculo visceral y SI – NO produceexperiencias sensitivas (tache lo queno corresponda).PREGUNTAS A CONTESTAR DURANTE LAAUTOINSTRUCCION:a) Organización <strong>de</strong>l cerebelo.b) Función <strong>de</strong>l archicerebelo.c) Función <strong>de</strong>l paleocerebelo.d) Función <strong>de</strong>l neocerebelo.– HIPOTONIA: pérdida <strong>de</strong>l tono motor <strong>de</strong> lamusculatura periférica en el lado <strong>de</strong> la lesión.– NISTAGMO CEREBELOSO: producido por eltemblor <strong>de</strong> los globos oculares, se presenta cuandose quiere fijar los ojos en un objeto situado a unlado <strong>de</strong>l individuo.PREGUNTAS DE AUTOEVALUACION1: La única vía que sale <strong>de</strong> la cortezacerebelosa hacia los núcleos es laformada por los axones <strong>de</strong> las células:a) Estrelladasb) Granulares
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 40REGULACION DE LA POSTURA( Dra. Lilian Barrios)IntroducciónEl análisis <strong>de</strong> la fisiología <strong>de</strong>l sistemanervioso se hace más comprensible cuandola zona en estudio es asociada con lasperiferias sensitiva o motora. El sistemanervioso consta <strong>de</strong> sistemas sensorialesque constantemente nos informan acerca <strong>de</strong>nuestra situación física en el medio que noscircunda.Por otro lado, nuestros sistemas motoresson los agentes que nos permiteinteraccionar con el mismo puesto, que nospermite movernos, hablar, escribir, etc. y <strong>de</strong>esa manera alterar nuestra relación físicacon el medio que nos ro<strong>de</strong>a, buscandonuestra a<strong>de</strong>cuación al mismo.Los músculos <strong>de</strong>l organismo son losencargados <strong>de</strong> realizar esta tarea. Laprecisión <strong>de</strong>l movimiento <strong>de</strong> cada músculoesquelético es controlado separadamentepor un grupo <strong>de</strong> neuronas <strong>de</strong>l sistemanerviosos central.Estas neuronas se <strong>de</strong>nominan neuronasesqueletomotoras y en cada masa muscular,las neuronas esqueletomotoras relacionadascon ella, controlan solo a una parte <strong>de</strong> lacantidad <strong>de</strong> fibras musculares que la forman.Cada neurona esqueletomotora y las fibrasque inerva constituyen la estructura<strong>de</strong>nominada unidad motora.Las distintas unida<strong>de</strong>s motoras constituyenel <strong>de</strong>stino final común <strong>de</strong> las señales <strong>de</strong>comando <strong>de</strong>l área motora <strong>de</strong>l sistemanervioso central.La armonía en un grupo muscular funcionalsituado alre<strong>de</strong>dor <strong>de</strong> una articulación pue<strong>de</strong>verse en el simple acto <strong>de</strong> elevar la piernapara alejar el píe <strong>de</strong> un estímulo doloroso(por ej. al pisar un objeto punzante queproduce dolor).Tal acción requiere la flexión <strong>de</strong> larodilla y es realizada cuando el bícepsfemoral y su sinergista el semitendinoso,se contraen; al mismo tiempo, su músculoextensor antagonista, el cuadrícepsfemoral, se relaja. Los grados <strong>de</strong>contracción y relajación se hallan <strong>de</strong> talmanera sincronizados y coordinados porlas neuronas esqueletomotoras que actúansobre los diferentes músculos, que setiene por resultado una flexión graduada.Esta armonía funcional no se <strong>de</strong>be a laactuación <strong>de</strong> una sola articulación, lamisma opera mediante variasarticulaciones. Al pisar el objetopunzante que produjo dolor, la flexión <strong>de</strong>la rodilla se sincroniza con ladorsiflexión <strong>de</strong>l pie y la flexión <strong>de</strong> laca<strong>de</strong>ra.La sincronización <strong>de</strong> la contracción y larelajación entre las unida<strong>de</strong>s motoras <strong>de</strong>estos músculos flexores y extensores esproducida por la inervación recíproca <strong>de</strong> losgrupos musculares; es <strong>de</strong>cir la inhibición <strong>de</strong>los músculos antagonistas y la facilitación<strong>de</strong> los músculos sinergistas produce unaacción recíproca <strong>de</strong> fina gradación en losmúsculos involucrados en este movimiento.Por otro lado, diferentes receptoressensoriales