Neurohistologia. Temario completo.pdf - VeoApuntes.com
Neurohistologia. Temario completo.pdf - VeoApuntes.com
Neurohistologia. Temario completo.pdf - VeoApuntes.com

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
Neurohistología<br />
<strong>Temario</strong> <strong><strong>com</strong>pleto</strong><br />
Universidad CEU San Pablo<br />
Alberto Gómez Esteban
Alberto Gómez Esteban<br />
Índice de contenidos<br />
Tema 1. Introducción al estudio de las neuronas____________________3<br />
Por Alberto Gómez y Laura del Olmo<br />
Tema 2. Células de la glía_____________________________________17<br />
Por Alberto Gómez y Laura del Olmo<br />
Tema 3. Sinapsis____________________________________________32<br />
Tema 4. La fibra nerviosa_____________________________________43<br />
Tema 5. Estructura de la médula espinal__________________________53<br />
Por Alberto Gómez y Laura del Olmo<br />
Tema 6. Histología del globo ocular______________________________64<br />
Tema 7. Histología del sistema auditivo__________________________81<br />
Tema 8. Histología del cerebelo_______________________________100<br />
Tema 9. Histología de la corteza cerebral________________________106<br />
2
Alberto Gómez Esteban<br />
Tema 1. Introducción al estudio de las neuronas<br />
Introducción<br />
El tejido nervioso es un tejido altamente especializado cuya principal función es integrar<br />
las funciones anatomofisiológicas.<br />
Sus células proceden del NEUROECTODERMO y van diferenciándose progresivamente<br />
en una serie de poblaciones celulares cuya función queda claramente delimitada:<br />
• NEURONAS (neurocitos). Son células excitables que se encargan de formar<br />
circuitos.<br />
• CÉLULAS GLIALES (gliocitos). Son células que proporcionan apoyo estructural y<br />
metabólico a las neuronas.<br />
El descubrimiento de las células del tejido nervioso arranca a mediados del siglo XIX,<br />
cuando a partir de los años 50 los microscopios aumentan progresivamente su resolución,<br />
y permiten mejorar la <strong>com</strong>prensión de la histología nerviosa.<br />
Se observaron las siguientes particularidades con respecto al tejido nervioso:<br />
• Tiene una singular citoarquitectura, debido a que, a diferencia de otros tejidos,<br />
tiene una matriz extracelular anómala.<br />
• Carece de tejido epitelial, ni glandular, ni conectivo, siendo una especie de “coto<br />
cerrado” con una densa circulación sanguínea.<br />
Además carece de circulación linfática y las células de su parénquima adoptan<br />
una morfología reticular.<br />
El aspecto reticular del sistema nervioso es el que más intrigó a los investigadores que<br />
<strong>com</strong>enzaron a estudiarlo.<br />
Se crearon dos corrientes acerca del estudio de la neurohistología:<br />
TEORÍA DE LA INDEPENDENCIA NEURONAL. Se trata de un concepto que surge<br />
a mediados del siglo XIX y parte de las descripciones de BAIKERS acerca de las<br />
neuronas.<br />
Esta teoría enuncia que las células neuronales son diferenciables entre sí, y<br />
tienen un principio y un final.<br />
3
Alberto Gómez Esteban<br />
TEORÍA DEL RETÍCULO NERVIOSO. Se trata de una teoría enunciada por GOLGI.<br />
Afirma que las células nerviosas forman un sincitio de forma reticular en el que<br />
no es posible diferenciar entre células.<br />
Sin embargo fue RAMÓN Y CAJAL el que consiguió mediante <strong>com</strong>plejas técnicas,<br />
demostrar que las neuronas tenían principio y final, demostrando la teoría de la<br />
independencia neuronal y la teoría de la polaridad dinámica.<br />
Durante el siglo XX se fue fortaleciendo el conocimiento acerca de la estructura neuronal<br />
gracias a los avances en microscopía electrónica, de modo que hemos estudiado las<br />
diferencias entre neuronas y glía entre otras cosas.<br />
Además los avances en inmunohistoquímica (IHQ) nos han permitido estudiar el tejido<br />
nervioso por sus marcadores neuronales.<br />
En definitiva hemos llegado a finales del siglo XX con un gran conocimiento<br />
ultraestructural y molecular acerca de las células del tejido nervioso.<br />
Estructura neuronal<br />
Soma neuronal<br />
El CUERPO o SOMA NEURONAL se trata del citoplasma que rodea la parte más<br />
prominente de la célula.<br />
Contiene un NÚCLEO especial, de gran tamaño y con nucléolo evidente, lo que denota<br />
su alto estado de diferenciación.<br />
La célula neuronal tras diferenciarse queda detenida en su estadio G 0 perdiendo su<br />
capacidad de división.<br />
Excepcionalmente existen neuronas capaces de multiplicarse, <strong>com</strong>o por ejemplo las<br />
neuronas de la mucosa olfatoria.<br />
Además, el soma contiene otros <strong>com</strong>ponentes citoplasmáticos:<br />
• PIGMENTOS CELULARES. Se acumulan en forma de gránulos en el citoplasma<br />
del soma, debido a que las neuronas tienen una vida tan larga que acumulan gran<br />
cantidad de productos de desecho.<br />
Estos gránulos forman una especie de coloración natural que se pueden teñir con<br />
sustancias específicas.<br />
Las sustancias que normalmente contienen estos gránulos son melanina,<br />
lipofuscina, gránulos neurosecretores…<br />
4
Alberto Gómez Esteban<br />
• APARATO DE GOLGI. La neurona es muy productiva a nivel molecular, y por<br />
ello contiene una gran maquinaria secretora que maneja a nivel subcelular la<br />
producción de moléculas que destina a sus prolongaciones celulares.<br />
El APARATO DE GOLGI y el RETÍCULO ENDOPLASMÁTICO se encuentran<br />
especialmente desarrollados en las neuronas debido a su necesidad funcional<br />
de producir moléculas.<br />
• LISOSOMAS. Son tanto primarios <strong>com</strong>o secundarios.<br />
Estos orgánulos contienen enzimas que se encargan de realizar funciones<br />
proteolíticas intra o extracelulares.<br />
• MITOCONDRIAS. Las neuronas tienen un número especialmente grande destinado<br />
a su gran producción de energía.<br />
Las neuronas son unas de las células más activas del organismo debido a que la<br />
descarga de potenciales de acción es un proceso enormemente costoso<br />
energéticamente.<br />
Las mitocondrias de la neurona, junto con su elevado consumo de oxígeno y<br />
glucosa están destinadas a surtir de energía a la célula neuronal.<br />
El elevado ritmo metabólico de la neurona tiene su contraparte negativa, ya que le<br />
supone un intenso estrés oxidativo.<br />
• CITOESQUELETO. Se trata de una estructura constituida por neurofibrillas.<br />
Las NEUROFIBRILLAS son haces gruesos de neurofilamentos que pueden<br />
llegar a alcanzar los 3 μm de grosor, por lo que son visibles con el microscopio<br />
óptico.<br />
Estas estructuras proporcionan el soporte de nuestras neuronas y sus<br />
prolongaciones.<br />
5
Alberto Gómez Esteban<br />
Los NEUROFILAMENTOS son estructuras muy largas capaces de sostener y<br />
mantener el dinamismo de las prolongaciones neuronales.<br />
Los NEUROTÚBULOS en cambio son estructuras pequeñas (25 nm) del<br />
citoesqueleto implicados en el transporte axonal y el mantenimiento de su<br />
estructura.<br />
• CUERPOS DE NISSL. Son una sustancia que si se observa por el microscopio<br />
óptico le aporta un aspecto tigroide a la neurona, debido a que forman un<br />
punteado basófilo por todo el soma.<br />
Consisten en cisternas aplanadas del RETÍCULO ENDOPLASMÁTICO LISO<br />
(REL) mezcladas con POLIRRIBOSOMAS que encontramos tanto en el soma <strong>com</strong>o<br />
en las dendritas.<br />
Encontramos cuerpos de Nissl en las dendritas y en el soma pero NUNCA en el<br />
axón.<br />
Prolongaciones celulares<br />
1. Dendritas<br />
Las DENDRITAS son procesos celulares muy ramificados con carácter dinámico, con<br />
neurotúbulos y neurofilamentos en su interior.<br />
La principal función de las dendritas es extender la influencia receptiva de la célula con<br />
el fin de ofrecer mayor superficie para que sinapten con ella.<br />
La neurona gracias a sus DENDRITAS expande su superficie a zonas periféricas al<br />
soma, haciéndola más receptiva a todo aquello que se deba transmitir al cuerpo celular.<br />
Las dendritas están estructuralmente reguladas por el citoesqueleto, y además tienen<br />
numerosos receptores funcionales.<br />
6
Alberto Gómez Esteban<br />
2. Axones<br />
El axón o CILINDROEJE es un proceso neuronal único (aunque algunas neuronas<br />
pueden tener 2 axones), cuya longitud es variable pero tiene un diámetro constante.<br />
La longitud del axón generalmente depende de la envergadura de la neurona.<br />
Su diámetro generalmente oscila entre 120 μm.<br />
Los axones son el mecanismo celular encargado de conducir el impulso nervioso a<br />
largas y medias distancias para <strong>com</strong>unicarse con otras estructuras.<br />
Podemos dividir al axón estructuralmente en varios segmentos, de más cercano al soma, a<br />
más lejano:<br />
• CONO AXÓNICO. Se trata de la parte del soma con forma de embudo del que sale<br />
el axón.<br />
• SEGMENTO INICIAL<br />
• TRONCO DEL AXÓN. Se trata del segmento intermedio. Es la parte más larga<br />
del axón.<br />
• RAMIFICACIONES COLATERALES<br />
• TELODENDRIA. Son ramificaciones más distales dentro del axón<br />
• BOTON TERMINAL. Son zonas engrosadas al final del axón implicadas en la<br />
sinapsis.<br />
Contienen vesículas llenas de neurotransmisores y un aparato sintético<br />
especializado.<br />
El axón contiene un especializado citoesqueleto con abundantes neurotúbulos y<br />
neurofilamentos así <strong>com</strong>o una membrana celular especializada que se denomina<br />
MEMBRANA AXÓNICA.<br />
Asimismo también contiene abundantes mitocondrias y un retículo endoplasmático<br />
liso muy especializado para la síntesis de moléculas.<br />
Los axones NO tienen cuerpos de Nissl.<br />
Las células de la glía proporcionan a la mayoría de axones una VAINA DE MIELINA que<br />
lo recubre para acelerar el impulso.<br />
La función de los axones es la de servir de medio de transporte desde el cuerpo celular<br />
hasta territorios alejados del mismo, por lo que tiene un transporte molecular<br />
especializado que se denomina FLUJO AXOPLASMÁTICO.<br />
7
Alberto Gómez Esteban<br />
El flujo axoplasmático puede ser bidireccional, existiendo estos tipos:<br />
• TRANSPORTE RETRÓGRADO. Se dirige de la periferia al soma celular.<br />
Este método de transporte proporciona al soma nutrientes y metabolitos, y es de<br />
tipo CELULÍPEDO (en dirección a la célula).<br />
Sirve para aportar al citoplasma principal sustancias captadas por el axón.<br />
• TRANSPORTE ANTERÓGRADO. Se dirige del cuerpo a la periferia axónica.<br />
Se trata de un movimiento CELULÍFERO/CELULÍFUGO (hacia fuera de la célula)<br />
para aportarle a partes distales del axón, sustancias sintetizadas en el propio soma.<br />
Existen varias velocidades para este transporte:<br />
Velocidad lenta (13 mm/día). Se trata de un flujo molecular estable, que<br />
se realiza de forma lenta y pasiva desde el soma a los botones axónicos.<br />
Está facilitado por el PERISTALTISMO AXÓNICO que lleva a cabo la vaina<br />
axonal.<br />
Velocidad rápida (100500 mm/día). Este transporte depende del<br />
citoesqueleto especializado del axón.<br />
Permite el transporte rápido de numerosas moléculas que sintetiza la<br />
eficiente maquinaria celular del soma, y que deben llegar a los extremos<br />
del axón.<br />
*Importancia del axón*<br />
En resumen, según lo que hemos visto, la importancia fundamental del axón reside en las<br />
siguientes funciones:<br />
Canaliza moléculas desde el soma que entrega a territorios periféricos.<br />
También mediante esta función mantiene el trofismo de otros órganos (por<br />
ejemplo la motoneurona α emite factores miotróficos vía transporte axonal).<br />
Se trata de un canal que recupera moléculas y las conduce al soma neuronal.<br />
8
Alberto Gómez Esteban<br />
El DESARROLLO AXONAL es un proceso fundamental, debido a que el axón se tiene<br />
que desarrollar en dirección a regiones distales muy concretas, para sinaptar con células<br />
específicas.<br />
Existen factores que estimulan el flujo axonal, y sirven para desarrollar los axones.<br />
Desde principios del siglo XX se sabe de la existencia de FACTORES NEUROTRÓFICOS<br />
(neurotrofinas) que son capaces de estimular el desarrollo axonal ya desde la época<br />
embrionaria, aunque no se conocía su naturaleza química.<br />
En los años 60 LEVI-MONTALCINI descubre las primeras neurotrofinas de naturaleza<br />
quimiotáctica.<br />
Los factores neurotróficos estimulantes del desarrollo axonal tienen entre sus acciones<br />
estimular la motilidad axonal mediante el quimiotactismo, mediante el cual la neurona<br />
envía su axón a otros territorios distales gracias a un gradiente de quimioquinas.<br />
El axón en desarrollo y el órgano diana con el que debe contactar presentan en su<br />
superficie moléculas de adhesión que les orientan espacialmente en su desarrollo.<br />
Durante los años 70-80 se fueron caracterizando 3 familias de factores neurotróficos:<br />
1. FACTORES NEUROTRÓFICOS. Estimulan tanto el desarrollo <strong>com</strong>o el flujo<br />
(transporte) axonal.<br />
2. FACTORES QUIMIOTÁCTICOS (quimioquinas)<br />
3. MOLÉCULAS DE ADHESIÓN ESPECÍFICAS<br />
Estas moléculas en conjunto modifican el crecimiento en longitud, y la orientación de<br />
éste crecimiento axonal sobre otros tipos celulares y territorios periféricos.<br />
9
Alberto Gómez Esteban<br />
Neurogénesis<br />
Ya hemos dicho que las neuronas forman el parénquima del sistema nervioso<br />
Se sabe que hay zonas del encéfalo que tienen la capacidad de reproducir progenitores<br />
neurogénicos de neuronas y de glía. No están muy claros pero se consideran una futura<br />
terapia para gente necesitada de más neuronas.<br />
Hay zonas del encéfalo (p.e. hipocampo en el giro dentado, o bien células<br />
subventriculares) que tienen PROGENITORES NEUROBLÁSTICOS, pero nuestro<br />
encéfalo está muy limitado a la hora de formar nuevas neuronas en la etapa adulta.<br />
Parece que la mayoría de neuronas nacen en la época fetal y se ubican en lugares<br />
donde sinaptan específicamente y se encuentran ahí durante toda la vida.<br />
Durante el desarrollo encefálico las neuronas se diferencian progresivamente. Hay tres<br />
estadios en el desarrollo entendido <strong>com</strong>o la diferenciación neuronal:<br />
1. NEUROGÉNESIS. Sigue los mismos principios morfogenéticos y de diferenciación<br />
que cualquier otro tejido (epitelial, glandular…).<br />
2. CRECIMIENTO DE LAS PROLONGACIONES. Se trata del desarrollo de axones<br />
y dendritas.<br />
Se dan otros principios distintos a los anteriores que se denominan<br />
MORFOGÉNESIS ESPECÍFICA DEL ENCÉFALO y se encarga de desarrollar<br />
estas prolongaciones y formar las conexiones entre diferentes neuronas.<br />
Aquí son importantes las rutas que guían las prolongaciones durante su<br />
migración de unos núcleos cerebrales a otros.<br />
Hay genes que en cada clase de neurona indican el número de prolongaciones y<br />
algunas conexiones<br />
3. REFINAMIENTO DE LAS CONEXIONES. Hace especificas las conexiones<br />
sinápticas (axodendríticas, axosómicas, etc…)<br />
Estas conexiones vienen determinadas por los propios axones que transmiten<br />
señales que son “sentidas” por las dendritas de otras neuronas cercanas, que toman<br />
estas señales <strong>com</strong>o factores de migración.<br />
10
Alberto Gómez Esteban<br />
Tipos de neuronas<br />
Clasificación morfológica<br />
La clasificación neuronal más elemental es la morfológica, es decir, la que hace referencia<br />
al número de prolongaciones:<br />
1. NEURONAS MULTIPOLARES. Tienen prolongaciones en todas las direcciones<br />
del espacio, y son las más frecuentes en el sistema nervioso central.<br />
Estas células tienen dendritas en todas direcciones en torno al soma celular y<br />
luego un único axón que dirige hacia una zona efectora.<br />
Suelen ser motoneuronas, pero también tenemos algunas que son interneuronas<br />
que en vez de dirigirse hacia un sitio concreto para una función muy específica,<br />
mantienen relaciones con otras neuronas para servir de intermediarias.<br />
Las neuronas multipolares adoptan muchas morfologías:<br />
Neuronas en cesto. Sus dendritas delimitan un hueco central que intenta<br />
abrazar a otras neuronas.<br />
Neuronas estrelladas. Tienen forma de araña y tienen dendritas en todas<br />
direcciones.<br />
Neuronas granulares. Se localizan por ejemplo en la capa granulosa del<br />