periféricos informanconstantemente al SNC acerca <strong>de</strong> laposición, la velocidad, el tacto, la presión ylos estados <strong>de</strong> frío y calor <strong>de</strong> unaextremidad. Esta información sensorial quellega momento a momento al SNC es usadapara dirigir la motilidad, lo que permite a lossistemas motores producir una gama notable<strong>de</strong> movimientos. La integraciónsensoriomotora tiene lugar a diferentesniveles <strong>de</strong> los sistemas motores y cada nivelcontribuye a su manera a la fina regulación<strong>de</strong> los movimientos musculares.Los movimientos varían mucho en cuanto asu complejidad. Algunos movimientos sonsimples y no requieren una <strong>de</strong>cisiónconsciente: por ejemplo el reflejo miotático.Otros movimientos, como la escritura, sonmás complejos y requieren un constantecomponente consciente. Lo que diferencia aun movimiento simple <strong>de</strong> uno mas complejoes el grado <strong>de</strong> integración sensoriomotora,esto es; el grado en que la informaciónsensorial cambiante modifica una<strong>de</strong>terminada conducta motora programada.(Ejemplo: una persona que está escribiendouna carta, si siente olor a quemado,interrumpirá inmediatamente ese acto motor,para averiguar a que se <strong>de</strong>be el olor quepercibe)Los músculos esqueléticos estáninmóviles, a menos que se or<strong>de</strong>ne unacontracción. El tono <strong>de</strong> estos músculos ylos reflejos posturales esqueléticos
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 41dinámicos, tónicos y fásicos, incluyendoreflejos entre extremida<strong>de</strong>s y <strong>de</strong> lacabeza sobre el cuerpo, básicos paracaminar, correr y nadar, son generados ymantenidos por sistemas <strong>de</strong> controlsensitivomotores <strong>de</strong>l tallo cerebral yespinales longitudinales. De esta manera,los centros nerviosos superiores seahorran una preocupación excesiva encuanto a los mecanismos <strong>de</strong> control <strong>de</strong>lnivel inferior, que subyacen alrendimiento coordinado <strong>de</strong> los músculosesqueléticos.Los controles <strong>de</strong> niveles más bajos son losque proporcionan sostén reflejo <strong>de</strong>l cuerpocontra la fuerza <strong>de</strong> gravedad y el estado <strong>de</strong>alerta muscular para la acción.Muchas funciones espinales importantes eintegradoras <strong>de</strong>l tallo cerebral, estánorganizadas en forma relativamenteautónoma en forma segmentaria ( horizontal)y también a lo largo <strong>de</strong>l neuroeje(longitudinalmente), con el tallo cerebral y lamédula espinal organizados funcionalmente,para un control integrado entre lasextremida<strong>de</strong>s y la cabeza sobre el cuerpo.REGULACION DE LA POSTURALa postura se <strong>de</strong>fine como laresistencia muscular activa al<strong>de</strong>splazamiento corporal por lagravedad o la fuerza aceleratoria.El mantenimiento <strong>de</strong> una posturaerecta es el sustrato esencial parapo<strong>de</strong>r realizar movimientos <strong>de</strong> tipofásico con un fin <strong>de</strong>terminado.En el mantenimiento <strong>de</strong> la posturaerecta juegan un papel esenciallos ajustes reflejos en los músculosextensores mediales, comorespuesta a las fuerzas quetien<strong>de</strong>n al <strong>de</strong>splazamientocorporal. Debido a ello, losmúsculos extensores medialesreciben el nombre <strong>de</strong> músculosantigravitatorios.Si bien fisiológicamente casi todos losmúsculos pue<strong>de</strong>n ser clasificados comoflexores o extensores, (siendoextensor fisiológico aquel músculocuya acción se realiza en contra<strong>de</strong> la gravedad), una clasificaciónen medial (ó proximal) y lateral (ódistal) <strong>de</strong> las masas musculares,permite un mejor análisis <strong>de</strong>lcontrol central <strong>de</strong>l movimiento.La musculatura medial o proximal incluye,por ejemplo, los músculos <strong>de</strong>l eje vertebral y<strong>de</strong> las cinturas escapular y pelviana, ya seanextensores o flexores. Las acciones <strong>de</strong> estosmúsculos se ejercen sobre el eje corporal ylos músculos proximales. De esta maneracumplen funciones que requierenmovimientos