cerebelo. También emiten dendritas en todas direcciones<br />
Neuronas piramidales. Son motoneuronas triangulares en sección que<br />
desde la base de su pirámide emiten un axón que se dirige hacia abajo.<br />
Son propias de la corteza cerebral (capas III y V).<br />
Neuronas de Purkinje. Se localizan en la capa de células ganglionares del<br />
cerebelo y tienen gran tamaño (> 100 micras).<br />
11
Alberto Gómez Esteban<br />
Neuronas de los ganglios vegetativos<br />
Dentro de las neuronas multipolares existe otra clasificación en dos grupos:<br />
• NEURONAS GOLGI TIPO I. Tienen muchas dendritas y un axón largo,<br />
debido a que inervan estructuras muy distales.<br />
Un ejemplo típico son las motoneuronas cerebrales o medulares que<br />
recorren enormes distancias antes de establecer sinapsis.<br />
• NEURONAS DE GOLGI TIPO II. Tienen pocas dendritas y un axón corto<br />
debido a que su recorrido es de poca distancia.<br />
Un ejemplo típico son las neuronas granulares de la corteza cerebelosa.<br />
2. NEURONAS UNIPOLARES. Son lo contrario a las anteriores, ya que su cuerpo<br />
celular en forma de saco tiene una sola prolongación.<br />
Un ejemplo de este tipo de neuronas está en los fotorreceptores de la retina, con<br />
un cuerpo esférico y un axón corto.<br />
12
Alberto Gómez Esteban<br />
3. NEURONAS PSEUDOUNIPOLARES. Son neuronas que nacen con dos polos (de<br />
origen son bipolares) pero cuando se diferencian trasladan sus dos<br />
prolongaciones, juntándolas en una única prolongación que se ramifica en dos.<br />
Tienen un brazo central que se ramifica en dos.<br />
Las encontramos en las siguientes localizaciones:<br />
Ganglios cocleares<br />
Ganglios espinales<br />
4. NEURONAS BIPOLARES. Son células con una morfología bifuncional, con una<br />
dendrita a un lado y un axón al otro.<br />
Las encontramos en los nervios de la retina, del oído y de la zona olfatoria.<br />
13
Alberto Gómez Esteban<br />
Clasificación funcional<br />
La segunda forma de clasificarlas es funcional y muy sencilla:<br />
1. NEURONAS MOTORAS (motoneuronas). Envían ordenes eferentes desde el<br />
sistema nervioso central<br />
2. NEURONAS SENSITIVAS. Son neuronas que transportan información en<br />
dirección al sistema nervioso central<br />
3. INTERNEURONAS. Funcionan <strong>com</strong>o enlace entre otras neuronas.<br />
Clasificación neuroquímica<br />
La tercera clasificación es la clasificación neuroquímica, es decir, por el contenido<br />
molecular o criterio neuroquímico, que hace referencia a la distinción de las neuronas con<br />
respecto a sus neurotransmisores y hormonas.<br />
Este criterio es muy importante ya que ha permitido cartografiar el cerebro haciendo la<br />
quimioarquitectura celular del cerebro, es decir, la distribución neuronal en el encéfalo con<br />
respecto a sus moléculas de secreción.<br />
La ventaja de este criterio es que permite entender el funcionamiento del sistema<br />
nervioso y también hacer una aproximación neurofarmacológica de la estructura<br />
cerebral mediante agonistas y antagonistas de estas transmisiones químicas:<br />
1. NEURONAS COLINÉRGICAS. Utilizan acetilcolina (ACh) <strong>com</strong>o neurotransmisor.<br />
Se distinguen con anticuerpos que marcan la acetilcolinesterasa (¿o<br />
acetilcolintransferasa), que es una enzima necesaria para la síntesis de esta<br />
molécula.<br />
2. NEURONAS NORADRENÉRGICAS/ADRENÉRGICAS. Son las neuronas que<br />
producen noradrenalina y en menor cantidad adrenalina.<br />
Se las distingue con anticuerpos contra la feniletanolaminmetiltransferasa.<br />
3. NEURONAS SEROTONINÉRGICAS. Contienen serotonina (5-HT) <strong>com</strong>o<br />
neurotransmisor<br />
4. NEURONAS DOPAMINÉRGICAS. Utilizan dopamina <strong>com</strong>o neurotransmisor.<br />
5. NEURONAS GABAÉRGICAS. Utilizan GABA (ácido gamma-aminobutírico).<br />
Generalmente tienen funciones inhibitorias.<br />
14
Alberto Gómez Esteban<br />
6. NEURONAS PEPTIDÉRGICAS. Utilizan polipéptidos en la señalización, que entre<br />
otros serán los siguientes:<br />
Encefalinas<br />
Sustancia P<br />
Colecistoquinina (CCK)<br />
Otros<br />
Neurotensina<br />
7. NEURONAS AMINOACIDÉRGICAS. Utilizan secreciones de aminoácidos, entre<br />
otros los siguientes:<br />
Glicina<br />
Otros<br />
Aspártico<br />
8. NEURONAS DEL GRUPO DE RECEPTORES OPIÁCEOS. Utilizan péptidos<br />
opiáceos endógenos <strong>com</strong>o señalización.<br />
Cuando las neuronas pierden estos neurotransmisores hay que estimularlos con<br />
neurofármacos, pero cuando hay que anestesiar una vía se utilizan inhibidores de estas<br />
neuronas del encéfalo.<br />
15
Alberto Gómez Esteban<br />
16
Alberto Gómez Esteban<br />
Tema 2. Células de la glía<br />
Introducción<br />
Características generales de la glía<br />
Las células de la glía o GLIOCITOS son la población celular no neuronal del sistema<br />
nervioso.<br />
Representan más del 50% del peso del sistema nervioso, debido a que superan<br />
ampliamente a las neuronas en número.<br />
La proporción de neuronas:glía varía ampliamente según el territorio donde nos<br />
encontremos, pero se han llegado a contabilizar proporciones de 1:10 o incluso 1:50.<br />
Debido a estos motivos afirmamos que las células de la glía son la población celular<br />
predominante dentro del tejido nervioso.<br />
Tienen las siguientes características:<br />
Generalmente son de pequeño tamaño, aunque existen excepciones.<br />
Adoptan una morfología ramificada al igual que las neuronas, probablemente<br />
debido a que tienen el mismo origen embrionario (neuroectodermo).<br />
Se encuentran en alta densidad numérica, pudiendo acumularse en focos<br />
concretos.<br />
Se localizan entre neuronas, y las prolongaciones fibrosas de las mismas.<br />
Siguen manteniendo su capacidad proliferativa (a diferencia de las neuronas).<br />
Funciones de la glía<br />
Desde un punto de vista funcional, las células de la glía llevan a cabo los siguientes<br />
<strong>com</strong>etidos:<br />
1. Se encargan de proporcionar su sostén mecánico (estructural) al tejido<br />
nervioso.<br />
2. Se encargan de regular la nutrición de las neuronas<br />
3. Se encargan de regular el microambiente químico perineuronal, regulando<br />
asimismo la <strong>com</strong>posición iónica del medio.<br />
17
Alberto Gómez Esteban<br />
4. Se encargan de sintetizar las vainas de mielina, que protegen a las neuronas y las<br />
aíslan para aumentar la velocidad de su impulso.<br />
5. Tienen la <strong>com</strong>petencia defensiva del sistema nervioso, mediante dos funciones:<br />
‣ Regulan el acceso del sistema inmune al encéfalo.<br />
‣ Fagocitan los desechos celulares y moleculares que se acumulan en el<br />
intersticio del sistema nervioso<br />
6. Son las responsables de dirigir la regeneración de las fibras nerviosas dañadas,<br />
tanto en el sistema nervioso central, <strong>com</strong>o en el periférico.<br />
Clasificación general de la glía<br />
La clasificación de las células de la glía se realiza según su localización:<br />
CÉLULAS GLIALES DEL SISTEMA NERVIOSO CENTRAL<br />
• ASTROCITOS<br />
• OLIGODENDROCITOS<br />
Estos dos tipos de células forman la MACROGLÍA.<br />
• CÉLULAS EPENDIMARIAS (ependimocitos)<br />
• MICROGLÍA<br />
CÉLULAS GLIALES DEL SISTEMA NERVIOSO PERIFÉRICO<br />
• CÉLULAS DE SCHWANN<br />
• CÉLULAS SATÉLITE (capsulares)<br />
18
Alberto Gómez Esteban<br />
Astrocitos<br />
Los astrocitos son células gliales grandes (son un <strong>com</strong>ponente de la MACROGLÍA) que<br />
emiten numerosas prolongaciones periféricas.<br />
Sus prolongaciones abrazan los capilares del encéfalo para formar la MEMBRANA GLIAL<br />
LIMITANTE PERIVASCULAR.<br />
Se trata de un envoltorio pericapilar resultante de la acumulación de prolongaciones<br />
citoplasmáticas del astrocito en el entorno capilar<br />
Gracias a la vaina perivascular, los capilares del encéfalo no ponen en contacto cualquier<br />
tipo de metabolito o sustancia directamente con las neuronas, sino que gracias a esta<br />
membrana, regulan la disponibilidad molecular en el ambiente perineuronal<br />
En definitiva, se trata de un envoltorio que permite crear una barrera, y así la sangre no<br />
llega directamente a la neurona, sino que sufre un filtrado selectivo previo a través de la<br />
membrana glial limitante.<br />
Las PROLONGACIONES PERIVASCULARES regulan el acceso de moléculas<br />
desde los capilares a las neuronas.<br />
La membrana limitante glial perivascular estabiliza enormemente el microambiente<br />
molecular que rodea a la neurona, ya que si ésta envoltura no existiera, nuestras<br />
neuronas serían más vulnerables a los vaivenes plasmáticos.<br />
Los astrocitos también emiten prolongaciones hacia la superficie del encéfalo para formar<br />
la MEMBRANA GLIAL LIMITANTE SUBPIAL.<br />
La membrana glial limitante subpial (bajo la piamadre) es también muy importante debido a<br />
que representa un envoltorio citoplasmático para todo el ambiente neuronal, sobre<br />
todo en la corteza cerebral.<br />
Por último también emiten las PROLONGACIONES PERINEURONALES que son<br />
procesos citoplasmáticos que funcionan <strong>com</strong>o un envoltorio que rodea a las propias<br />
neuronas.<br />
Las prolongaciones perineuronales cumplirían, entre otras funciones, la de ser el puente<br />
entre la actividad eléctrica neuronal, y la regulación del flujo sanguíneo cerebral<br />
(unidad 24 de neurofisiología).<br />
19
Alberto Gómez Esteban<br />
*Resumen*<br />
El ASTROCITO emite tres tipos de prolongaciones:<br />
• MEMBRANA GLIAL LIMITANTE PERIVASCULAR. Está formada por<br />
prolongaciones que abrazan los capilares dentro del parénquima cerebral.<br />
La función de esta membrana es la de regular el paso de moléculas desde los<br />
capilares hasta el microambiente perineuronal.<br />
• MEMBRANA GLIAL LIMITANTE SUBPIAL. Está formada por prolongaciones que<br />
emiten hacia la superficie del encéfalo, bajo la piamadre.<br />
Representa una importante envoltura citoplasmática para todo el encéfalo pero<br />
especialmente para la corteza cerebral.<br />
• PROLONGACIONES NEURONALES. Está formada por prolongaciones que el<br />
astrocito emite hacia las propias neuronas.<br />
Cumplen funciones protectoras y nutricionales.<br />
La clasificación de los tipos de astrocitos se realiza en función del lugar hacia donde<br />
emitan sus prolongaciones:<br />
1. Astrocitos protoplasmáticos<br />
Representan entre el 15-25% del total de astrocitos y se localizan en la sustancia gris.<br />
Tienen procesos citoplasmáticos que contactan con los capilares y los somas<br />
neuronales.<br />
Algunos de ellos forman parte de la membrana glial subpial.<br />
20
Alberto Gómez Esteban<br />
2. Astrocitos fibrosos<br />
Son la variedad más numerosa, representando entre el 80-85% de todos los astrocitos.<br />
Se encuentran en la sustancia blanca y tienen un diámetro más pequeño que los<br />
astrocitos protoplasmáticos.<br />
Tienen prolongaciones muy finas, pero también bastante largas que rodean<br />
principalmente a los capilares.<br />
Sus prolongaciones suelen tener un número exagerado de gliofibrillas, las cuales<br />
explicaremos a continuación<br />
También suelen rodear por debajo a la piamadre (membrana glial subpial) y les<br />
proporcionan soporte metabólico a las fibras nerviosas.<br />
Las GLIOFIBRILLAS forman el citoesqueleto de las células gliales, encontrándose tanto<br />
en su cuerpo celular <strong>com</strong>o en sus prolongaciones.<br />
Son haces de neurofilamentos que mantienen la estructura del cuerpo celular y las<br />
prolongaciones citoplasmáticas, y contiene una clase especial de proteína: la PROTEÍNA<br />
GLIOFIBRILAR ÁCIDA (PGFA).<br />
La PGFA es de gran importancia ya que nos permite marcar astrocitos mediante técnicas<br />
de inmunohistoquímica.<br />
Los astrocitos fibrosos tienen una enorme cantidad de gliofibrillas en su citoesqueleto,<br />
mientras que se encuentran mucho menos exageradas en los astrocitos protoplasmáticos.<br />
21
Alberto Gómez Esteban<br />
Como curiosidad, en el hígado y en el páncreas hay un FIBROBLASTO PERIVASCULAR<br />
que está muy ramificado y se encarga de rodear capilares pancreáticos y hepáticos.<br />
Sus prolongaciones también tienen “gliofibrillas” y por tanto estas células serán PGFA + .<br />
Si se diferencian a miofibroblastos, dejan de expresar PGFA para <strong>com</strong>enzar a expresar<br />
la molécula de α-actina, propia de fibroblastos.<br />
Estas células <strong>com</strong>ienzan a producir matriz colágena, que predomina sobre el<br />
parénquima hepático dando lugar a CIRROSIS.<br />
Por tanto se puede determinar mediante técnicas de IHQ el inicio temprano de una cirrosis:<br />
Células PGFA + → Hígado sano<br />
Células α-actina + → Hígado cirroso<br />
*Resumen*<br />
Las principales diferencias existentes entre los astrocitos que hemos estudiado son<br />
las siguientes:<br />
• NÚMERO:<br />
Astrocitos protoplasmáticos. Minoritarios (15-25%)<br />
Astrocitos fibrosos. Mayoritarios (80-85%)<br />
• LOCALIZACIÓN:<br />
Astrocitos protoplasmáticos. Sustancia gris<br />
Astrocitos fibrosos. Sustancia blanca<br />
• PROLONGACIONES<br />
Astrocitos protoplasmáticos. Amplias pero cortas<br />
Astrocitos fibrosos. Delgadas y largas (parecidas a dendritas)<br />
• CITOESQUELETO<br />
Astrocitos protoplasmáticos. Pocas gliofibrillas<br />
Astrocitos fibrosos. Muchas gliofibrillas.<br />
22
Alberto Gómez Esteban<br />
3. Astrocito plasmofibrilar<br />
Se trata de un astrocito mixto entre protoplasmático y fibrilar.<br />
Estos astrocitos se localizan entre la sustancia gris y la sustancia blanca, y envían dos<br />
tipos de procesos:<br />
• Procesos protoplasmáticos. Son cortos pero amplios, <strong>com</strong>o sábanas.<br />
Envían estos procesos a la sustancia gris<br />
• Procesos fibrosos. Son más largos, pero muy delgados, similares a las dendritas<br />
neuronales.<br />
Envían estos procesos a la sustancia blanca.<br />
El hecho de que envíen los dos tipos de procesos, propios de cada uno de los astrocitos<br />
que hemos visto anteriormente, determina el hecho de que sean astrocitos híbridos.<br />
4. Astrocitos especiales<br />
Los hay de varios tipos:<br />
• ASTROCITOS EN VELO. Se localizan en la capa granular del cerebelo, rodeando a<br />
los glomérulos cerebelosos.<br />
Los glomérulos cerebelosos son estructuras neuronales que se encuentran en la<br />
capa granular, y están formados por sinapsis y capilares; los astrocitos en velo se<br />
encargarían de rodear ciertas neuronas asociadas a capilares en esa zona.<br />
• CÉLULAS DE MÜLLER. Se encuentran en la retina.<br />
• CÉLULA GLIAL DE BERGMAN. Se encuentran en el cerebelo.<br />
23
Alberto Gómez Esteban<br />
Funciones de los astrocitos<br />
Estas células cumplen las siguientes funciones dentro del sistema nervioso, englobadas<br />
dentro de las que hemos visto anteriormente:<br />
1. Son depredadores iónicos, es decir, captan iones de potasio y algunos otros aniones<br />
de tal forma que evitan desestabilizar la capacidad excitatoria de las neuronas<br />
Actúan <strong>com</strong>o estabilizadores al regular la disponibilidad iónica sobre la membrana<br />
neuronal.<br />
2. Se encargan del manejo o regulación de la glucosa.<br />
Las células gliales activadas por algunos neurotransmisores estimulan la<br />
glucogenolisis y por tanto se convierten en células productoras de glucosa para las<br />
neuronas.<br />
3. Sirven <strong>com</strong>o elemento de la BARRERA HEMATOENCEFÁLICA entre la sangre y la<br />
neurona ya que forma la cubierta perivascular.<br />
Los elementos de esta estructura son tres:<br />
ENDOTELIO CAPILAR. Los capilares del encéfalo son continuos, por lo que<br />
el paso de sustancias está muy limitado.<br />
Además las células endoteliales encefálicas están unidas mediante uniones<br />
adherentes.<br />
LÁMINA BASAL DEL ENDOTELIO. Lógicamente también será continua.<br />
PROCESOS BASALES DEL ASTROCITO. Como la membrana glial limitante.<br />
4. Son células altamente proliferativas con capacidad cicatricial. Son capaces de<br />
cicatrizar lesiones cerebrales.<br />
Los astrocitos estudiados en biopsias tumorales observamos que proliferan en torno a<br />
los tumores encefálicos para formar lo que se denomina reacción glial peritumoral<br />
(gliosis peritumoral).<br />
Desgraciadamente el tumor en el encéfalo se sobrepone a la contención glial y lo<br />
que hace es beneficiarse de las neurotrofinas y factores de crecimiento de las<br />
células gliales, lo que acelera el crecimiento tumoral.<br />
Los tumores cerebrales primitivos y secundarios (metástasis) se benefician de las<br />
reacciones defensivas del astrocito en el encéfalo.<br />
24
Alberto Gómez Esteban<br />
Oligodendrocitos<br />
Los oligodendrocitos son células pequeñas de citoplasma oscuro que se localizan tanto<br />
en la sustancia gris <strong>com</strong>o en la sustancia blanca.<br />
Tienen un diámetro de unas 8 μm, siendo células muy pequeñas, pero en <strong>com</strong>paración<br />
tienen un gran núcleo.<br />
El oligodendrocito es una célula pequeña con procesos citoplasmáticos cuya misión es<br />
formar las VAINAS DE MIELINA durante el proceso de recubrimiento axonal de las<br />
fibras nerviosas en el encéfalo.<br />
Los oligodendrocitos son exclusivos del sistema nervioso central, mientras que las<br />
células de Schwann cumplen su misma función (creación del recubrimiento de mielina)<br />
en el sistema nervioso periférico.<br />
Hay dos principales diferencias en cuanto a estas dos células:<br />
• LOCALIZACIÓN<br />
Oligodendrocito. Sistema nervioso central (SNC)<br />
Célula de Schwann. Sistema nervioso periférico (SNP)<br />
• PROLONGACIONES<br />
Oligodendrocito. Tiene 8-10 prolongaciones que se dirigen a diferentes<br />
fibras nerviosas para mielinizarlas.<br />
También puede dirigir todas sus prolongaciones a la misma fibra<br />
mielinizando sus distintos segmentos, pero de una sola fibra nerviosa.<br />
Célula de Schwann. Solo tiene una prolongación que dirige a un único<br />
segmento de una sola fibra.<br />
25
Alberto Gómez Esteban<br />
Existen dos tipos principales de oligodendrocito:<br />
• OLIGODENDROCITOS INTERFASCICULARES. Son los que se localizan en la<br />
sustancia blanca mielinizando axones.<br />
• OLIGODENDROCITOS PERINEURONALES. Se localizan en la sustancia gris.<br />
Realizan una función satélite para las neuronas, de forma parecida a <strong>com</strong>o<br />
hacían los astrocitos protoplasmáticos.<br />
Sus funciones son poco conocidas.<br />
Patologías de los oligodendrocitos<br />
Hay enfermedades encefálicas desmielinizantes que hacen que se pierda la cobertura<br />
mielínica de las fibras nerviosas en el sistema nervioso central.<br />
Estas enfermedades afectan a pacientes con mal tratamiento y producen una destrucción<br />
progresiva de la cobertura de la fibra nerviosa en el encéfalo produciendo un importante<br />
deterioro mental.<br />
Se piensa que conociendo las líneas celulares de los oligodendrocitos, algún día pacientes<br />
con enfermedades desmielinizantes recuperen la cobertura si se les inyecta progenitoras<br />
de oligodendrocitos que no estén afectadas por la enfermedad.<br />
Este proceso se conoce <strong>com</strong>o terapia celular del sistema nervioso y hay numerosas<br />
terapias destinadas a descubrir la localización en el encéfalo de estas progenitoras.<br />
Los progenitores de oligodendrocitos tienen un factor de transcripción que es el Olig2,<br />
que se encuentra exclusivamente operando en estas células y regula genes que luego<br />
intervienen en la función del oligodendrocito.<br />
Se ha descubierto una molécula de la superficie del oligodendrocito diferenciado que es<br />
un proteoglicano denominado NG2, el cual es un marcador de esta estirpe.<br />
Esta molécula también se encuentra en los progenitores de oligodendrocito y por ello<br />
las células NG2 + son las células que se buscan en terapias contra la desmielinización<br />
del SNC.<br />
Haciendo cultivos primarios de células gliales humanas podemos estudiar enfermedades<br />
neurológicas, degenerativas, carcinomas, etc…<br />
26
Alberto Gómez Esteban<br />
Microglía<br />
Es una célula muy importante en el encéfalo que representa el 20% de todas las células<br />
gliales encefálicas, y el 10-15% del total de células del sistema nervioso.<br />
La microglía es una población celular abundante con células de pequeño tamaño, que se<br />
sitúan cercanas a los capilares (microcirculación) del encéfalo, adoptan una<br />
morfología estrellada y están asociadas a estos capilares.<br />
Fueron descubiertas en 1880 por Nissl, y llamaron la atención debido a la presencia de<br />
fagosomas en su citoplasma.<br />
En 1920 Del Rio Ortega describió esta célula <strong>com</strong>o específica del sistema nervioso central,<br />
similar a un macrófago.<br />
En 1980 se demostró que tenían su origen en precursores hematopoyéticos, sin<br />
embargo a pesar de describirse este origen, también se ha demostrado que proceden en la<br />
época embrionaria del neuroectodermo, por tanto tienen un doble origen:<br />
‣ Origen inicial neuroectodérmico de la cresta neural<br />
‣ Origen secundario del tejido hematopoyético (sistema mononuclearfagocítico) de<br />
la médula ósea<br />
En el cerebro la microglía es el primer tipo celular que se activa cuando existe un<br />
trastorno.<br />
Por ejemplo la microglía se activará si tenemos una trombosis cerebral o una lesión<br />
traumática, o bien si padecemos enfermedades neurodegenerativas (Párkinson,<br />
Alzheimer), encefalitis…<br />
27
Alberto Gómez Esteban<br />
*Clínica*<br />
Si contraemos el virus de la rabia (por ejemplo, si nos muerde un perro infectado), este<br />
patógeno migra desde la laceración periférica, al sistema nervioso central mediante<br />
flujo axonal retrógrado.<br />
Infecta neuronas motoras en el SNC causando trastornos nerviosos. dando lugar<br />
finalmente a una encefalitis.<br />
La microglía <strong>com</strong>o decíamos detecta la patología del encéfalo, gracias a su dinamismo y a<br />
receptores:<br />
• Receptores para antígenos extraños<br />
• Receptores para moléculas de deshecho<br />
• Receptores para proteínas inflamatorias<br />
Además son muy sensibles al estrés oxidativo y a la hipoxia.<br />
Una vez activadas estas células se convertirán en la población que activa la defensa del<br />
encéfalo, en primer lugar proliferando en el territorio donde deben actuar, y coordinando<br />
un plan de acción estratégico en el que cumplen dos funciones:<br />
Activación de otras células gliales<br />
Protección neuronal<br />
La microglía se concentra en las zonas de lesión (por ejemplo tumores, donde proliferan<br />
localmente, o encefalitis, donde proliferan globalmente).<br />
Son células de citoplasma denso con fagosomas, numerosos lisosomas, cuerpos<br />
residuales y tienen la capacidad de presentar antígenos.<br />
También pueden secretar algunas neurotrofinas que van a activar a los astrocitos y a<br />
las neuronas, y además activan la proliferación de oligodendrocitos y el acceso de<br />
leucocitos al encéfalo.<br />
Normalmente al encéfalo no le conviene tener leucocitos en su parénquima debido a<br />
que liberan numerosos agentes oxidantes que pueden lesionar a las neuronas.<br />
La microglía fisiológicamente libera factores antimigratorios para que no entren al<br />
encéfalo los leucocitos, pero si existen lesiones graves, dejan entrar leucocitos.<br />
Los leucocitos que entran crónicamente al encéfalo producen desmielinizacion y<br />
degeneración neuronal.<br />
28
Alberto Gómez Esteban<br />
La microglía fagocita neuronas muertas, lo que genera desaparición progresiva y lenta<br />
de neuronas que <strong>com</strong>ienza a sentirse en la madurez (perdida memorial, de capacidad de<br />
aguante, etc…).<br />
Células ependimarias<br />
Las células ependimarias son células gliales que proceden del neuroectodermo, pero<br />
adoptan una morfología epitelial para revestir las cavidades del sistema nervioso central:<br />
Ventrículos cerebrales (superficie interna)<br />
Agujero ependimario. Se trata del canal central de la médula espinal.<br />
Las células ependimarias se encuentran revistiendo la superficie interna de cavidades<br />
que contienen líquido cefalorraquídeo.<br />
Estas células se superponen entre este líquido y el parénquima cerebral, actuando <strong>com</strong>o<br />
superficie reguladora del tránsito de moléculas entre el líquido cefalorraquídeo y el<br />
parénquima cerebral.<br />
Dividimos las células ependimarias en dos grandes poblaciones según su morfología:<br />
1. Ependimocitos<br />
Constituyen la mayoría de células ependimarias.<br />
Son células cúbicas con una morfología epitelial, pero no asientan sobre membrana<br />
basal.<br />
Están unidas entre ellas por uniones adherentes.<br />
Acumulan glucógeno en su citoplasma y tienen un aparato de Golgi grande, numerosos<br />
lisosomas, microfilamentos y además presentan cilios en su superficie.<br />
Debajo de estas células observamos prolongaciones subependimarias de otras células<br />
gliales que forman un lecho subependimario de prolongaciones de astrocitos.<br />
Los ependimocitos revisten el sistema ventricular, pero bajo ellos los astrocitos<br />
forman el LECHO SUBEPENDIMARIO.<br />
29
Alberto Gómez Esteban<br />
2. Tanicitos<br />
Se trata de la población minoritaria de células ependimarias.<br />
También adoptan una morfología cúbica, pero además tienen un proceso<br />
citoplasmático que se dirige hacia el interior del parénquima cerebral (proyecciones<br />
citoplasmáticas que se internalizan en el encéfalo) con los que rodean a los capilares.<br />
Los tanicitos al enviar estas proyecciones capilares y estando en contacto con el liquido<br />
cefalorraquídeo sirven para que difundan moléculas desde los capilares hasta el<br />
liquido y viceversa.<br />
Consideraciones generales<br />
Las células ependimarias constituyen un epitelio especializado que sirve <strong>com</strong>o interfase<br />
entre el líquido cefalorraquídeo y los capilares/parénquima cerebral.<br />
Estas células forman parte de la glía y proceden del neuroectodermo por lo que no son<br />
estrictamente epitelio, pero adoptan morfología y funciones epiteliales, por lo que se<br />
considera casi <strong>com</strong>o epitelio.<br />
La palabra glía significa “cola/pegamento” ya que aglutinan y organizan las neuronas<br />
Una de las funciones más importantes que tienen las células de la glía es la de formar la<br />
BARRERA HEMATOENCEFÁLICA.<br />
Esta estructura tiene tres <strong>com</strong>ponentes:<br />
• ENDOTELIO CAPILAR. Los capilares del encéfalo son continuos, por lo que el<br />
paso de sustancias está muy limitado.<br />
Además las células endoteliales encefálicas están unidas mediante uniones<br />
adherentes.<br />
• LÁMINA BASAL DEL ENDOTELIO. Lógicamente también será continua.<br />
• PROCESOS BASALES DEL ASTROCITO. Como la membrana glial limitante que<br />
ya hemos estudiado.<br />
30
Alberto Gómez Esteban<br />
31
Alberto Gómez Esteban<br />
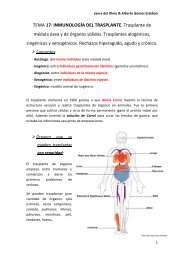
Tema 3. Sinapsis<br />
Introducción<br />
La sinapsis son contactos intercelulares entre neuronas o entre neuronas y células no<br />
neuronales, que sirven para la transmisión de la excitación entre dos o más células.<br />
Podemos clasificar las sinapsis en dos amplios grupos:<br />
• SINAPSIS ELÉCTRICAS. Son muy elementales y la estructura intercelular que permite<br />
desarrollarlas son los canales tipo GAP de <strong>com</strong>unicación directa.<br />
Se desarrollan entre las neuronas para integrarlas funcionalmente a través del<br />
intercambio de iones y de pequeñas moléculas.<br />
Gracias a las uniones intercelulares las neuronas se acoplan o se sincronizan<br />
funcionalmente, ya que las GAP permiten el flujo intercelular de iones y pequeñas<br />
moléculas entre citoplasmas adyacentes.<br />
Las uniones GAP están reguladas por hexámeros de conexina que crean conductos de<br />
baja resistencia eléctrica que se acoplan entre células vecinas y cuya permeabilidad se<br />
encuentra regulada por el calcio y por factores hormonales que regulan la<br />
conformación del conexón.<br />
Estas sinapsis no sirven para la señalización a larga distancia.<br />
• SINAPSIS QUÍMICAS. Son las que utilizan neurotransmisores para la propagación<br />
del mensaje eléctrico entre las neuronas o entre neuronas y miocitos por ejemplo.<br />
Cuando es entre neuronas los neurotransmisores sirven para transmitir la<br />
despolarización eléctrica entre células contiguas o conectadas.<br />
La sinapsis química transforma una señal eléctrica en una señal química que a su<br />
vez de nuevo produce una despolarización en la siguiente célula (señal eléctrica).<br />
La principal diferencia entre ambas sinapsis es que las químicas utilizan neurotransmisores<br />
<strong>com</strong>o señalizadores del mensaje eléctrico.<br />
32
Alberto Gómez Esteban<br />
Sinapsis química<br />
El mecanismo de funcionamiento de las sinapsis químicas consiste en la llegada de la<br />
despolarización de membrana hasta el lugar de la sinapsis (botón axónico).<br />
Esto aumenta la concentración de calcio intracelular, que da lugar a la liberación de<br />
las vesículas de neurotransmisor.<br />
El neurotransmisor interactúa con receptores en la siguiente célula lo que activa una<br />
señalización eléctrica en el elemento postsináptico, que puede darse por dos vías:<br />
• CANAL IÓNICO. El receptor de neurotransmisor está acoplado a un canal iónico<br />
que se abre ante la llegada del neurotransmisor.<br />
Esto puede provocar dos respuestas:<br />
Despolarización (activación)<br />
Hiperpolarización (inhibición)<br />
• RECEPTOR ACOPLADO A PROTEÍNA G. La llegada del neurotransmisor activa a<br />
la proteína G, que activa una cascada de transducción que entre otras cosas, abre<br />
un canal iónico<br />
La activación de estos receptores conlleva actividad eléctrica, pero además suele<br />
acarrear cambios a largo plazo en el elemento postsináptico.<br />
33
Alberto Gómez Esteban<br />
Sinapsis estándar<br />
En el ELEMENTO PRESINÁPTICO distinguimos el botón terminal de un axón, es decir,<br />
cuando el axón llega a la célula diana, se dilata en una estructura llamada botón axónico<br />
(0,5→4 μm de diámetro).<br />
El botón axónico contiene una membrana (axolema) que tiene una estructura <strong>com</strong>pleja<br />
asociada al retículo presináptico,<br />
El retículo presináptico es un conjunto de proyecciones moleculares densas que tienen<br />
un patrón geométrico hexagonal y permiten crear “pasillos” para el acceso de vesículas<br />
sinápticas hasta el axolema (membrana axonal contigua a la hendidura sináptica).<br />
Los pasillos que delimita el retículo presináptico se denominan SINAPTOPOROS, que son<br />
sitios activos por los que las vesículas sinápticas pasan hasta abrirse al exterior del<br />
botón axónico.<br />
34
Alberto Gómez Esteban<br />
Las vesículas sinápticas cuando llegan al sinaptoporo se abren por exocitosis y salen al<br />
exterior. Las vesículas sinápticas tienen un tamaño variable (4060 nm) y contienen en<br />
su interior neurotransmisores.<br />
Las VESÍCULAS SINÁPTICAS se clasifican en:<br />
• ESFÉRICAS (S). Suelen ser de morfología electrodensa. Contienen normalmente<br />
acetilcolina<br />
• APLANADAS (F). Suelen ser poco densas (electrolúcidas). Normalmente contienen<br />
GABA y glicina<br />
• VESÍCULAS DENSAS (C). También son esféricas. Contienen normalmente<br />
noradrenalina<br />
La clase de moléculas que contienen ofrece electronegatividad mayor o menor, por lo que<br />
estas descripciones no se basan en la concentración de neurotransmisor, sino en<br />
electrolucidez o electrodensidad.<br />
La entrada de calcio en el botón axónico, el neurotransmisor se vierte a un espacio<br />
denominado HENDIDURA SINÁPTICA que es un espacio que tiene una dimensión<br />
variable (en general es de pocos nanómetros), y es donde se concentra el<br />
neurotransmisor.<br />
El neurotransmisor actúa en la zona postsináptica sobre receptores específicos, los<br />
cuales variarán según el tipo de neurotransmisor. Podrán variar los siguientes parámetros:<br />
• MECANISMO DE ACCIÓN<br />
CANAL IÓNICO (catiónico o aniónico)<br />
PROTEÍNA G (normalmente activa la adenilato ciclasa)<br />
• EFECTO CELULAR<br />
Excitación (despolarización, o inducción a ella)<br />
Inhibición (hiperpolarización que dificulta la generación de nuevos<br />
potenciales)<br />
En el ELEMENTO POSTSINÁPTICO, el neurotransmisor provoca la entrada de iones<br />
(normalmente calcio) y en ocasiones la despolarización de dicha membrana,<br />
transmitiéndose un potencial de acción a lo largo de la célula activada.<br />
35
Alberto Gómez Esteban<br />
Dentro de mecanismo de receptores postsinápticos se conocen dos submecanismos que<br />
dependen del tipo de receptor de neurotransmisor:<br />