completos <strong>de</strong> todo el cuerpo y<strong>de</strong> los miembros, tales como la postura, lamarcha y el equilibrioLa musculatura lateral o distal incluye losmúsculos intrínsecos <strong>de</strong> los <strong>de</strong>dos, y losmúsculos distales <strong>de</strong> las extremida<strong>de</strong>s.Tales músculos flexores o extensores, nocumplen función postural y en general sonmediadores <strong>de</strong> activida<strong>de</strong>s manipuladoras ó<strong>de</strong> tipo fásico.Reflejos posturalesHay dos gran<strong>de</strong>s clases <strong>de</strong> reflejosposturales:Los reflejos estatocinéticos que seoriginan al realizar <strong>de</strong>splazamientosaceleratorios <strong>de</strong>l cuerpo yLos reflejos estáticos, que sonoriginados por los <strong>de</strong>splazamientosantigravitatorios impuestos al cuerpoComo los reflejos estatocinéticos seoriginan en estímulos <strong>de</strong> tipo aceleratorio, larama aferente <strong>de</strong> tales circuitos reflejosimplica a los laberintosUn ejemplo <strong>de</strong> reflejo estatocinético causadopor aceleración lineal es la reacciónvestibular <strong>de</strong> ubicación.Este reflejo es investigado en animales(gato) con los ojos cerrados. Para laobtención <strong>de</strong>l reflejo se sostiene un animal<strong>de</strong> la pelvis, con la cabeza orientada haciaabajo y se lo <strong>de</strong>scien<strong>de</strong> bruscamente. Ese<strong>de</strong>scenso brusco estimula la máculautricular y la respuesta refleja consiste enuna extensión <strong>de</strong> los miembros anteriores.Esta respuesta refleja pue<strong>de</strong> consi<strong>de</strong>rarsecomo una reacción adaptativa que prepara alanimal para realizar un soporte apropiado
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 42con sus miembros sobre una superficie <strong>de</strong>contacto. La <strong>de</strong>strucción <strong>de</strong>l utrículo suprimela respuesta. Sin embargo, si el animal tienelos ojos abiertos, la respuesta refleja esiniciada mediante estímulos visuales y enese caso se <strong>de</strong>nomina reacción visual <strong>de</strong>ubicación.En realidad, en los seres humanos la pérdida<strong>de</strong> información laberíntica no generaproblemas <strong>de</strong> reflejos posturales siempreque haya estímulos visuales disponiblesLos reflejos estatocinéticos causados porla aceleración angular, como se vedurante la rotación, se originan enseñales provenientes <strong>de</strong> los canalessemicirculares.Cuando una persona rota la cabeza (porej. hacia la <strong>de</strong>recha) se produce un flujo<strong>de</strong> endolinfa <strong>de</strong> los canalessemicirculares horizontales hacia laizquierda, hasta que se sobrepase lainercia <strong>de</strong> dicho fluido. Este flujo haciala izquierda mueve las células pilosas <strong>de</strong>las crestas semicirculares produciendouna <strong>de</strong>scarga <strong>de</strong> la vía aferenteoriginando reflejos <strong>de</strong> importancia enojos, cuello y miembros superiores.Los reflejos oculares se <strong>de</strong>nominannistagmo vestibular y son mediados poreferencias <strong>de</strong>l núcleo vestibular medial a losmúsculos oculares externos.La respuesta refleja al rotar la cabezahacia la <strong>de</strong>recha consiste en unmovimiento conjugado lento <strong>de</strong> los ojoshacia la izquierda (en dirección al flujo<strong>de</strong> la endolinfa. La respuesta reflejapermite que el centro <strong>de</strong> la mirada semantenga <strong>de</strong> manera correcta.A medida que prosigue la rotación, ya no sepue<strong>de</strong> mantener el punto <strong>de</strong> fijación y losojos se mueven rápidamente hacia la<strong>de</strong>recha para establecer un nuevo punto <strong>de</strong>fijación.Esta <strong>de</strong>sviación conjugada lenta endirección al flujo <strong>de</strong> la endolinfa y larápida vuelta en la dirección opuestaocurre repetidamente hasta que la inercia<strong>de</strong> la endolinfa es sobrepasada y laendolinfa pue<strong>de</strong> seguir entonces ladirección <strong>de</strong> la rotación.La lenta <strong>de</strong>sviación hacia la izquierda se<strong>de</strong>nomina componente lento <strong>de</strong>lnistagmo y el rápido retorno a la<strong>de</strong>recha se <strong>de</strong>nomina componenterápido, si bien por convención, ladirección <strong>de</strong>l nistagmo se nombracon la dirección <strong>de</strong>l componenterápido.Cuando la endolinfa fluye en el sentido <strong>de</strong> larotación <strong>de</strong>l canal semicircular, el nistagmo<strong>de</strong>saparece. Pero si la cabeza <strong>de</strong>ja<strong>de</strong> rotar, hay un estímulo <strong>de</strong>naturaleza <strong>de</strong>saceleratorio en elque la <strong>de</strong>sviación lenta <strong>de</strong> los ojosva hacia la <strong>de</strong>recha y el retornorápido hacia la izquierda.En ciertas enfermeda<strong>de</strong>s <strong>de</strong>l tallocerebral pue<strong>de</strong> aparecer nistagmo enausencia <strong>de</strong> estímulo y constituyeentonces un signo patológico. .En cuanto a los miembros superiores, larotación hacia la <strong>de</strong>recha produce extensión<strong>de</strong>l brazo <strong>de</strong>recho y la relajación o flexión<strong>de</strong>l izquierdoLa otra clase principal <strong>de</strong> reaccionesposturales son los reflejos estáticos, queproducen ajustes a los <strong>de</strong>splazamientosimpuestos por la gravedadDependiendo <strong>de</strong>l sitio efecto <strong>de</strong> la acciónrefleja los reflejos estáticos pue<strong>de</strong>nclasificarse como locales,segmentarios o generalesReflejos locales: uno <strong>de</strong> los reflejos másimportantes que sirve para mantener lapostura es mediado por la médula espinal yoriginado por la activación <strong>de</strong> lasterminaciones primarias (Ia) <strong>de</strong>l husomuscular. Este reflejo es el reflejo miotáticoEl huso neuromuscular es una estructura <strong>de</strong>varios milímetros <strong>de</strong> largo y unos pocos cientos <strong>de</strong>micrones <strong>de</strong> diámetro. Consta <strong>de</strong> una vaina <strong>de</strong>tejido conectivo que envuelve <strong>de</strong> 2 a 12 fibrasmusculares intrafusales. Cada extremo <strong>de</strong>l huso estáadosado a fibras musculares extrafusales. Loshusos se distribuyen a lo largo <strong>de</strong>l músculo y su<strong>de</strong>nsidad parece que varía con el grado <strong>de</strong>requerimiento para un músculo dado. Los músculosintrínsecos <strong>de</strong> los <strong>de</strong>dos poseen una <strong>de</strong>nsidadconsi<strong>de</strong>rablemente gran<strong>de</strong> <strong>de</strong> husos por masamuscular en relación a los gran<strong>de</strong>s músculoscomprometidos en el control postural.Existen por lo menos dos tipos <strong>de</strong> fibrasintrafusales: las fibras <strong>de</strong> bolsa
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 43nuclear y las fibras <strong>de</strong>nuclear.ca<strong>de</strong>naLas fibras <strong>de</strong> bolsa nuclear , que son más largas ymás anchas que las fibras <strong>de</strong> ca<strong>de</strong>na nuclear, poseenuna región expandida o ecuatorial con núcleos<strong>de</strong>nsamente agregados. Más allá <strong>de</strong> la zonaecuatorial se hallan elementos contráctiles.Las fibras <strong>de</strong> ca<strong>de</strong>na nuclear poseen unnúmero menor <strong>de</strong> núcleos dispuestos enserie en el centro <strong>de</strong> la fibra. Estas fibrastambién tienen elementos contráctiles en suspolos.El huso neuromuscular es un receptor <strong>de</strong>estiramiento (mecanorreceptor) quetransmite información sobre el largomuscular y la velocidad <strong>de</strong> cambio <strong>de</strong>este largo en función <strong>de</strong>l tiempo. Lainformación la transmiten a través <strong>de</strong>fibras nerviosas sensoriales aferentes .Fig. 1: huso muscularA nivel <strong>de</strong>l huso muscular nacen dos tipos <strong>de</strong>aferencias sensoriales: las terminacionesprimarias ( ó anuloespirales) que se enrollanalre<strong>de</strong>dor <strong>de</strong> las regiones centrales <strong>de</strong> lasfibras <strong>de</strong> bolsa nuclear. Estas fibras songran<strong>de</strong>s (12 a 20 µm), mielínicas, yconducen a 70 a 120 m/seg. Pertenecen algrupo <strong>de</strong> fibras <strong>de</strong> tipo Ia e informanvelocidad <strong>de</strong> estiramiento <strong>de</strong> lafibra muscular (receptoresdinámicos).Otras aferencias <strong>de</strong> tipo propioceptivo seoriginan en las terminaciones secundariasque ro<strong>de</strong>an a la región pericentral <strong>de</strong> lasfibras <strong>de</strong> ca<strong>de</strong>na nuclear <strong>de</strong>l huso.Estas aferencias también son receptores <strong>de</strong>estiramiento pero dan fundamentalmenteinformación sobre la longitudmuscular. Son más pequeñas, poseenmielina, su diámetro es <strong>de</strong> 5 a 12 µm conuna velocidad <strong>de</strong> conducción <strong>de</strong> 30 a 70m/seg. perteneciendo al grupo <strong>de</strong> fibras <strong>de</strong>tipo II. Un huso típico contiene dos fibras<strong>de</strong> bolsa nuclear y alre<strong>de</strong>dor <strong>de</strong> 5 fibras <strong>de</strong>ca<strong>de</strong>na nuclear.Las fibras intrafusales a<strong>de</strong>más <strong>de</strong> poseerinervaciones sensitivas también poseeninervación motora mediante pequeñasneuronas motoras <strong>de</strong>nominadasγmotoneuronas.Los axones <strong>de</strong> las γmotoneuronas tienenmenor diámetro que las <strong>de</strong> las α y son <strong>de</strong>dos tipos principales: fibras que terminan enforma <strong>de</strong> placa sobre los fascículoscontráctiles <strong>de</strong> las fibras <strong>de</strong> bolsa nuclear yotras, mas extendidas ( en forma <strong>de</strong> riel) queterminan en los elementos contráctiles <strong>de</strong>las fibras <strong>de</strong> ca<strong>de</strong>na nuclear.Los dos tipos morfológicos <strong>de</strong> fibrasfusimotoras, las terminaciones en placa y lasterminaciones en riel se correspon<strong>de</strong>rían ados clases <strong>de</strong> fibras fusimotorasfuncionalmente distintas. La <strong>de</strong>scarga <strong>de</strong> lasfibras fusimotoras estáticas aumentaría alaumentar la <strong>de</strong>scarga <strong>de</strong> las fibrassensoriales estáticas, mientras que las γmotoneuronas dinámicas respon<strong>de</strong>rían a la<strong>de</strong>scarga <strong>de</strong> las terminaciones sensorialesprimarias.Los husos musculares se encuentran enparalelo con la masa muscular y por lotanto, al estirarse el músculo ( y durante larelajación) también se estiran los husos.(No fue hasta que la actividad <strong>de</strong> estosreceptores se registró durante elestiramiento <strong>de</strong>l músculo que se pudo haceruna distinción entre terminaciones primariasy secundarias).Al realizar dicha investigación se observóque las <strong>de</strong>scargas <strong>de</strong> la terminal primaria <strong>de</strong>lhuso posee un componente <strong>de</strong> respuestadinámica <strong>de</strong> modo que durante elestiramiento muscular su <strong>de</strong>scarga <strong>de</strong>pen<strong>de</strong><strong>de</strong>l largo <strong>de</strong>l músculo y su velocidad <strong>de</strong>estiramiento. En cambio, la terminalsecundaria prácticamente no poseerespuesta dinámica. En condicionesestáticas, hay poca diferencia en el patrón<strong>de</strong> <strong>de</strong>scargas <strong>de</strong> ambas terminales.Los mecanismos <strong>de</strong> las diferentespropieda<strong>de</strong>s <strong>de</strong> las terminaciones primariasy secundarias no se compren<strong>de</strong>n bien perose estima que reflejan diferencias en las
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 44propieda<strong>de</strong>s viscoelásticas <strong>de</strong> las fibras <strong>de</strong>bolsa y ca<strong>de</strong>na nuclear.Es posible que el componente estático <strong>de</strong> la<strong>de</strong>scarga <strong>de</strong> la terminación primaria podría<strong>de</strong>rivar <strong>de</strong> su terminación sobre la fibra <strong>de</strong>ca<strong>de</strong>na nuclear mientras que el componentedinámico estaría mediado por su terminaciónsobre la fibra <strong>de</strong> bolsa nuclear.A partir <strong>de</strong>l huso muscular se origina elreflejo miotático, uno <strong>de</strong> los reflejos másimportantes para la mantención <strong>de</strong> lapostura.Este reflejo es mediado por la médulaespinal y