‣ Receptor asociado a un canal iónico. El efecto se da únicamente en forma de<br />
potencial de acción, y es un efecto rápido pero poco duradero.<br />
‣ Receptores asociados a proteínas G. El efecto del neurotransmisor media la<br />
activación de adenilato ciclasa, creando un efecto más lento pero también más<br />
prolongado.<br />
Las sinapsis son <strong>com</strong>plejos muy especializados que desarrollan las células para<br />
transmitir unidireccionalmente los impulsos nerviosos, y lo hacen desde las etapas<br />
más precoces del desarrollo intrauterino.<br />
La sinaptogénesis es progresiva desde que empezamos a desarrollar tejido neuronal.<br />
36
Alberto Gómez Esteban<br />
Clasificación de las sinapsis<br />
Clasificación por conexión<br />
Las sinapsis pueden ser:<br />
• AXOSOMÁTICAS. Las terminaciones axónicas contactan con el cuerpo celular o<br />
soma de otra neurona.<br />
• AXODENDRÍTICAS. Las terminaciones contactan con una dendrita de otra<br />
neurona.<br />
También debemos contemplar que no solo contacte con la dendrita, sino con sus<br />
espinas (CONTACTO AXOESPINOSO).<br />
• AXOAXÓNICAS. Un axón hace sinapsis sobre otro axón, para regular la liberación<br />
de neurotransmisores de éste.<br />
• DENDRODENDRÍTICAS. Son sinapsis entre dendritas de células contiguas.<br />
• DENDROSOMÁTICAS. Son menos frecuentes<br />
• SOMATOSOMÁTICAS. La propia superficie de la célula en ocasiones contacta<br />
con la superficie de una célula vecina.<br />
Esto amplifica mucho las posibilidades, permitiendo que las neuronas a través de la<br />
sinapsis ejerzan efectos excitatorios o inhibitorios sobre el terminal de contacto.<br />
Las sinapsis en ocasiones se clasifican según los neurotransmisores utilizados. Esto<br />
depende del contenido molecular de las vesículas almacenadas en el terminal axónico.<br />
Es <strong>com</strong>ún a todas las sinapsis que en general todas ellas propagan un mensaje eléctrico<br />
a través de un intermediario químico (neurotransmisor).<br />
La despolarización eléctrica que llega al terminal axónico desencadena la exocitosis de<br />
constituyentes químicos que a su vez de nuevo despolarizan la parte postsináptica de<br />
la estructura, para reemprender la transmisión eléctrica.<br />
También es preciso concebir que no haya otra despolarización, sino que la<br />
despolarización presináptica y el consiguiente neurotransmisor hiperpolaricen al<br />
terminal postsináptico, dificultando una nueva despolarización en él (sinapsis<br />
inhibitoria).<br />
37
Alberto Gómez Esteban<br />
Clasificación funcional<br />
Según GRAY las sinapsis se clasifican del siguiente modo:<br />
• SINAPSIS TIPO I. Son las clásicas sinapsis excitatorias<br />
• SINAPSIS TIPO II. Son de tipo inhibitorio.<br />
Las sinapsis inhibitorias tienen variaciones estructurales sobre el patrón general<br />
de una sinapsis química excitatoria:<br />
La HENDIDURA SINÁPTICA es mucho más estrecha (< 200 Ä)<br />
El APARATO SUBSINÁPTICO es menos grueso y más elemental.<br />
Las VESÍCULAS SINÁPTICAS son mucho más aplanadas y menos<br />
densas.<br />
Recordemos que las vesículas aplanadas y electrolúcidas contenían GABA y<br />
glicina, que son neurotransmisores clásicamente inhibitorios.<br />
Transporte de neurotransmisores<br />
Los neurotransmisores normalmente son sintetizados en el propio botón axonal<br />
presináptico.<br />
Además hay neurotransmisores que son sintetizados en el soma neuronal.<br />
En caso de los neurotransmisores sintetizados en el soma, deben ser conducidos del<br />
cuerpo neuronal al axón, lo que dependerá del TRANSPORTE ANTERÓGRADO.<br />
En este tipo de transporte tiene mucha importancia el mecanismo de flujo axonal que en<br />
general se halla regulado por una proteína (cinesina) que transporta las vesículas<br />
sinápticas hasta el botón terminal.<br />
La vía (dirección) que siguen las vesículas transportadas por cinesina es marcada por<br />
los microtúbulos del axón.<br />
Al mismo tiempo las VESÍCULAS SINÁPTICAS, que contienen neurotransmisor en su<br />
interior, presentan en su superficie constituyentes moleculares que son proteínas de<br />
anclaje vesicular.<br />
Las proteínas de anclaje vesicular sirven para reconocer a otras proteínas que están<br />
exclusivamente presentes en la membrana presináptica del terminal axónico, que<br />
además reconocen recíprocamente a las proteínas de la vesícula.<br />
38
Alberto Gómez Esteban<br />
Cuando llegan las vesículas por flujo axonal, las proteínas de anclaje vesiculares y las<br />
presinápticas regulan la fijación de las vesículas sinápticas a la membrana presináptica.<br />
Esto da lugar a que las vesículas queden dispuestas y que cuando el calcio aumente su<br />
concentración dentro del botón axónico se produzca la exocitosis por fusión de la<br />
membrana vesicular con la membrana presináptica.<br />
Cuando muchas vesículas se fusionan con el terminal, cabria esperar que la superficie<br />
presináptica aumentara muchísimo debido a que la membrana de las vesículas se sumaría<br />
a la del terminal, pero no ocurre así.<br />
Al mismo tiempo que se dan los procesos de exocitosis mediados por calcio, también se<br />
producen procesos de endocitosis de vesículas llenas de neurotransmisor.<br />
El neurotransmisor recaptado normalmente es degradado dentro del propio terminal<br />
axónico.<br />
Estos procesos obedecen a un ciclo reproductor que pone en la zona postsináptica<br />
neurotransmisores y los retira cuando han cumplido su función.<br />
En el botón axónico se hace necesaria la presencia de enzimas que degraden<br />
neurotransmisores y causan que el material pueda ser eliminado.<br />
Este mecanismo gasta energía y por ello los terminales axónicos tienen algunas<br />
mitocondrias, que van a abastecer de energía a todo el citoesqueleto que opera en<br />
este punto tan preciso de la prolongación neuronal.<br />
39
Alberto Gómez Esteban<br />
Sinaptogénesis<br />
La relación sináptica es dinámica y surge y desaparece en la propia fisiología neuronal, por<br />
lo tanto la sinaptogénesis se convierte en un hecho de gran importancia para la fisiología.<br />
Ganar o perder <strong>com</strong>plejidad en determinadas <strong>com</strong>unicaciones neuronales, es muy<br />
importante para el correcto funcionamiento cerebral.<br />
La SINAPTOGÉNESIS se trata de la formación de nuevas sinapsis dentro del tejido<br />
nervioso, sin necesidad de ganar o perder células neuronales para ello.<br />
Este proceso tiene gran importancia en muchas funciones cognitivas <strong>com</strong>o la memoria,<br />
en la cual, una parte muy importante se debe a modificaciones estructurales en las<br />
sinapsis (unidad 21 de neurofisiología).<br />
La sinaptogénesis es objeto de investigación en medicina. Este proceso es protegido y<br />
regulado por las CÉLULAS GLIALES, que regulan el desarrollo axonal y dendrítico.<br />
Las células de glía producen NEUROTROFINAS, <strong>com</strong>o el Factor de Crecimiento Neural<br />
(NGF).<br />
La sinaptogénesis depende de moléculas de adhesión que permiten a los axones<br />
reconocer el lugar donde deben establecerlas.<br />
Normalmente las moléculas de adhesión son de estos tipos:<br />
Superfamilia de las inmunoglobulinas<br />
Familia de las cadherinas<br />
Estas moléculas de adhesión tienen la misión de permitir que las prolongaciones<br />
axónicas o dendríticas reconozcan el entorno específico de otras células sobre las<br />
cuales harán sinapsis.<br />
De este modo se construyen <strong>com</strong>plejos circuitos que amplían el potencial<br />
neurospsíquico.<br />
40
Alberto Gómez Esteban<br />
Descripción de los tipos de sinapsis<br />
Antes hemos mencionado brevemente los tipos de sinapsis (axosomáticas, axodendríticas,<br />
axoaxónicas…) pero es preciso detenernos un poco para detallar más estas sinapsis.<br />
Sinapsis axosomáticas<br />
Dentro de este grupo hay tres subgrupos:<br />
Otros<br />
• BOTONES TERMINALES DE HELD. Estos botones terminales son puntos en los<br />
que el soma contacta con el centro del cuerpo.<br />
Esto es típico en sinapsis de células multipolares, <strong>com</strong>o las que se encuentran en<br />
el asta anterior (motora) de la médula espinal.<br />
• TERMINACIONES EN CÁLIZ. Se trata de axones que actúan <strong>com</strong>o si fueran<br />
pequeñas prolongaciones especializadas.<br />
Estas prolongaciones son la multiplicidad de la telodendria (ramificaciones<br />
axónicas distales, unidad 1) que abrazan al soma neuronal de otra célula.<br />
Esto crea un cesto que crea contactos axosomáticos con otra neurona.<br />
Estas terminaciones son frecuentes en el cerebelo.<br />
• TERMINACIONES EN CESTO. Se producen en el cerebelo <strong>com</strong>o contacto entre<br />
las células de la granulosa con células de Purkinje.<br />
En el cerebelo hay varias clases de SINAPSIS AXODENDRÍTICAS que se establecen en<br />
el entorno más íntimo entre las fibras trapezoides (trepadoras) y las dendritas de las<br />
células de Purkinje<br />
41
Alberto Gómez Esteban<br />
42
Alberto Gómez Esteban<br />
Tema 4. Fibra nerviosa<br />
Introducción<br />
Una de las principales diferencias que nos ofrece el sistema nervioso con respecto al<br />
resto de tejidos es que forma circuitos.<br />
Estos circuitos forman un sistema de conexiones que permiten la integración funcional<br />
del organismo.<br />
Si observamos estos circuitos a nivel microscópico, vemos que los haces de fibras<br />
nerviosas tienen unas características morfológicas muy similares en todo el sistema<br />
nervioso.<br />
Las FIBRAS NERVIOSAS son prolongaciones axonales de las neuronas envueltas por<br />
células de la glía especializadas:<br />
• OLIGODENDROCITOS. Forman la cubierta mielínica de los axones del sistema<br />
nervioso central (SNC)<br />
• CÉLULAS DE SCHWANN. Forman la cubierta mielínica de las fibras nerviosas de<br />
los nervios periféricos (SNP).<br />
Envoltura de mielina<br />
Aunque ya observamos en la unidad 2 las características generales de las células<br />
encargadas de formar la vaina de mielina, aquí lo ampliaremos algo más.<br />
Los OLIGODENDROCITOS emitían numerosas prolongaciones citoplasmáticas (¿20-<br />
50) que recubren pequeños segmentos de axones en el encéfalo.<br />
Este recubrimiento podía darse de dos formas:<br />
‣ Recubrimiento de un solo segmento de muchos axones<br />
‣ Recubrimiento de muchos segmentos de un solo axón<br />
En el sistema nervioso periférico los axones son mucho más largos y gruesos, y por<br />
ello la CÉLULA DE SCWANN envuelve un pequeño segmento de un solo axón, que<br />
puede llegar a medir varias micras.<br />
Para poder revestir estos axones, la célula de Schwann emite una prolongación<br />
citoplasmática que va a dar varias vueltas en torno al axón (hasta 50 vueltas).<br />
43
Alberto Gómez Esteban<br />
Una vez han envuelto al axón, obtienen un <strong>com</strong>pactado de su membrana plasmática,<br />
expulsando todo el contenido citoplasmático en dirección a su propio cuerpo.<br />
El citoplasma de la CÉLULA DE SCWANN queda en torno a su núcleo, mientras que la<br />
mayor parte de su membrana queda enrollada dando varias vueltas alrededor del axón;<br />
esta membrana es rica en esfingomielina.<br />
La MIELINA es el recubrimiento formado por estas células gliales, que además de<br />
proteger al axón, lo estabiliza electrofisiológicamente, permitiendo la CONDUCCIÓN<br />
SALTATORIA de los potenciales de acción.<br />
Entre varias células de Schwann contiguas existe una segmentación, es decir, un pequeño<br />
espacio del axón que no está recubierto por mielina, lo que denominamos NODO DE<br />
RANVIER.<br />
Los segmentos mielínicos entre los nodos de Ranvier se denominan SEGMENTOS<br />
INTERNODALES.<br />
Si realizamos una microfotografía ampliada de un nodo de Ranvier, observamos que las<br />
sucesivas hojas de la célula de Schwann se depositan a modo de lengüetas en el axón,<br />
en lo que se denomina ZONA PARANODAL que es ondulada.<br />
44
Alberto Gómez Esteban<br />
El NODO DE RANVIER es una zona del axón especialmente rica en canales iónicos de<br />
calcio, que se abren cuando llega el potencial de acción, contribuyendo a propagarlo a<br />
lo largo del axón de forma rápida<br />
En la imagen de la página anterior, podemos ver que el citoplasma principal de la célula<br />
de Schwann queda por encima del nodo de Ranvier.<br />
Las células de Schwann vecinas entran en contacto por encima del nodo de Ranvier y se<br />
unen entre sí mediante UNIONES ESTRECHAS HETEROTÍPICAS.<br />
Las sucesivas vueltas que da una misma célula de Schwann para formar un fascículo<br />
de la vaina de mielina se unen entre sí mediante UNIONES HOMOTÍPICAS, que vemos en<br />
el espesor de la vaina de mielina para mantener su cohesión.<br />
*Uniones*<br />
Relacionadas con las células de Schwann existen dos tipos de uniones:<br />
• UNIONES HETEROTÍPICAS. Se dan entre dos células de Schwann diferentes, por<br />
encima del nodo de Ranvier.<br />
• UNIONES HOMOTÍPICAS. Se dan en el espesor de una vaina de mielina creada por<br />
una única célula de Schwann.<br />
Las uniones homotípicas están destinadas a mantener la cohesión de la vaina de<br />
mielina.<br />
Determinadas enfermedades autoinmunes generan autoanticuerpos contra la claudina y<br />
ocludina, que son dos proteínas involucradas en la formación de las UNIONES<br />
HOMOTÍPICAS.<br />
Si se generan anticuerpos contra estas proteínas, las uniones quedan debilitadas, y se dan<br />
enfermedades autoinmunes desmielinizantes.<br />
La vaina de mielina se forma por el sucesivo enrollamiento de la prolongación<br />
citoplasmática de la célula de Schwann en torno al axón, lo que <strong>com</strong>o hemos dicho,<br />
provoca que el citoplasma que existía en esta prolongación se desplace al núcleo.<br />
Casi todo el citoplasma se desplaza al núcleo, pero hay zonas específicas en las que<br />
queda algo de citoplasma entre vueltas sucesivas; estas zonas se denominan<br />
INCISURAS DE SCHMIDT-LANTENAN y marcan una zona clara en el espesor de la<br />
vaina.<br />
45
Alberto Gómez Esteban<br />
La mielina <strong>com</strong>o hemos mencionado es una sucesión de vueltas de una célula en torno a<br />
un axón; esas vueltas implican a la membrana plasmática de la célula de Schwann, que<br />
tiene dos hemicaras:<br />
Hemicara citoplasmática. Se trata de la banda de la bicapa lipídica que se<br />
continúa con el citoplasma de la célula de Schwann.<br />
Hemicara externa. Se trata de la banda de la bicapa lipídica que se continúa con el<br />
medio extracelular.<br />
Estrechamente relacionadas con este concepto, si observamos el espesor de una vaina de<br />
mielina al microscopio, observaremos dos estructuras:<br />
• LÍNEA DENSA MAYOR. Se forma al aproximarse las dos hemicaras<br />
citoplasmáticas en una misma vuelta de la vaina de mielina.<br />
• LÍNEA DENSA MENOR. Se forma al aproximarse las dos hemicaras externas en<br />
una misma vuelta de la vaina de mielina.<br />
Además son las dos hemicaras externas las que establezcan entre sí las UNIONES<br />
HOMOTÍPICAS.<br />
Debido a que existen aproximadamente 50 vueltas de la vaina de mielina, habrá<br />
aproximadamente unas 100 líneas densas.<br />
Algunas células delimitan entre sus LÍNEAS DENSAS MENORES un pequeño espacio<br />
extracelular, por el cual pueden fluír iones y ponerse en contacto con el axón, aunque<br />
esto no es lo <strong>com</strong>ún.<br />
46
Alberto Gómez Esteban<br />
Recordemos que habia zonas delimitadas por LÍNEA DENSA MAYOR en las que<br />
existían espacios rellenos de una pequeña cantidad de citoplasma de la célula de<br />
Schwann; estos espacios se denominaban INCISURAS DE SCHMIDT-LANTENAN.<br />
Debido a la existencia de estos espacios, puede darse COMUNICACIÓN CELULAR<br />
HOMOTÍPICA entre los citoplasmas de los distintos niveles de la vaina de mielina.<br />
Estas zonas de <strong>com</strong>unicación expresan conexina-32, que se trata del monómero que en<br />
forma hexamérica formaría una unión <strong>com</strong>unicante GAP.<br />
Estas moléculas permiten que se desplacen iones del citoplasma más cercano al medio<br />
extracelular hasta el citoplasma más cercano al axón, y con ello a la membrana del axón.<br />
Si se crean autoanticuerpos contra la conexina-32 se producen procesos<br />
desmielinizantes <strong>com</strong>o la ESCLEROSIS MÚLTIPLE.<br />
Estructura de la fibra nerviosa<br />