originado por la activación <strong>de</strong> lasterminaciones primarias (Ia) ante elestiramiento musculartambién con los sinergistas, produciendo sucontracción.El reflejo miotático es mas importante en losmúsculos extensores mediales, que tienen unaimportante función anigravitatoria y, <strong>de</strong> este modo,este reflejo es un mecanismo básico <strong>de</strong> la postura.Por ej.: cuando una persona esta <strong>de</strong> pié, la fuerza <strong>de</strong>la gravedad tien<strong>de</strong> a estirar los músculoscuadríceps. El estiramiento <strong>de</strong> este músculo generael reflejo miotático que contrae dicho músculo yesto permite mantener la extensión a nivel <strong>de</strong> larodilla.Esto permite que la pierna funcione como un pilar<strong>de</strong> soporte y se contraponga al <strong>de</strong>splazamiento <strong>de</strong>lcuerpo impuesto por la fuerza <strong>de</strong> la gravedad.Como las terminaciones primarias poseen pocaadaptación, el reflejo pue<strong>de</strong> mantenerse durantemucho tiempo.Es muy importante señalar que el reflejomiotático es el mecanismo primario <strong>de</strong> lamantención <strong>de</strong>l tono muscular ya que el tonomuscular se <strong>de</strong>fine como : la resistencia<strong>de</strong> un músculo al estiramiento pasivo oactivo.(Fig.2: reflejos a partir <strong>de</strong>l huso muscular y elórgano tendinoso <strong>de</strong> Golgi ).Cuando se estira una masa muscular ( porejemplo al explorar el reflejo rotuliano conun martillo <strong>de</strong> reflejos), la terminaciónprimaria Ia que nace <strong>de</strong> los husos <strong>de</strong> lamasa muscular involucrada iniciapotenciales <strong>de</strong> acción.La fibra Ia penetra en la médula espinal,don<strong>de</strong> se bifurca dando origen a una ramaque ascien<strong>de</strong> por la columna dorsal y a otraque termina fundamentalmente en elsegmento medular en el que penetró estafibra .Las fibras que ascien<strong>de</strong>n daninformación sobre la velocidad <strong>de</strong>estiramiento muscular y sobre la longitud <strong>de</strong>lmúsculo a varias estructuras <strong>de</strong>l SNC, (Ej.cerebelo).La rama segmentaria, que queda en lamédula espinal se conecta <strong>de</strong> modomonosináptico con las α motoneuronas queinervan el músculo que dio origen al reflejo,no solo con los músculos protagonistas sinoEsto ocasiona una <strong>de</strong>formación <strong>de</strong> lasterminaciones primarias y secundarias en laregión central y pericentral <strong>de</strong> las fibrasintrafusales, creando un potencial <strong>de</strong>receptor en las fibras <strong>de</strong> tipo Ia o II que seoriginan en el huso.Por el contrario, la contracción <strong>de</strong>l músculoacortará el huso muscular y reducirá <strong>de</strong> estemodo las <strong>de</strong>scargas <strong>de</strong> las terminacionessensoriales que parten <strong>de</strong>l huso muscularOtros reflejos posturales integrados a nivelespinal están constituidos por losreflejos <strong>de</strong> soporte positivo ynegativo. Estos reflejos se obtienenestimulando con un <strong>de</strong>do la pata <strong>de</strong> animalesa los cuales se ha seccionadotransversalmente la médula espinal [animalespinal], y <strong>de</strong> esa manera se produce unaextensión <strong>de</strong> los miembros <strong>de</strong> los mismos (reacción <strong>de</strong> soporte positiva) . La<strong>de</strong>saparición <strong>de</strong>l reflejo se <strong>de</strong>nominareacción <strong>de</strong> soporte negativa. Por medio <strong>de</strong>lreflejo <strong>de</strong> reacción <strong>de</strong> soporte positiva, losanimales espinales pue<strong>de</strong>n permanecer <strong>de</strong>pie durante un tiempo <strong>de</strong> 2 a 3 minutosReflejos segmentarios ó reaccionesestáticas segmentarias: son aquellos