Cuando una célula de Schwann rodea a un axón, según la forma en la que lo haga, puede<br />
crear dos tipos de fibra nerviosa:<br />
• FIBRA NERVIOSA MIELÍNICA. La célula de Schwann da varias vueltas en torno<br />
al axón, dandole una envoltura de mielina, cuyas características acabamos de<br />
estudiar.<br />
Cada célula de Schwann abraza un solo segmento de un axón, por lo que<br />
necesitaremos muchas células para mielinizar por <strong><strong>com</strong>pleto</strong> un solo axón.<br />
• FIBRA NERVIOSA AMIELÍNICA. La célula de Schwann se posiciona en torno al<br />
axón, pero no lo envuelve en toda su extensión.<br />
Existen fibras amielínicas en todo el sistema nervioso (SNC y SNP) y se<br />
caracterizan entre otras cosas por tener una velocidad de conducción del<br />
impulso mucho más lenta.<br />
47
Alberto Gómez Esteban<br />
Lesión de la fibra<br />
Las CÉLULAS DE SCHWANN además de proteger a los axones, y de aumentar su<br />
velocidad de conducción, también participan en la regeneración de una fibra nerviosa<br />
dañada.<br />
Estas células son fundamentales para regenerar una fibra nerviosa dañada, y son las<br />
principales artífices de que se pueda recuperar una cierta funcionalidad tras una lesión<br />
nerviosa.<br />
No sabemos muy bien las moléculas que utilizan estas células para regenerar axones<br />
dañados, pero actualmente sabemos que estas células sufren procesos de<br />
retrodiferenciación en lesiones axonales.<br />
Las células de Schwann en los procesos reparativos secretan numerosos factores de<br />
crecimiento y NEUROTROFINAS <strong>com</strong>o el factor de crecimiento neural.<br />
El FACTOR DE CRECIMIENTO NEURAL es un polipéptido que permite al axón dañado<br />
orientarse en dirección al órgano al que tienen que inervar, y por ello es una molécula<br />
fundamental de la regeneración neuronal, y para la migración de axones a través de<br />
nuestros tejidos.<br />
Cuando se lesiona un axón existen dos posibilidades:<br />
1. Lesión proximal<br />
El traumatismo se da en el inicio del axón, en la parte más cercana al soma neuronal, lo<br />
que conduce a una DEGENERACIÓN PROXIMAL de la fibra nerviosa.<br />
En estos casos el cuerpo neuronal sufre unos cambios funcionales (¿degeneración<br />
axonal) que conducen a alteraciones en la función de la neurona.<br />
Desde este momento <strong>com</strong>ienza la regeneración, ya que la célula fomenta el crecimiento<br />
de un nuevo axón que sustituye al otro, y debe recorrer una cierta longitud para<br />
inervar la estructura que inervaba previamente.<br />
Debido a esta longitud que debe recorrer, es necesaria la proliferacion de numerosas<br />
células de Schwann que orienten el crecimiento del nuevo axón.<br />
Este proceso dura algunas semanas desde que <strong>com</strong>ienza a crecer el axón, hasta que se<br />
restablece la inervacion del órgano denervado.<br />
48
Alberto Gómez Esteban<br />
2. Lesión distal<br />
El traumatismo se produce en algun punto del axón alejado del soma, lo que conduce a<br />
un proceso que llamamos DEGENERACIÓN DEL EXTREMO DISTAL (Walleriana).<br />
Tras esta lesión, la parte del axón que ha perdido el contacto con el soma se degenera, y<br />
la parte unida al soma <strong>com</strong>ienza a emitir prolongaciones que tratan de encontrar al<br />
órgano que inervaba originalmente.<br />
Las células de Schwann <strong>com</strong>ienzan a proliferar creando correderas cuyo <strong>com</strong>etido es<br />
guiar al axón en desarrollo hacia su órgano diana; estas células crean la BANDA DE<br />
BUNNER que se encarga de orientar el desarrollo axonal.<br />
La proliferación de células de Schwann en ambos tipos de lesión es estimulada por la<br />
acción de los macrófagos presentes en la zona distal, encargados de fagocitar los<br />
restos de la mitad del axón que perdió el contacto con el soma.<br />
Envueltas conectivas de la fibra nerviosa<br />
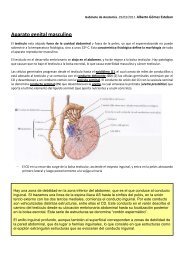
Endoneuro<br />
El endoneuro es una vaina de tejido conjuntivo reticular que rodea a cada fibra<br />
nerviosa individual.<br />
Las funciones del endoneuro son principalmente dos:<br />
Proporciona la matriz extracelular que rodea a la célula de Schwann.<br />
Además esta matriz conectiva se encarga de absorber los factores de<br />
crecimiento necesarios para que en un momento dado las células de Schwann<br />
entren en fase proliferativa.<br />
Proporciona el cemento conectivo que aglutina varias fibras nerviosas para<br />
formar un FASCÍCULO NERVIOSO.<br />
Esta envoltura conectiva se encarga de unir varias fibras nerviosas en un<br />
fascículo nervioso para que no se deshilachen.<br />
Cuando se lesiona una fibra nerviosa, el endoneuro que la rodea ya cuenta por sí mismo<br />
con los suficientes factores tróficos para reactivar la regeneración de la fibra, lo que explica<br />
la gran capacidad de regeneración de las lesiones de este tipo.<br />
El endoneuro además tiene capacidad para atraer al seno de los fascículos nerviosos<br />
capilares que se encargaran de la nutrición de las fibras nerviosas.<br />
49
Alberto Gómez Esteban<br />
Por este motivo, cuando se rompe el endoneuro en una lesión, se liberan factores<br />
angiogénicos que permiten que se forme nueva irrigación destinada a nutrir las células<br />
en regeneración.<br />
Los capilares que circulan por el interior de los fascículos nerviosos se encargan sobre<br />
todo de nutrir a las células de Schwann, para que estas sigan mielinizando de forma<br />
adecuada sus estructuras.<br />
Cuando se perturba la irrigación de un fascículo, <strong>com</strong>ienzan a aparecer defectos de<br />
conducción nerviosa debidos a la desmielinización causada por la muerte isquémica de<br />
las células de Schwann.<br />
Las células de Schwann no duran toda nuestra vida, al contrario que las neuronas, sino<br />
que se van sustituyendo, para lo cual es importante que reciban un aporte vascular<br />
estable.<br />
La vascularización no aporta únicamente nutrientes y oxígeno, sino que también existe un<br />
aporte de neurotrofinas, y permite el acceso del sistema inmune para que actúe en<br />
caso de inflamación.<br />
Perineuro<br />
El perineuro es otra clase de tejido conjuntivo que se organiza en torno a cada<br />
FASCÍCULO NERVIOSO.<br />
El perineuro no está tan especializado en la biología de la fibra nerviosa, sino que aglutina<br />
varios fascículos nerviosos para formar un NERVIO.<br />
Está formado por tejido conectivo fibroso denso, cuya función, además de aglutinar los<br />
fascículos nerviosos, es la de servir de lecho por el que discurren los vasos sanguineos<br />
que se encargan de nutrir al nervio internamente.<br />
Es un tejido muy vascularizado y con numerosos fibroblastos que se encargarían de<br />
regenerar el tejido conjuntivo perineural en caso de lesión del nervio.<br />
Esta envuelta es la que más se inflama en las NEUROPATÍAS, aunque para evitarlo los<br />
fibroblastos de este tejido liberan fisiológicamente factores antimigratorios, que evitan en<br />
condiciones normales el paso de leucocitos al interior del nervio.<br />
50
Alberto Gómez Esteban<br />
Epineuro<br />
El epineuro es una estructura que encontramos por encima del perineuro, rodeando a un<br />
NERVIO en forma de cubierta protectora.<br />
Se trataría de la estructura de tejido conjuntivo denso y fibroso que rodea más<br />
externamente a un nervio.<br />
Es similar a una aponeurosis debido a que está formado por tejido conectivo denso fibroso<br />
y apenas vascularizado cuya función es proteger al nervio e independizarlo de su<br />
entorno.<br />
*Resumen*<br />
1. El ENDONEURO rodea externamente a la fibra nerviosa.<br />
Está formado por tejido conectivo reticular<br />
Se encarga de aglutinar varias fibras nerviosas para formar un FASCÍCULO<br />
NERVIOSO.<br />
Además se encarga de reclutar capilares que nutran los <strong>com</strong>ponentes celulares del<br />
fascículo.<br />
2. El PERINEURO rodea externamente los fascículos nerviosos.<br />
Está formado por tejido conectivo denso fibroso vascular<br />
Se encarga de aglutinar todos los fascículos nerviosos para formar un NERVIO<br />
PERIFÉRICO.<br />
Además se trata del lecho por el que discurre la vascularización interna del nervio.<br />
3. El EPINEURO rodea externamente a los nervios periféricos.<br />
Está formado por tejido conectivo denso fibroso avascular.<br />
Se encarga de otorgar al nervio periférico protección mecánica y aislarlo de su<br />
medio.<br />
51
Alberto Gómez Esteban<br />
52
Alberto Gómez Esteban<br />
Tema 5. Estructura de la médula espinal<br />
Introducción<br />
La médula espinal se trata de una estructura nerviosa alargada a través de la cual el<br />
encéfalo se <strong>com</strong>unica con otras partes de nuestro cuerpo.<br />
Se localiza dentro del canal vertebral y se extiende desde el foramen magno (cráneo)<br />
hasta la zona donde se unen las vértebras L1-L2.<br />
En esa extensión encontramos un tejido nervioso que tiene una longitud de unos 43-45<br />
cm mientras que la propia columna vertebral consta de unos 73 cm.<br />
La MÉDULA ESPINAL no ocupa toda la extensión de la columna vertebral, sino que<br />
llega a L1-L2<br />
En su zona más inferior (L1-L2) la médula se divide en multitud de raíces nerviosas<br />
libres denominadas en su conjunto COLA DE CABALLO.<br />
Es algo más abajo (sobre L3-L4) donde realizamos las punciones lumbares debido a que el<br />
trócalo (aguja) no pinchará directamente la médula, sino un espacio lleno de raíces<br />
nerviosas que se apartan ante su paso.<br />
La médula es un ORGANO SEGMENTARIO y filogenéticamente es la estructura más<br />
antigua del sistema nervioso central.<br />
Cuando se desarrollaron los primeros animales, éstos estaban evolutivamente formados<br />
por segmentos iguales; en estos primeros animales apareció la médula espinal <strong>com</strong>o<br />
primer órgano integrador organizado en segmentos o METÁMERAS.<br />
Desde el punto de vista microanatómico es una estructura mucho más sencilla que el<br />
encéfalo pero no solo es el lugar de paso de los axones del encéfalo hacia los órganos<br />
sino que también es capaz de organizar respuestas por sí misma.<br />
Los REFLEJOS MIOTÁTICOS son coordinados de forma autónoma por la médula, y<br />
esto nos permite evaluar el estado funcional medular:<br />
Reflejo bicipital → Valoración de C5-C6<br />
Reflejo tricipital → Valoración de C6-C7<br />
Reflejo rotuliano → Valoración de L3-L4<br />
53
Alberto Gómez Esteban<br />
*Aclaración*<br />
Para hablar del reflejo rotuliano hemos dicho que valoraba el estado de la médula en sus<br />
niveles L3-L4, pero ya sabemos que la médula deja de existir <strong>com</strong>o tal a nivel de L2.<br />
Cuando hablamos de niveles L3-L4 nos referimos a los niveles medulares cuyas raíces<br />
nerviosas emergen por dichos niveles.<br />
Hablando de la rótula, donde se inserta el músculo cuádriceps, probablemente los niveles<br />
medulares cuyo nervio espinal sale por L3-L4 se encuentren en los primeros niveles<br />
lumbares.<br />
Una lesión medular da lugar a pérdidas de sensibilidad que se originan desde el nivel de<br />
la lesión hasta los niveles inferiores; además existe parálisis para movimientos<br />
voluntarios.<br />
La médula, <strong>com</strong>o ya hemos dicho, tiene dos límites anatómicos:<br />
• Límite superior. Se continúa con el encéfalo a nivel del bulbo raquídeo (médula<br />
oblongada).<br />
Se considera que el bulbo pasa a ser médula a partir de la DECUSACIÓN<br />
PIRAMIDAL.<br />
• Límite inferior. La médula inferiormente termina en una especie de cono (CONO<br />
MEDULAR) que es el origen de numerosas fibras nerviosas.<br />
El vértice del cono medular se encuentra a nivel de L1-L2.<br />
La médula está <strong>com</strong>puesta por 31 segmentos o METÁMERAS que se corresponden con<br />
nervios espinales.<br />
Los nervios espinales abandonan el canal medular por los agujeros o canales<br />
paravertebrales para inervar sus estructuras de destino.<br />
54
Alberto Gómez Esteban<br />
Estructura general de la médula<br />
Si hacemos un corte transversal a la médula nos damos cuenta de que hay dos zonas bien<br />
diferenciadas:<br />
• SUSTANCIA BLANCA. Se sitúa en la periferia de la médula espinal.<br />
Está formada por axones mielinizados tanto ascendentes <strong>com</strong>o descendentes.<br />
• SUSTANCIA GRIS. Se sitúa en el centro de la médula espinal.<br />
Al igual que en todo el sistema nervioso central, está <strong>com</strong>puesta de somas,<br />
dendritas e interacciones sinápticas.<br />
Estructura externa<br />
La médula espinal vista exteriormente presenta un aspecto blanquecino, debido a que<br />
está <strong>com</strong>puesta superficialmente por fibras mielínicas.<br />
‣ La médula periféricamente está <strong>com</strong>puesta por SUSTANCIA BLANCA<br />
‣ La médula en su centro está <strong>com</strong>puesta por SUSTANCIA GRIS<br />
Se continúa por en niveles superiores con el tronco del encéfalo a nivel del bulbo raquídeo.<br />
Inferiormente el CONO MEDULAR está unido al ligamento coccígeo (filum terminale)<br />
que une el final de la médula al hueso sacro.<br />
El LIGAMENTO COCCIGEO se trata de una continuación de la piamadre que rodea la<br />
médula y está formado por colágeno y células de la glía.<br />
La médula queda fija dentro del canal medular gracias a tres estructuras:<br />
Raíces nerviosas, que fijan la médula a cada agujero paravertebral<br />
Ligamento coccígeo, que fija la médula al hueso sacro (inferiormente)<br />
Bulbo raquídeo, que fija la médula al encéfalo (superiormente)<br />
La médula espinal es un cilindro de 0’81cm de diámetro, pero este diámetro no es<br />
estable en toda su extensión, sino que existen 2 engrosamientos o intumiscencias:<br />
• INTUMISCENCIA CERVICAL. Se corresponde a la zona medular cuyas raíces<br />
nerviosas conforman el plexo braquial.<br />
• INTUMISCENCIA LUMBAR. Se corresponde a la zona medular cuyas raíces<br />
nerviosas conforman el plexo lumbosacro.<br />
55
Alberto Gómez Esteban<br />
Observamos pues que las intumiscencias se corresponden a zonas donde es preciso<br />
inervar abundante musculatura voluntaria, <strong>com</strong>o ocurre en las extremidades.<br />
Además la médula no es un cilindro liso, sino que está marcada por fisuras y surcos:<br />
• FISURA VENTRAL. Es muy profunda y contiene los vasos espinales anteriores:<br />
Arteria espinal anterior<br />
Venas espinales anteriores (2)<br />
• FISURA DORSOMEDIAL<br />
Estos dos surcos son los más grandes y dividen la médula en 2 mitades (hemimédulas)<br />
sagitalmente.<br />
Además de estos existen otros surcos menores:<br />
• SURCO VENTRO-LATERAL. Se trata de la zona de entrada de la raíz posterior<br />
(sensitiva)<br />
• SURCO DORSO-LATERAL. Se trata de la zona de salida de la raíz anterior<br />
(motora)<br />
• SURCO INTERMEDIO-DORSAL (sólo en la región cervical)<br />
Estos surcos ayudan a delimitar cordones medulares dentro de la sustancia blanca.<br />
56
Alberto Gómez Esteban<br />
Cordones medulares<br />
Los cordones medulares son zonas de SUSTANCIA BLANCA que representan la zona<br />
más periférica de la médula.<br />
Están delimitados por los surcos que acabamos de ver:<br />
• CORDÓN ANTERIOR. Se ubica entre dos surcos:<br />
Fisura ventral (la más grande)<br />
Surco ventrolateral<br />
• CORDÓN LATERAL. Se ubica entre dos surcos:<br />
Surco ventrolateral<br />
Surco dorsolateral<br />
• CORDÓN POSTERIOR. Se ubica entre dos surcos:<br />
Surco dorsomedial (grande)<br />
Surco dorsolateral<br />
57
Alberto Gómez Esteban<br />
Sustancia gris<br />
La sustancia gris de la médula adopta una forma de “mariposa” o de “H”. En la parte<br />
media encontramos el canal central de la médula espinal (canal ependimario), que fuera<br />
en su día el tubo neural.<br />
La sustancia gris que rodea el epéndimo se denomina SUSTANCIA GRIS INTERMEDIA.<br />
En general, la sustancia gris medular está <strong>com</strong>puesta por dos grandes partes:<br />
• ASTA ANTERIOR. Anatómicamente tiene una base (más medial) y una cabeza<br />
(más anterior).<br />
Funcionalmente es un asta somatomotora, ya que está constituida por neuronas<br />
que inervan musculatura somítica.<br />
Sus axones salen por las raíces anteriores.<br />
• ASTA POSTERIOR. Anatómicamente tiene una base (medial), un cuello<br />
(intermedio) y una cabeza (dorsal), y es más larga que el asta anterior.<br />
Funcionalmente el asta posterior es sensorial y es donde están las neuronas<br />
sensoriales de la médula espinal.<br />
Estas neuronas reciben impulsos del resto del organismo por las raíces<br />
posteriores.<br />
Ambas astas están unidas por una banda transversal de sustancia gris que denomina<br />
COMISURA GRIS.<br />
58
Alberto Gómez Esteban<br />
La sustancia gris proporcionalmente varía en los distintos niveles medulares:<br />
• MÉDULA CERVICAL. En este nivel es muy abundante.<br />
Sobre todo observamos que el asta anterior está muy aumentada debido a la<br />
elevada densidad de motoneuronas encargadas de inervar el miembro superior.<br />
Este engrosamiento constituye la INTUMISCENCIA CERVICAL.<br />
• MÉDULA TORÁCICA. En este nivel es menos abundante.<br />
El asta anterior de la médula torácica es pequeña, pero en cambio encontramos en<br />
toda su extensión un acumulo lateral de sustancia gris que forma el ASTA<br />
LATERAL.<br />
El asta lateral contiene neuronas preganglionares del sistema nervioso<br />
simpático.<br />
• MÉDULA LUMBAR. En este nivel vuelve a ser muy abundante.<br />
Observamos de nuevo el crecimiento del asta anterior por el mismo motivo que a<br />
nivel cervical, es decir, aumenta el número de motoneuronas para inervar el<br />
miembro inferior.<br />
Este engrosamiento constituye la INTUMISCENCIA LUMBAR.<br />
Además proporcionalmente la SUSTANCIA BLANCA va descendiendo en cantidad<br />
conforme vamos descendiendo a niveles más sacros.<br />
Esto se debe a que los fascículos descendentes desde el encéfalo se van agotando, y<br />
habrá menos fascículos ascendentes, que se incorporan progresivamente desde<br />
niveles inferiores.<br />
Sabiendo esto, nos es posible distinguir entre niveles medulares en un corte transversal,<br />
únicamente atendiendo a tres parámetros:<br />
1. Presencia o no de ASTA LATERAL (C8-L3)<br />
2. Tamaño del ASTA ANTERIOR<br />
3. Proporción de SUSTANCIA BLANCA<br />
59
Alberto Gómez Esteban<br />
Citoarquitectura de la médula<br />
Asta anterior<br />
Es donde se ubican principalmente las neuronas motoras grandes:<br />
• MOTONEURONAS. Son neuronas piramidales grandes, que pueden adoptar una<br />
morfología o bien triangular o bien estrelladas.<br />
Son neuronas GOLGI TIPO I con un axón largo.<br />
Hay de tres clases:<br />
MOTONEURONAS RADICULARES. Sus axones forman parte de la raíz<br />
anterior.<br />
También se denominan motoneuronas α.<br />
MOTONEURONAS CORDONALES. Se denominan así porque sus axones<br />
se introducen en los cordones de sustancia blanca próximos para poder<br />
asociar diferentes niveles medulares.<br />
MOTONEURONAS PEQUEÑAS. Solo tienen entre 10-20 μm de diámetro.<br />
También se llaman motoneuronas γ y se encargan de inervar husos<br />
neuromusculares.<br />
• NEURONAS DE RENSHAW. Son motoneuronas de axón corto que reciben<br />
impulsos de las motoneuronas que hemos visto anteriormente.<br />
Son inhibidoras y se encargan de controlar los trenes de impulsos de las<br />
motoneuronas grandes.<br />
Comisura gris<br />
Esta sustancia gris se localiza en el límite de ambas astas.<br />
Contiene principalmente neuronas de asociación de los siguientes tipos:<br />
• NEURONAS INTERCALARES (internunciales). Se encargan de asociar las<br />
neuronas del asta posterior con las del asta anterior del mismo lado.<br />
Son neuronas GOLGI TIPO II<br />
También se denominan neuronas de ajuste ya que tienen memoria para modular<br />
las funciones del asta anterior y asta posterior (memorizan sistemas de control).<br />
60
Alberto Gómez Esteban<br />
• NEURONAS COMISURALES. Se encargan de conectar las neuronas de astas de<br />
lados opuestos (una hemimedula con la otra).<br />
Asta posterior<br />
Mayoritariamente contiene neuronas receptoras de axón corto que se encuentran<br />
influidas por impulsos aferentes de las raíces posteriores.<br />
Anatómicamente distinguimos tres partes:<br />
• BASE. Es la zona más medial.<br />
Contiene principalmente neuronas que reciben SENSIBILIDAD INTEROCEPTIVA<br />
(sensitivos del interior del organismo) desde órganos procedentes del endodermo<br />
(vísceras)<br />
• CUELLO. Se trata de la región intermedia, posterior a la base.<br />
Recibe principalmente ESTÍMULOS PROPIOCEPTIVOS de tejidos procedentes del<br />
mesodermo (huesos, músculos y ligamentos)<br />
• CABEZA. Es la zona más distal del asta posterior.<br />
Recibe principalmente ESTÍMULOS EXTEROCEPTIVOS de órganos procedentes<br />
del ectodermo (piel y tejido subcutáneo).<br />
*Recordatorio*<br />
El soma de las neuronas que penetran por las raíces posteriores se encuentra en el<br />
GANGLIO ESPINAL que se encuentra en una posición posterolateral con respecto a la<br />
médula.<br />
Las neuronas del ganglio espinal son pseudounipolares.<br />
61
Alberto Gómez Esteban<br />
Asta lateral<br />
Es una zona relacionada con el SISTEMA NERVIOSO AUTÓNOMO, y la encontraremos<br />
únicamente en dos localizaciones:<br />
• SEGMENTOS C8-L3. En esta zona también se denomina sustancia gris<br />
intermediolateral.<br />
Aquí encontramos los somas preganglionares de neuronas del sistema nervioso<br />
simpático.<br />
• SEGMENTOS S2-S4. Aquí encontraremos neuronas del sistema nervioso<br />
parasimpático.<br />
62
Alberto Gómez Esteban<br />
63
Alberto Gómez Esteban<br />
Tema 6. Histología del globo ocular<br />
Introducción<br />
Los animales hemos desarrollado un órgano de la visión que enfoca la luz en una zona<br />
donde ésta puede ser convertida en patrones eléctricos.<br />
Embriológicamente el globo ocular deriva de proyecciones encefálicas del diencéfalo<br />
anterior que forman las VESÍCULAS ÓPTICAS, que quedan unidas al encéfalo mediante<br />
el tallo óptico, que es hueco.<br />
Las vesículas ópticas obtienen un recubrimiento procedente del ectodermo de la<br />
superficie de la cabeza que van a formar la CÓRNEA.<br />
Además el globo ocular recibe una invasión mesenquimal que formará finalmente la<br />
CAPA MEDIA VASCULAR del globo ocular.<br />
Una vez <strong>com</strong>pletado su desarrollo, consideramos que el globo ocular está formado por tres<br />
partes principales:<br />
• PARTE EXTERNA. Se trata de la capa procedente del ectodermo superficial y<br />
tiene naturaleza epitelial.<br />
Está formada por dos partes:<br />
CÓRNEA<br />
ESCLERÓTICA<br />
Su función es la de proteger el globo ocular.<br />
• CAPA MEDIA. Se trata de la capa derivada del tejido mesenquimal.<br />
64
Alberto Gómez Esteban<br />
Consta de tres elementos que se engloban con el nombre de ÚVEA:<br />
IRIS. Es la parte más anterior.<br />
Se trata de un diafragma muscular que regula la entrada de luz al interior<br />
del globo ocular.<br />
CUERPOS CILIARES. Es una parte intermedia<br />
Son entre otras cosas el órgano encargado de secretar humor acuoso.<br />
COROIDES. Es la parte más posterior.<br />
Se trata de una capa vascular cuya misión es nutrir la retina.<br />
• CAPA INTERNA. Se trata de la capa derivada del neuroectodermo del<br />
diencéfalo.<br />
Se trata de la capa fotosensible o RETINA.<br />
Está <strong>com</strong>puesta por una serie de capas celulares que incluyen neuronas y células<br />
de la glía.<br />
Partes del ojo:<br />
Rojo: Derivados neuroectodérmicos<br />
Naranja: Derivados mesodérmicos<br />
Azul: Derivados ectodérmicos<br />
65
Alberto Gómez Esteban<br />
Capa externa<br />
1. Córnea<br />
La córnea es la estructura más anterior del globo ocular.<br />
Es una estructura transparente y convexa de 0’81’1 mm de grosor, con un radio de<br />
curvatura pequeño y un diámetro de 1 cm.<br />
En su zona más lateral se continúa con la ESCLERA mediante una estructura llamada<br />
limbo esclerocorneal.<br />
Forma parte del sistema dióptrico de enfoque luminoso, con un índice de refracción de<br />
1’36 y por este motivo es vital para la adecuada visión, y si se altera produce disfunción<br />
visual<br />
Histológicamente está <strong>com</strong>puesta de fuera a dentro por los siguientes elementos:<br />
EPITELIO CORNEAL ANTERIOR. Se trata de un epitelio escamoso estratificado<br />
(5-8 capas) de unos 50 μm de altura.<br />
Su misión es la de proteger la córnea de las partículas en suspensión que<br />
impactan con el globo ocular, y de este modo mantiene íntegra la transparencia de<br />
la córnea.<br />
Este epitelio tiene dos características principales:<br />
Está muy inervado (Rama V 1 del nervio trigémino)<br />
La estimulación de este epitelio produce el REFLEJO CORNEAL<br />
Tiene una alta capacidad proliferativa, debido a que es una barrera<br />
defensiva para el globo ocular.<br />
La descamación de las células de este epitelio provoca que proliferen a un<br />
ritmo muy elevado.<br />
Se trata <strong>com</strong>o decíamos de un epitelio estratificado no queratinizado (EPEnQ)<br />
que consta de 5 a 8 estratos unidos entre sí mediante desmosomas.<br />
Bajo este epitelio encontramos la MEMBRANA DE BOWMAN a la cual las células<br />
de este epitelio se unen mediante hemidesmosomas.<br />
66
Alberto Gómez Esteban<br />
• MEMBRANA DE BOWMAN. Se encuentra inmediatamente bajo el epitelio corneal<br />
anterior, al que se une mediante hemidesmosomas.<br />
Mide de 5 a 7 μm de espesor y está formada por colágeno tipo I.<br />
Se trata de un tejido PAS+<br />
• ESTROMA CORNEAL. El estroma o sustancia propia de la córnea es una capa<br />
colágena muy transparente que forma la capa más gruesa de la córnea.<br />
Está formada por fibras de colágeno (tipos I y V) que se disponen adoptando una<br />
morfología reticular muy resistente a la deformación y a los traumatismos.<br />
Su transparencia es esencial para la visión, así que este será el parámetro<br />
fundamental que determina la ordenación de las fibras de colágeno.<br />
La sustancia fundamental del estroma corneal es rica en proteoglucanos:<br />
Condroitín sulfato<br />
Queratán sulfato<br />
Este estrato contiene fibroblastos especiales que se denominan CORNEALOCITOS.<br />
También se encuentra inervado por fibras del V par.<br />
Se trata de un tejido avascular, y además es un órgano inmunoprivilegiado<br />
debido a que las células propias de este estrato producen factores<br />
antiangiogénicos y antimigratorios, los cuales impiden la formación de vasos y la<br />
entrada del sistema inmune.<br />
No es conveniente que el tejido inmune se introduzca en la córnea debido a que las<br />
células inmunes para avanzar lo hacen degradando la matriz mediante<br />
metaloproteasas; sabiendo que la disposición geométrica de la matriz es<br />
fundamental para su transparencia, parece lógico que su degradación y cicatrización<br />
<strong>com</strong>prometerían esta transparencia.<br />
67
Alberto Gómez Esteban<br />
• MEMBRANA DE DESCEMET. Se trata de una membrana basal enormemente<br />
gruesa (510 μm) y es producida por el endotelio corneal.<br />
Contiene fibras de colágeno tipo VI dispuestas de forma hexagonal.<br />
Tiene dos partes:<br />
Parte fibrosa. Se encuentra en la parte más anterior, y está en contacto con<br />
el estroma.<br />
Parte posterior. Es más posterior y sirve de apoyo al epitelio corneal<br />
posterior.<br />
• EPITELIO CORNEAL POSTERIOR (endotelio corneal). Se trata de un epitelio<br />
monoestratificado en contacto con el humor acuoso.<br />
La función principal de este epitelio es evitar la edematización de la córnea, y por<br />
ello su misión es evitar que entre el humor acuoso a la córnea.<br />
Contiene multitud de bombas iónicas que se encargan de regular el gradiente<br />
iónico y evitan la entrada de fluido a la córnea.<br />
68
Alberto Gómez Esteban<br />
2. Esclerótica<br />
Se trata de la capa que recubre todo el globo ocular, menos la parte más anterior, que<br />
es recubierta por la córnea.<br />
Está formada por tejido fibroso denso, con colágeno y fibras elásticas producidas por<br />
los fibroblastos de su estroma.<br />
Se trata de un tejido blanquecino debido a que sus fibras colágenas no adoptan una<br />
morfología especial <strong>com</strong>o veíamos en la córnea.<br />
Además esta capa presta inserción a los 6 músculos extrínsecos oculares.<br />
Tiene las siguientes capas, de fuera a dentro:<br />
• CAPA FIBROSA EXTERNA. Está formada por haces de colágeno entrecruzados<br />
que forman una bolsa donde se encuentra el globo ocular.<br />
Es una capa tensional, debido a que estas fibras evitan el excesivo agrandamiento<br />
del globo ocular, y lo protegen de daños.<br />
Tiene pocos vasos sanguíneos, pero muy visibles debido al contraste de color.<br />
• CAPA FINA DE MELANOCITOS. Sus fibras de colágeno están muy<br />
desordenadas, dando lugar a una capa opaca cuya finalidad es evitar el paso de<br />
luz.<br />
• CAPA CRIBOSA. Se trata del lecho por donde discurren los vasos y nervios<br />
externos del ojo.<br />
• CAPA SUPRACOROIDEA. Es la capa más profunda de la esclera.<br />
Separa la esclerótica de la coroides que se encuentra inmediatamente por debajo.<br />
69
Alberto Gómez Esteban<br />
Capa media: Úvea<br />
La capa media o ÚVEA forma la capa vascularizada y pigmentada del ojo, y se divide en<br />
tres regiones:<br />
• COROIDES<br />
• CUERPO CILIAR<br />
• IRIS<br />
1. Coroides<br />
Se trata de una porción que se extiende desde la porción posterior del globo ocular hasta<br />
la ora serrata.<br />
Se trata de una capa muy vascularizada y pigmentosa que <strong>com</strong>prende las siguientes<br />
partes:<br />
• MEMBRANA DE BRUCH. Se encuentra en contacto con el epitelio pigmentario<br />
de la retina.<br />
Se <strong>com</strong>porta <strong>com</strong>o membrana basal del epitelio fotosensible retiniano y<br />
ocasionalmente acumula depósitos de calcio y lípidos.<br />
• CAPA CORIOCAPILAR. Contiene capilares fenestrados altamente permeables<br />
cuya función es nutrir la retina en su parte externa.<br />
• CAPA VASCULAR. Contiene grandes arterias musculares y venas que luego<br />
darán lugar a los capilares de la capa coriocapilar.<br />
Estas arterias y venas están asociadas a tejido conjuntivo fibroso que se sitúa<br />
por encima del tejido de la capa supracoroidea.<br />
• CAPA SUPRACOROIDEA. Se trata de una capa asociada a melanocitos, por lo<br />
que será una especie de tejido conectivo pigmentario.<br />
Esta capa, al contener melanocitos puede desarrollar el MELANOMA OCULAR.<br />
Contribuye a la protección de la esclerótica.<br />
• ESPACIO PERICOROIDEO. Se encuentra en contacto con la esclerótica.<br />
70
Alberto Gómez Esteban<br />
2. Cuerpos ciliares<br />
Forman la porción intermedia de la úvea, y se encuentran en continuación con la<br />
coroides, que se pliega para formar estas estructuras.<br />
Consisten en un anillo fibromuscular que se encuentra entre la ora serrata y la raíz del<br />
iris.<br />
Su principal función es permitir la a<strong>com</strong>odación del CRISTALINO (proceso explicado en<br />
la unidad 12 de neurofisiología).<br />
Este anillo muscular está unido al cristalino, por lo que su contracción determina que este<br />
se ensanche o se alargue.<br />
Además es un órgano encargado de secretar HUMOR ACUOSO.<br />
Contiene dos zonas:<br />
• ZONA ANTERIOR. Se trata de una zona plegada que contiene los procesos<br />
ciliares.<br />
Los procesos ciliares son unas 70-80 evaginaciones que se encargan de sujetar el<br />
cristalino.<br />
• ZONA POSTERIOR. Es una zona lisa<br />
El EPITELIO CILIAR se trata de un epitelio biestratificado que se encuentra en la zona<br />
anterior, revistiendo los procesos ciliares. Se encarga de la secreción de humor acuoso.<br />
71
Alberto Gómez Esteban<br />
Cuenta con dos partes:<br />
El EPITELIO NO PIGMENTARIO (externo) es cúbico con microvellosidades, el<br />
cual se encarga de sostener al cristalino.<br />
Sus microvellosidades son el punto de partida de las FIBRAS ZONULARES.<br />
Estas células están unidas por uniones estrechas que lo impermeabilizan.<br />
El EPITELIO PIGMENTARIO (interno) también es cúbico y está interconectado<br />
con el epitelio externo para trasladar el exudado plasmático hacia la cámara anterior<br />
del ojo.<br />
Ambos epitelios (externo e interno) generan HUMOR ACUOSO.<br />
El HUMOR ACUOSO es esencial para la supervivencia de la córnea, debido a que su<br />
epitelio posterior está bañado por humor acuoso, y la membrana de Descemet regula el<br />
paso de nutrientes a la misma.<br />
En la zona anterior también encontraremos los MÚSCULOS DEL CUERPO CILIAR, los<br />
cuales regulan el diámetro del cristalino para permitir la a<strong>com</strong>odación de la vista.<br />
Estos músculos contienen fibras de dos tipos:<br />