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 45reflejos en que el estímulo se origina en unmiembro y la respuesta refleja se produceen el miembro opuesto. Ej.: reflejo extensorcruzado que se produce ante una respuesta<strong>de</strong> flexión <strong>de</strong> <strong>de</strong>fensa <strong>de</strong>l miembro inferior.Reflejos generales: constituyen el grupomás gran<strong>de</strong> <strong>de</strong> reflejos posturales ycompren<strong>de</strong>n a aquellos reflejos en que unestímulo en un sitio particular <strong>de</strong>l cuerpoproduce respuestas en varias masasmusculares.Entre estos reflejos po<strong>de</strong>mos incluir a losreflejos generados en respuesta acambios en la posición <strong>de</strong> lacabeza. Estudiados en pacientes<strong>de</strong>corticados , se pue<strong>de</strong> observar que, algirar la cabeza <strong>de</strong>l paciente hacia la <strong>de</strong>rechacon respecto a la posición <strong>de</strong>l cuerpo <strong>de</strong>lpaciente , se producirá una extensión <strong>de</strong>lmiembro superior <strong>de</strong>l mismo lado y unaflexión <strong>de</strong>l lado opuesto y viceversa, si serota hacia el lado opuesto.La flexión <strong>de</strong> la cabeza, por otro lado, causaflexión <strong>de</strong> los miembros anteriores yextensión sostenida <strong>de</strong> los posteriores. Encambio la extensión <strong>de</strong> la cabeza, produceextensión <strong>de</strong> los miembros anteriores yflexión <strong>de</strong> los miembros posteriores.Reflejos <strong>de</strong> en<strong>de</strong>rezamiento: estánconstituidos por una serie <strong>de</strong> reflejosintegrados en su mayor parte a nivelmesencefálico que tienen como objetivoconservar una posición erecta normal con lacabeza levantada.Ante la inclinación <strong>de</strong> la cabeza a uno y otrolado (en reflejos estudiados en animales <strong>de</strong>experimentación <strong>de</strong>spués <strong>de</strong> seccionar eleje neural en el bor<strong>de</strong> superior <strong>de</strong>lmesencéfalo: [animal mesencefálico]);por excitación <strong>de</strong>l laberinto, el animalcontrae los músculos <strong>de</strong>l cuello buscandomantener la cabeza nivelada. Este reflejo se<strong>de</strong>nomina reflejo <strong>de</strong> en<strong>de</strong>rezamientolaberíntico.Si el animal es colocado sobre un costado,la presión sobre ese lado <strong>de</strong>l cuerpo iniciaun reflejo <strong>de</strong> en<strong>de</strong>rezamiento <strong>de</strong> la cabeza,aún con animales con laberintos <strong>de</strong>struidos.Este reflejo se <strong>de</strong>nomina reflejo <strong>de</strong>en<strong>de</strong>rezamiento <strong>de</strong> la cabezasobre el cuerpo.Fig.3: reflejos posturales en pacientes<strong>de</strong>corticadosEsta respuesta sería <strong>de</strong> origenlaberíntico (sus núcleos están ubicados anivel <strong>de</strong> bulbo raquí<strong>de</strong>o)La variación <strong>de</strong> posición <strong>de</strong> la cabeza, nosolo estimula el laberinto, sino también loshusos <strong>de</strong> músculos cervicales que serían losresponsables <strong>de</strong> los reflejos tónicos <strong>de</strong>lcuello. Estos reflejos han sido<strong>de</strong>mostrados en animales <strong>de</strong>scerebrados,los cuales: cuando su cabeza es giradahacia un lado, extien<strong>de</strong>n más rígidamentelos miembros <strong>de</strong> ese lado.Si la cabeza <strong>de</strong>l animal en estudio semantiene <strong>de</strong>recha y se inclina el cuerpo <strong>de</strong>lmismo, el estiramiento <strong>de</strong> los músculos <strong>de</strong>lcuello hace que estos se contraigan einicien una serie <strong>de</strong> reflejos miotáticos quepermiten el en<strong>de</strong>rezamiento <strong>de</strong>l tórax, elabdomen y los miembros. Este reflejo se<strong>de</strong>nomina reflejo <strong>de</strong>en<strong>de</strong>rezamiento <strong>de</strong>l cuello.Por otro lado, la presión sobre el cuerpo <strong>de</strong>lanimal, inicia reflejos <strong>de</strong> en<strong>de</strong>rezamiento<strong>de</strong>l mismo, aún cuando se mantenga sucabeza inclinada, por medio <strong>de</strong>l reflejo <strong>de</strong>en<strong>de</strong>rezamiento <strong>de</strong>l cuerpo sobreel cuerpo.Adicionalmente, en animales con cortezacerebral intacta, se producen reflejos <strong>de</strong>en<strong>de</strong>rezamiento ópticos, queen<strong>de</strong>rezan el cuerpo en ausencia <strong>de</strong>laberinto y <strong>de</strong> estimulación corporal.
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 46Reflejos posturales <strong>de</strong>pendientes <strong>de</strong>l nivelcortical son los reflejos <strong>de</strong> salto y <strong>de</strong>apoyo. El reflejo <strong>de</strong> salto permitemantener el equilibrio a un animal cuandoeste es empujado lateralmente y el reflejo <strong>de</strong>apoyo se hace evi<strong>de</strong>nte cuando un animalque estaba suspendido en el aire es puestoen contacto con una superficie. Ambosreflejos se alteran fuertemente cuando losanimales son <strong>de</strong>corticados.En resumen: los mecanismos reguladores<strong>de</strong> la postura son múltiples y compren<strong>de</strong>nuna gran cantidad <strong>de</strong> movimientos reflejosque son integrados a todos los niveles <strong>de</strong>lSNC:Médula espinalTallo encefálicoCorteza cerebral.Dichos reflejos están relacionados no solo con lapostura estática sino también con la generación <strong>de</strong>un fondo postural estable que permite larealización <strong>de</strong> los movimientos voluntarios y <strong>de</strong> losreflejos fásicos, dinámicos (ej. reflejo flexor <strong>de</strong><strong>de</strong>fensa para retirar un miembro <strong>de</strong> algún estímulodoloroso).BibliografíaCINGOLANI – HOUSSAY- FisiologíaHumana - 2000GANONG - Fisiología Médica - 18 a edición.BERNE R LEVY M - Fisiología a edición -2001EIZAGUIRRE – FIDONE. Fisiología <strong>de</strong>lSistema Nervioso. 1982.BEST Y TAYLOR - Bases Fisiológicas <strong>de</strong> lapráctica médica.– 2003GUYTON - Tratado <strong>de</strong> Fisiología Médica -10 a edición.
CATEDRA Nº 1 DE FISIOLOGIA HUMANA 2006 47PRINCIPALES REFLEJOS POSTURALESREFLEJO ESTIMULOS RESPUESTAS RECEPTORES INTEGRADO ENReflejo miotático Estiramiento Contracción muscular Husos musculares Médula espinal,bulboReacción <strong>de</strong>soporte positivaReacción <strong>de</strong>soporte negativaReflejos tónicos<strong>de</strong>l laberintoReflejos tónicos<strong>de</strong>l cuelloReflejos <strong>de</strong>en<strong>de</strong>rezamientolaberínticoReflejos <strong>de</strong>en<strong>de</strong>rezamiento<strong>de</strong>l cuelloReflejos <strong>de</strong>en<strong>de</strong>rezamiento<strong>de</strong>l cuerpo sobre lacabezaReflejos <strong>de</strong>en<strong>de</strong>rezamiento<strong>de</strong>l cuerpo sobre elcuerpoReflejos ópticos <strong>de</strong>en<strong>de</strong>rezamientoReacciones <strong>de</strong>apoyoReacción <strong>de</strong> saltoContacto con la plantao la palmaEstiramientoExtensión <strong>de</strong>l pie para sostenerel cuerpoLiberación <strong>de</strong> la reacción <strong>de</strong>soporte positivaGravedad Contracción <strong>de</strong> los músculosextensores <strong>de</strong> las extremida<strong>de</strong>sGiro <strong>de</strong> la cabezaHacia:1) un lado2) arriba3) abajoCambio en el patrón <strong>de</strong> lacontracción <strong>de</strong> los extensores1) extensión <strong>de</strong> los miembros<strong>de</strong>l lado al que se gira lacabeza2) las patas posteriores seflexionan3) las patas anteriores seflexionanPropioceptores enflexores distalesPropioceptores enextensoresOrganos otolíticosPropioceptores <strong>de</strong>l cuelloMédula espinalMédula espinalBulbo raquí<strong>de</strong>oBulbo raquí<strong>de</strong>oGravedad La cabeza se conserva nivelada Organos otolíticos MesencéfaloEstiramiento <strong>de</strong> losmúsculos en el cuelloPresión sobre un lado<strong>de</strong>l cuerpoPresión a un lado <strong>de</strong>lcuerpoEn<strong>de</strong>rezamiento <strong>de</strong>l tórax y <strong>de</strong>los hombros y luego <strong>de</strong> la pelvisHusos muscularesMesencéfaloEn<strong>de</strong>rezamiento <strong>de</strong> la cabeza Exteroceptores MesencéfaloEn<strong>de</strong>rezamiento <strong>de</strong>l cuerpo auncuando la cabeza se mantenga aun ladoExteroceptoresMesencéfaloSeñales visuales En<strong>de</strong>rezamiento <strong>de</strong> la cabeza Ojos Corteza cerebralVarias señalesvisuales,exteroceptivas ypropioceptivasDesplazamientolateral estando <strong>de</strong> pieEl pie se coloca sobre lasuperficie <strong>de</strong> apoyo parasoportar el cuerpoDa un salto para mantener losmiembros en posición parasoportar el cuerpo( Cuadro reproducido <strong>de</strong>: Fisiología Médica: William F. Ganong. 17ª edición)DiversosHusos muscularesCorteza cerebralCorteza cerebral