MÚSCULO DE MÜLLER. Se dispone en forma de anillo circular.<br />
Cuando se contrae, disminuye el diámetro del cuerpo ciliar, y por tanto<br />
aumentará el radio del cristalino, ya que se ensancha.<br />
MÚSCULO DE BRUCH. Se dispone en forma radial.<br />
Cuando se contrae, el cristalino se alarga y por lo tanto disminuye su radio.<br />
72
Alberto Gómez Esteban<br />
Ambos músculos se encargan de regular la a<strong>com</strong>odación del ojo, y están inervados por<br />
el sistema nervioso parasimpático.<br />
El epitelio externo (no pigmentario) tiene unas fibras que forman el APARATO<br />
SUSPENSORIO DEL CRISTALINO.<br />
Esta estructura está formada por microfilamentos de 10 nm de diámetro, que se<br />
agrupan en haces de 150 unidades formando una FIBRA ZONULAR.<br />
Estas fibras tienen una doble disposición:<br />
Zona posterior (lisa). Envía fibras zonulares largas a la parte superior anterior<br />
del cristalino<br />
Zona anterior (plegada). Envía fibras zonulares cortas a la parte superior<br />
posterior del cristalino.<br />
Ambas fibras delimitan un espacio que se llama TRIANGULO DE PETIT.<br />
73
Alberto Gómez Esteban<br />
3. Iris<br />
Se localiza en la parte anterior de la úvea.<br />
Es una placa anular delgada de 12 mm de diámetro y tiene un orificio en su parte<br />
central que denominamos PUPILA y es por donde entra la luz al interior del globo ocular.<br />
Tiene dos zonas:<br />
• SUPERFICIE ANTERIOR. Es rugosa y contiene fibras de colágeno plegadas.<br />
Las células de esta superficie son fibroblastos y melanocitos.<br />
• ESTROMA DEL IRIS. Se trata de un tejido conjuntivo melánico (pigmentado).<br />
Aquí es donde tenemos la mayor concentración de melanocitos en el ojo, los<br />
cuales determinan el color del ojo.<br />
También observamos estructuras vasculares que le dan al iris una textura rugosa.<br />
• SUPERFICIE PSOTERIOR. Contiene un epitelio pigmentario cuyas células<br />
contienen melanosomas.<br />
La superficie posterior del iris se encuentra rozando con el cristalino.<br />
Los MÚSCULOS DEL IRIS se encuentran en la zona que rodea la pupila.<br />
Hay dos tipos de músculos:<br />
Músculo iridoconstrictor. Son un anillo circular de fibra muscular lisa, cuya<br />
contracción empequeñece el orificio de la pupila.<br />
Músculo iridodilatador. Son fibras radiales de fibra muscular lisa, cuya<br />
contracción agranda el orificio de la pupila.<br />
Estos músculos se encuentran regulados por el sistema nervioso vegetativo, y su<br />
función es regular la cantidad de luz que entra al ojo (unidad 12 de neurofisiología).<br />
74
Alberto Gómez Esteban<br />
Cristalino<br />
El cristalino es un órgano transparente biconvexo ubicado tras el iris cuya función es<br />
enfocar la imagen nítidamente sobre la retina.<br />
Se encuentra sujeto por las fibras zonulares, y unido mediante éstas al cuerpo ciliar,<br />
cuya contracción regula el radio del cristalino.<br />
Histológicamente distinguimos tres partes de externo a interno:<br />
• CÁPSULA ENVOLVENTE. Se trata de una especie de lámina basal que<br />
impermeabiliza la córnea.<br />
Esta lámina impide el acceso de células migratorias (leucocitos) del sistema<br />
inmune.<br />
No está vascularizada ni inervada.<br />
• EPITELIO ANTERIOR. Se trata de un epitelio cúbico que se encarga de renovar<br />
el cristalino.<br />
Las células tienen una disposición variable según la posición que adoptan dentro del<br />
cristalino y pasan de ser cúbicas a ser más alargadas (FIBRAS LENTICULARES). Estas<br />
fibras se disponen en el cristalino interno a modo de capas de cebolla.<br />
El cristalino se trata de un órgano carente de matriz extracelular, formado únicamente<br />
por células epitelioides que se renuevan muy lentamente y viven toda nuestra vida.<br />
Su transparencia se da gracias a la disposición geométrica especial de sus células, y<br />
gracias a su <strong>com</strong>pactación.<br />
Las células epiteliales del cristalino se encuentran interconectadas mediante uniones<br />
GAP, de forma que el epitelio cúbico más periférico (bañado por humor acuoso rico en<br />
nutrientes) pueden difundir nutrientes a zonas más centrales.<br />
A pesar de esta interconexión, lógicamente existirá un gradiente de concentración de<br />
nutrientes, que estarán mucho más concentrados en las zonas bañadas por humor<br />
acuoso, y menos concentrados a medida que nos internamos en el cristalino.<br />
75
Alberto Gómez Esteban<br />
Este fenómeno da lugar a la existencia de tres zonas en el cristalino:<br />
• PERIFERIA. Contiene células cúbicas con división ocasional.<br />
Se encuentra en la parte anterior del cristalino.<br />
• CORTEZA DEL CRISTALINO. Corresponde a una zona intermedia entre la<br />
periferia y el núcleo.<br />
Contiene fibras lenticulares vivas pero muy diferenciadas y quiescentes, con un<br />
aporte limitado de nutrientes.<br />
• ZONA CENTRAL. Contiene fibras lenticulares muertas debido a la insuficiencia<br />
de nutrientes, pero sus cuerpos celulares mantienen la transparencia del cristalino.<br />
76
Alberto Gómez Esteban<br />
Retina<br />
La retina es la parte nerviosa y fotosensible del globo ocular y se encarga de traducir<br />
los estímulos luminosos que recibe en señales eléctricas.<br />
Todas las estructuras anteriores iban encaminadas a enfocar la luz en esta zona, lo cual<br />
es vital para que la imagen que se forme sea nítida.<br />
Contiene dos partes principales:<br />
• ZONA ANTERIOR. Se trata de una zona no fotosensible formada por un epitelio<br />
pigmentado no sensitivo.<br />
Se superpone con la ora serrata y los cuerpos ciliares, llegando incluso a la parte<br />
posterior del iris.<br />
• ZONA POSTERIOR. Se trata de la zona fotosensible de la retina.<br />
Se extiende desde la ora serrata a la parte más posterior del globo ocular.<br />
77
Alberto Gómez Esteban<br />
Histológicamente la retina fotosensible contiene 10 capas, que enumeradas de exterior<br />
(en contacto con la coroides) a interior, son:<br />
1. EPITELIO PIGMENTARIO. Se trata de la capa más próxima a la coroides, y<br />
diferente al epitelio pigmentado no sensitivo de la zona anterior.<br />
Estas células son cúbicas y contactan con los extremos de la siguiente capa.<br />
2. CAPA DE LOS FOTORRECEPTORES. Contiene los segmentos externos de los<br />
conos y los bastones<br />
3. MEMBRANA LIMITANTE EXTERNA. Se trata de una capa en la que encontramos<br />
<strong>com</strong>plejos de unión entre dos células:<br />
Células de Müller, que son astrocitos especiales de la retina nerviosa (ver<br />
unidad 2)<br />
Fotorreceptores retinianos<br />
4. CAPA NUCLEAR EXTERNA. Contiene los núcleos alineados de las células<br />
fotorreceptoras.<br />
5. CAPA PLEXIFORME EXTERNA. Se trata de una amplia capa que contiene<br />
sinapsis de los fotorreceptores con interneuronas retinianas.<br />
Neuronas bipolares<br />
Células horizontales<br />
6. CAPA NUCLEAR INTERNA. Encontramos los núcleos de las principales<br />
interneuronas y gliocitos de la retina:<br />
Neuronas bipolares<br />
Células horizontales<br />
Células de Müller<br />
Neuronas amacrinas<br />
7. CAPA PLEXIFORME INTERNA. Se trata del lugar de sinapsis de las células<br />
bipolares con células ganglionares.<br />
Las CÉLULAS GANGLIONARES son importantes neuronas retinianas cuyos<br />
axones conforman el nervio óptico.<br />
Son las únicas células de la retina capaces de disparar un potencial de acción.<br />
8. CAPA DE LAS CÉLULAS GANGLIONARES. Contiene los núcleos de estas<br />
células.<br />
78
Alberto Gómez Esteban<br />
9. CAPA DE FIBRAS NERVIOSAS. Contiene los axones de las células<br />
ganglionares.<br />
Estos axones se dirigen a la parte posterior del ojo para formar el nervio óptico<br />
El lugar donde confluyen estos axones es la PAPILA DEL NERVIO ÓPTICO, la cual<br />
al carecer de fotorreceptores es un punto ciego fisiológico en el ojo.<br />
10. MEMBRANA LIMITANTE INTERNA. Se trata de la membrana basal de los<br />
astrocitos retinianos (células de Müller).<br />
La retina integra funcionalmente tres tipos de células:<br />
• CÉLULAS DE LA GLÍA. Son astrocitos especiales (células de Müller)<br />
• FOTORRECEPTORES. Son los conos y los bastones<br />
• NEURONAS RETINIANAS. Conducen el impulso desde los fotorreceptores:<br />
Células bipolares<br />
Células horizontales<br />
Células amacrinas<br />
Células ganglionares<br />
79
Alberto Gómez Esteban<br />
80
Alberto Gómez Esteban<br />
Tema 7. Histología del sistema auditivo<br />
Introducción<br />
El oído es el órgano de la audición y se encarga de recoger las ondas sonoras del<br />
medio externo, amplificarlas de forma conveniente, y transformarlas en un estímulo<br />
eléctrico que pueda ser interpretado por la corteza cerebral.<br />
El oído tiene tres <strong>com</strong>ponentes:<br />
• OÍDO EXTERNO. Recoge el sonido ambiental y lo conduce por el conducto<br />
auditivo hasta la membrana timpánica.<br />
Está <strong>com</strong>puesto por dos elementos:<br />
Pabellón auditivo (oreja)<br />
Conducto auditivo externo<br />
• OÍDO MEDIO. Se encarga de transformar ondas de presión sonoras en<br />
movimientos mecánicos de la membrana timpánica, que son conducidos al oído<br />
interno.<br />
Está <strong>com</strong>puesto por dos elementos:<br />
Membrana timpánica<br />
Cadena de huesecillos<br />
Además, tiene otros elementos que intervienen en las funciones intrínsecas del<br />
oído.<br />
• OÍDO INTERNO. Alberga los órganos sensitivos para la audición y el equilibrio.<br />
Recoge las vibraciones mecánicas que le transmite el oído medio, y las convierte en<br />
impulsos eléctricos.<br />
Está formado por dos elementos principalmente:<br />
Cóclea. Se trata del órgano de la audición<br />
Sistema vestibular. Se trata del órgano del equilibrio, que no participa en la<br />
audición.<br />
81
Alberto Gómez Esteban<br />
Oído externo<br />
1. Oreja<br />
La oreja o PABELLÓN AUDITIVO se trata de un órgano encargado de recoger las ondas<br />
sonoras y encauzarlas hasta el conducto auditivo externo.<br />
Contiene un eje central de cartílago elástico rodeado por piel de dos tipos:<br />
• PIEL ANTERIOR DE LA OREJA. Se trata de una estructura cutánea inmóvil,<br />
sujeta al pericondrio.<br />
Carece de estructuras pilosas.<br />
• PIEL POSTERIOR DE LA OREJA. Se trata de una piel móvil debido a que tiene<br />
más hipodermis.<br />
Esta piel sí que cuenta con estructuras pilosas.<br />
El lóbulo de la oreja se encuentra inferior y se trata de una estructura grasa altamente<br />
vascularizada.<br />
Debido a su alta vascularización enrojece cuando hay una cierta congestión<br />
cardiovascular, parámetro que es usado en clínica para detectar con antelación<br />
patologías <strong>com</strong>o el infarto de miocardio.<br />
El esqueleto cartilaginoso del oído tiene un pericondrio muy flexible rico en fibras<br />
elásticas.<br />
82
Alberto Gómez Esteban<br />
2. Conducto auditivo externo<br />
El conducto auditivo externo es una vía que se dirige desde el pabellón auricular hasta<br />
el tímpano.<br />
Se trata de un canal de 2’5 cm de longitud y un diámetro de 8 mm.<br />
Contiene dos partes:<br />
• PARTE CARTILAGINOSA. Corresponde al tercio externo del conducto auditivo.<br />
Está revestida por una epidermis que carece de papilas dérmicas, pero si tiene<br />
estructuras pilosas y glándulas sebáceas.<br />
Destacan las GLÁNDULAS CERUMINOSAS, que son de tipo túbulo-alveolar,<br />
apocrinas que secretan el cerumen.<br />
El cerumen es una secreción parduzca de naturaleza lipídica, y rica en pigmentos y<br />
moléculas insecticidas; se encarga de impermeabilizar la piel y protegerla de<br />
agentes exógenos <strong>com</strong>o los insectos.<br />
• PARTE ÓSEA. Corresponde a los dos tercios internos del conducto auditivo, y<br />
se encuentra en el espesor del hueso temporal.<br />
La piel que recubre esta porción es muy fina y carece tanto de glándulas<br />
ceruminosas <strong>com</strong>o de estructuras pilosas.<br />
Esta piel se adhiere al periostio, y se continúa hacia dentro para revestir la<br />
membrana timpánica.<br />
83
Alberto Gómez Esteban<br />
Oído medio<br />
El oído medio está formado por estructuras de origen mesodérmico, y de la cresta<br />
neural, procedentes de los arcos branquiales.<br />
Los <strong>com</strong>ponentes originados por la cresta neural y el mesodermo se unen para formar los<br />
<strong>com</strong>ponentes del oído medio, que están revestidos por un epitelio de origen<br />
endodérmico.<br />
El oído medio está formado por los siguientes <strong>com</strong>ponentes:<br />
1. Membrana timpánica<br />
Se trata de una fina membrana que separa el conducto auditivo externo de la cavidad<br />
timpánica.<br />
Tiene dos partes:<br />
• PARTE FLACCIDA. Se trata de una parte fácil de desplazar.<br />
• PARTE TENSA. Se trata de una parte más inmóvil.<br />
A su vez la membrana timpánica presenta dos caras, debido a que es una estructura que<br />
separa dos <strong>com</strong>partimentos distintos:<br />
Cara externa<br />
Se trata de la cara que mira al conducto auditivo externo.<br />
Tiene un recubrimiento cutáneo formado por piel fina que protege a la membrana<br />
timpánica.<br />
Esta piel fina se apoya sobre una dermis fina que se inserta en la propia membrana<br />
timpánica.<br />
La parte interna de la membrana timpánica es una estructura colágena fibrosa cuyas<br />
fibras se disponen en dos capas:<br />
• CAPA CIRCULAR. Sus fibras se disponen a modo de espiral.<br />
Se corresponde a la parte más interna de la membrana.<br />
• CAPA RADIAL. Sus fibras se disponen a modo de tela de araña.<br />
Se corresponde a la parte más periférica de la membrana.<br />
84
Alberto Gómez Esteban<br />
Esta estructura se <strong>com</strong>bina con cartílago hialino en su zona más externa para formar un<br />
anillo fibrocartilaginoso que proporciona soporte a la zona más central de fibras<br />
colágenas.<br />
La membrana timpánica contiene numerosas fibras elásticas que le proporcionan su<br />
capacidad de deformarse ante ondas sonoras.<br />
Además contiene numerosos capilares y fibras nerviosas, que hacen que sea una<br />
estructura enormemente sensible.<br />
Cara interna<br />
Se trata de la cara que mira hacia la cavidad timpánica.<br />
Esta cara contiene un revestimiento mucoso muy vascularizado por las ramas de la<br />
arteria del conducto auditivo y de la cavidad timpánica.<br />
La inervación de esta cara deriva del NERVIO AURICULOTEMPORAL, aunque también<br />
corre a cargo de ramas procedentes de otros pares craneales:<br />
Nervio vago (X par)<br />
Nervio glosofaríngeo (IX par)<br />
Además en esta cara se inserta el mango del MARTILLO, para continuar la transducción<br />
del movimiento vibratorio.<br />
85
Alberto Gómez Esteban<br />
2. Cavidad timpánica<br />
Se trata de un espacio irregular localizado en el seno del hueso temporal.<br />
Está delimitada por las siguientes estructuras:<br />
Lateral → Membrana timpánica<br />
Posterior → Celdillas mastoideas<br />
Anteromedial → Trompa auditiva<br />
Está revestida por una mucosa formada por un epitelio plano simple, que varía según<br />
la zona en la que nos encontremos, pudiendo ser cúbico, o pseudoestratificado.<br />
Este epitelio también varía con la edad, ya que en niños es ciliado y contiene células<br />
caliciformes, aunque estos elementos desaparecen gradualmente con la edad.<br />
El epitelio se apoya sobre una lámina propia de tejido conjuntivo laxo, muy<br />
vascularizada e inervada, lo cual es el principal factor para que las infecciones auditivas<br />
causen pronunciados edemas, y sean tan dolorosas.<br />
La cavidad timpánica está rellena de aire, y contiene en su interior la CADENA DE<br />
HUESECILLOS, que están articulados entre sí por pequeñas diartrosis para conducir el<br />
sonido.<br />
86
Alberto Gómez Esteban<br />
Los huesecillos son tres:<br />
• MARTILLO. Se encuentra unido al tímpano mediante el mango del martillo.<br />
• YUNQUE. Es intermedio y está conectado a los otros dos huesecillos.<br />
• ESTRIBO. Se encuentra unido a la membrana oval del oído interno mediante la<br />
base del estribo.<br />
Estos huesecillos se encuentran tensionados gracias a músculos presentes en la<br />
cavidad timpánica, los cuales son de tipo esquelético pero INVOLUNTARIOS, actuando<br />
únicamente de forma refleja.<br />
Estos músculos contienen un endomisio muy vascularizado e inervado, y son dos:<br />
MÚSCULO TENSOR DEL TÍMPANO. Se inserta en el mango del martillo<br />
Se encuentra inervado por el nervio trigémino (V par).<br />
MÚSCULO ESTAPEDIO. Se inserta en la cabeza del estribo.<br />
Se encuentra inervado por el nervio facial (VII par)<br />
El sistema de huesecillos se encarga de transmitir la vibración a través de la membrana<br />
timpánica hasta la membrana oval del oído interno.<br />
Estos huesecillos están interconectados entre sí y tienen un recubrimiento de<br />
periostio, que a su vez está recubierto por un epitelio cúbico simple.<br />
87
Alberto Gómez Esteban<br />
3. Trompa de Eustaquio<br />
La trompa de Eustaquio o CONDUCTO AUDITIVO INTERNO se trata de un canal de<br />
unos 4 cm de longitud que se encarga de <strong>com</strong>unicar la cavidad timpánica con la<br />
nasofaringe.<br />
Estructuralmente podemos distinguir dos partes:<br />
• PARTE ÓSEA. Representa el tercio más próximo a la cavidad timpánica.<br />
Se encuentra revestida por una mucosa que contiene epitelio cilíndrico simple<br />
ciliado.<br />
Esta porción se apoya en una lámina propia de tejido conjuntivo en la que<br />
podemos apreciar algunas glándulas mucosas.<br />
• PARTE CARGILAGINOSA. Representa los dos tercios más próximos a la<br />
faringe.<br />
Se encuentra revestida por un epitelio pseudoestratificado que contiene células<br />
caliciformes y glándulas túbulo-alveolares.<br />
Esta porción cartilaginosa a su vez se divide en dos partes:<br />
Cartílago elástico. Se continúa con la porción ósea.<br />
Cartílago hialino. Comprende la parte más próxima a la nasofaringe.<br />
En su porción más externa encontramos la AMÍGDALA TUBÁRICA que es un<br />
conglomerado de nódulos linfáticos con función defensiva.<br />
La amígdala tubárica junto con el resto de amígdalas faríngeas forman parte de un<br />
anillo linfoide llamado ANILLO DE WALDEYER, cuya función es proporcionarnos<br />
protección ante todos los patógenos ingeridos o inhalados por la faringe.<br />
La función de la TROMPA DE EUSTAQUIO es la de <strong>com</strong>unicar la cavidad timpánica<br />
con la nasofaringe para equilibrar presiones; normalmente esta estructura se encuentra<br />
cerrada, pero hay actos <strong>com</strong>o la masticación o los bostezos que provocan su apertura.<br />
88
Alberto Gómez Esteban<br />
Oído interno<br />
El oído interno es un conjunto de sacos y túbulos que están alojados en cavidades<br />
labradas en el espesor del hueso temporal.<br />
1. Laberinto óseo<br />
Está relleno de PERILINFA y está formado por las propias cavidades que encontramos en<br />
la porción petrosa del hueso temporal.<br />
Contiene dos porciones relacionadas con procesos distintos:<br />
• SISTEMA VESTIBULAR. Se trata de una cavidad ovoidea relacionada con el<br />
equilibrio y la percepción de la postura estática y dinámica.<br />
Contiene los CANALES SEMICIRCULARES que son tres túneles semiesféricos que<br />
se disponen espacialmente en tres planos perpendiculares entre sí, para abarcar<br />
180º de amplitud:<br />
Túnel superior<br />
Túnel posterior<br />
Túnel lateral<br />
Estos tres túneles se encuentran ubicados formando un ángulo de 90º entre sí,<br />
para poder abarcar movimientos provenientes de los tres ejes del espacio.<br />
Se reúnen en el VESTÍBULO, que se trata de la parte <strong>com</strong>ún que pone los tres<br />
túneles en <strong>com</strong>ún.<br />
Estos canales están revestidos por endostio y en su interior encontramos la PERILINFA.<br />
• CÓCLEA. El caracol o cóclea se trata de un canal espiral que constituye el órgano<br />
de la audición.<br />
Se trata de una estructura con unos 35 mm de longitud, y un diámetro variable de 3-<br />
9 mm.<br />
La cóclea termina en el HELICOTREMA superiormente.<br />
Tiene un eje central de hueso esponjoso que denominamos MODIOLO o<br />
COLUMELA, en cuyo interior encontramos el ganglio espiral de donde salen los<br />
nervios auditivos.<br />
89
Alberto Gómez Esteban<br />
La luz del canal está dividida por espacios, que son delimitados por membranas o<br />
láminas:<br />
LÁMINA ESPIRAL (membrana de Reissner). Divide el canal en dos<br />
porciones:<br />
Porción superior<br />
Porción inferior<br />
MEMBRANA BASILAR. Divide la porción inferior en otras dos porciones.<br />
Finalmente tenemos que estas membranas delimitan tres espacios o rampas, que<br />
de superior a inferior serán las siguientes:<br />
RAMPA VESTIBULAR (superior). Contiene perilinfa.<br />
‣ Lámina espiral (membrana de Reissner)<br />
RAMPA COCLEAR (media). Contiene endolinfa<br />
‣ Membrana basilar<br />
RAMPA TIMPÁNICA (inferior). Contiene perilinfa.<br />
El laberinto óseo de la cóclea da 2’5 vueltas en torno al MODIOLO, que es una columna<br />
ósea de 35 mm de longitud y casi 1 cm de ancho.<br />
El MODIOLO termina a nivel superior, en el punto en el que se unen la rampa vestibular<br />
y la timpánica, en lo que sería el vértice del caracol, o HELICOTREMA.<br />
90
Alberto Gómez Esteban<br />
En el modiolo encontramos también el GANGLIO ESPIRAL, que es donde se encuentran<br />
los somas de los axones que sinaptan con las células receptoras encargadas de la<br />
audición.<br />
En este punto encontramos una prominencia ósea que es el punto de inserción de las<br />
membranas cocleares:<br />
Membrana basilar<br />
Membrana de Reissner<br />
*Aclaración*<br />
La PERILINFA es el líquido que rellena el laberinto óseo.<br />
Se trata de un líquido rico en sodio (↑ Na + ) y pobre en potasio (↓ K + ).<br />
La ENDOLINFA es el líquido que rellena el laberinto membranoso.<br />
Se trata de un líquido pobre en sodio (↓ Na + ) y rico en potasio (↑ K + ).<br />
91
Alberto Gómez Esteban<br />
2. Laberinto membranoso<br />
Se trata de una estructura membranosa que sigue la morfología del laberinto óseo.<br />
La parte localizada entre el laberinto óseo, y la parte externa del laberinto membranoso<br />
está rodeada de PERILINFA.<br />
El laberinto membranoso en cambio se encuentra relleno de ENDOLINFA, y además<br />
contiene muchas proteínas que son exudados plasmáticos.<br />
Contiene <strong>com</strong>o en el caso anterior dos porciones con funcion neurosensorial.<br />
Sistema vestibular<br />
Contiene tres elementos:<br />
• CONDUCTOS SEMICIRCULARES. Contienen estructuras especializadas<br />
denominadas AMPOLLAS.<br />
Dentro de las ampollas encontramos zonas neurosensoriales que denominamos<br />
CRESTAS AMPULARES.<br />
• UTRÍCULO. La zona neurosensorial del utrículo se denomina MÁCULA<br />
UTRICULAR.<br />
• SÁCULO. También tiene una zona neurosensorial que se denomina MÁCULA<br />
SACULAR.<br />
El utrículo y el sáculo son los ÓRGANOS OTOLÍTICOS, denominados asi debido a que<br />
tienen unos pequeños cristales de carbonato cálcico denominados otolitos.<br />
92
Alberto Gómez Esteban<br />
Las CRESTAS AMPULARES junto con las MÁCULAS están formadas por epitelio<br />
pseudoestratificado ciliado de tipo neurosensorial, con dos tipos principales de células:<br />
• CÉLULAS PILOSAS TIPO I. Son células redondas con un cuello estrecho desde<br />
donde se proyectan los cilios sensoriales.<br />
Cada célula contiene 40-80 cilios, y un cilio más alto (estereocilio) que se denomina<br />
KINOCILIO.<br />
Estas células están envueltas en un cáliz, que se trata de una terminación<br />
sináptica del nervio vestibulococlear (porción vestibular); estas terminaciones<br />
forman sinapsis eléctricas y químicas con la base de la célula tipo I.<br />
• CÉLULAS PILOSAS TIPO II. Son células cilíndricas que también contienen cilios<br />
mecanorreceptores.<br />
Se diferencian de las anteriores en que las terminaciones no envuelven<br />
directamente la base de la célula, sino que se limitan a formar sinapsis con su<br />
dominio basolateral.<br />
• CÉLULAS DE SOSTÉN. Se encargan de proteger y nutrir la zona<br />
neurosensorial, debido a que a ella no llegan capilares.<br />
El fenómeno del equilibrio se da gracias a que el líquido de los conductos<br />
semicirculares, el utrículo y el sáculo (ENDOLINFA) se desplaza cuando se produce un<br />
movimiento de la cabeza.<br />
La endolinfa contacta con una membrana que flota por encima de las células<br />
neurosensoriales, causando su desplazamiento.<br />
93
Alberto Gómez Esteban<br />
Dicha membrana está formada por dos sustancias:<br />
MEMBRANA OTOLÍTICA. Se trata de un material gelatinoso rico en<br />
glucoproteínas que se encuentra en la cúpula de la cresta ampular.<br />
OTOLITOS. Son cristales de carbonato cálcico embebidos en la membrana<br />
otolítica que miden de 3-5 μm de diámetro.<br />
Cuando se desplaza la endolinfa produce un desplazamiento lateral de la membrana<br />
otolítica que se traduce en el movimiento de los cilios, lo que a su vez da lugar a la<br />
conversión de un estímulo mecánico en una señal eléctrica.<br />
94
Alberto Gómez Esteban<br />
Sistema auditivo<br />
1. Generalidades<br />
El sistema auditivo se encuentra en la CÓCLEA.<br />
Corresponde al conducto coclear (medio) en el que encontramos un epitelio<br />
neurosensible localizado en el ÓRGANO DE CORTI.<br />
El CONDUCTO COCLEAR se trata de la rampa media, delimitada por dos membranas:<br />
• Membrana de Reissner (superior)<br />
• Membrana basilar (inferior)<br />
Entre medias de estas dos membranas encontramos la MEMBRANA TECTORIA, en la<br />
que encontramos células bipolares del ganglio espiral que sinaptan con células del<br />
órgano de Corti.<br />
Se trata de un conducto relleno de ENDOLINFA.<br />
La <strong>com</strong>posición de la endolinfa se mantiene estable gracias a la ESTRÍA VASCULAR,<br />
que se trata de una region lateral del laberinto óseo, ricamente vascularizada.<br />
La estría vascular contiene las CÉLULAS DE LA ESTRÍA VASCULAR, que consisten en<br />
un epitelio pseudoestratificado, que curiosamente está vascularizado (los epitelios no<br />
suelen estarlo).<br />
La función de estas células es extraer del plasma electrolitos, que secretan a la<br />
endolinfa, y al mismo tiempo retirar sustancias de deshecho.<br />
La <strong>com</strong>posición electrolítica de la endolinfa es vital desde un punto de vista neuronal,<br />
debido a que la elevada <strong>com</strong>posición de potasio (K + ) de este fluido es responsable de la<br />
excitabilidad de las células neurosensoriales.<br />
La MEMBRANA DE REISSNER es una capa doble de células aplanadas con pequeñas<br />
diferencias estructurales entre ambas capas:<br />
‣ Las células que miran a la rampa vestibular son aplanadas con pequeñas<br />
microvellosidades.<br />
‣ Las que miran a la rampa coclear son sensibles a ondas sonoras, constituidas por<br />
firmes láminas de colágeno similares a las de la membrana timpánica.<br />
95
Alberto Gómez Esteban<br />
La LÁMINA ESPIRAL surge del modiolo, y a su vez el limbo espiral surge de la lámina<br />
espiral.<br />
El LIMBO ESPIRAL es la continuación del endostio del modiolo, que se trata del eje<br />
óseo de la cóclea.<br />
En esta lámina encontramos células que se encargan de producir la membrana tectoria,<br />
que son las CÉLULAS INTERDENTALES, encargadas de generar mucopolisacáridos y<br />
proteínas.<br />
El RODETE ESPIRAL es una prominencia localizada bajo la estría vascular, que está<br />
formada por células cúbicas de revestimiento que se continúan con células que<br />
proporcionan soporte al órgano de Corti.<br />
96
Alberto Gómez Esteban<br />
2. Órgano de Corti<br />
El órgano de Corti contiene el epitelio sensitivo de la cóclea.<br />
El epitelio sensitivo está formado por células sensitivas mecanorreceptoras ciliadas de<br />
dos tipos:<br />
• CÉLULAS PILOSAS EXTERNAS. Son las células que encontramos laterales al<br />
túnel de Corti.<br />
Estas células se disponen en tres hileras en toda la extensión de la cóclea y<br />
existirán 12.000 en cada oído.<br />
Corresponden a las fibras nerviosas de tipo II, que son minoritarias (10% de<br />
axones del nervio auditivo).<br />
• CÉLULAS PILOSAS INTERNAS. Son las células que encontramos mediales al<br />
túnel de Corti.<br />
Estas células se disponen en una hilera en toda la extensión de la coclea, y<br />
existirán unas 3500 en cada oído.<br />
Corresponden a las fibras nerviosas de tipo I, que son mayoritarias (90% de<br />
axones del nervio auditivo).<br />
Estas células en su parte inferior se encuentran fijadas a la MEMBRANA BASILAR,<br />
mientras que en su parte superior, sus cilios están fijados a la MEMBRANA TECTORIA.<br />
Cuando la membrana basilar se mueve por la llegada de un sonido, se desplazarán las<br />
células sensitivas asociadas a esta membrana, mientras que la membrana basilar quedará<br />
fija.<br />
Esto dará lugar a la despolarización de las células sensitivas debido a que el<br />
desplazamiento de la membrana basilar provoca el movimiento de los cilios hacia el<br />
estereocilio (unidad 15 de neurofisiología)<br />
Además de las células sensitivas, la cóclea contiene los siguientes elementos:<br />
• CÉLULAS PILAR. Son las células encargadas de formar el TÚNEL DE CORTI, y se<br />
denominan pilares porque tienen forma de columna.<br />
Son células de bastante altura (70 μm) que se apoyan en la membrana basilar, y<br />
las hay de dos tipos:<br />
Células pilar internas. Son mediales, más cercanas al modiolo.<br />
Células pilar externas. Son laterales, más cercanas a la estría vascular.<br />
97
Alberto Gómez Esteban<br />
• CÉLULAS FALÁNGICAS. Son células encargadas de sostener el epitelio<br />
sensorial a modo de pedestal.<br />
Hay dos tipos de célula falángica:<br />
Células falángicas externas (de Deiters). Se encargan de rodear las<br />
células sensoriales externas, es decir, las laterales al tunel de Corti.<br />
Tienen una serie de prolongaciones que forman láminas o placas<br />
cercanas a la superficie apical de las células sensoriales.<br />
Células falángicas internas. Se encargan de rodear las células<br />
sensoriales internas, es decir, las mediales al túnel de Corti.<br />
Carecen de prolongaciones, pero rodean extensamente a las células<br />
sensoriales internas.<br />
• CÉLULAS LIMITANTES. Son laterales a las células sensitivas externas, y su<br />
función es de revestimiento.<br />
Encontramos dos tipos predominantes:<br />
Células de Hensen<br />
Células de Bötcher<br />
98
Alberto Gómez Esteban<br />
99
Alberto Gómez Esteban<br />
Tema 8. Histología del cerebelo<br />
Introducción<br />
El cerebelo es un órgano situado tras el tronco del encéfalo y debajo del lóbulo occipital del<br />
cerebro.<br />
Se trata de un órgano relativamente grande (8-10 cm), que con sus 130 gramos de peso<br />
constituye el 10% del peso del sistema nervioso central.<br />
La CORTEZA CEREBELOSA es una gruesa lámina de sustancia gris localizada en la<br />
periferia del cerebelo, y es donde encontramos todas las células del cerebelo.<br />
Además de la corteza cerebelosa, también encontramos sustancia gris profunda,<br />
embebida dentro de la sustancia blanca, que corresponde a los NÚCLEOS PROFUNDOS<br />
DEL CEREBELO.<br />
El cerebelo está <strong>com</strong>puesto por LAMINILLAS CEREBELOSAS que consisten en una<br />
evaginación formada por dos <strong>com</strong>ponentes:<br />
• SUSTANCIA BLANCA. Forma el eje de la laminilla<br />
• SUSTANCIA GRIS. Recubre la sustancia blanca.<br />
Capas de la corteza cerebelosa<br />
100
Alberto Gómez Esteban<br />
1. Capa molecular<br />
La capa molecular o PLEXIFORME está situada justo por debajo de la piamadre<br />
cerebelosa.<br />
Se trata de una capa pobre en neuronas y contiene los siguientes tipos celulares:<br />
• CÉLULAS ESTRELLADAS. Son superficiales dentro de la capa molecular, y se<br />
denominan así debido a que su soma tiene forma estrellada, con un tamaño<br />
aproximado de 15-20 μm.<br />
Sus dendritas se limitan a esta capa y tienen pocas espinas.<br />
Su axón, del mismo modo se limita a esta capa y establece sinapsis inhibidoras<br />
con las células de Purkinje.<br />
• CÉLULAS EN CESTO. Se encuentran profundas dentro de la capa molecular.<br />
Sus dendritas también tienen pocas espinas.<br />
Su axón es paralelo a la superficie del cerebelo, y se ramifica sobre las células<br />
de Purkinje, con las que establece conexiones inhibitorias.<br />
Esta capa también es rica en ramificaciones dendríticas de las CÉLULAS DE<br />
PURKINJE, que ascienden hacia esta capa desde la suya propia.<br />
En esta capa encontramos axones amielínicos de CÉLULAS GRANULOSAS que<br />
discurren en forma de fibras paralelas para sinaptar con las células de Purkinje.<br />
Esta capa tiene funciones receptoras y asociativas.<br />
101
Alberto Gómez Esteban<br />
2. Capa de Purkinje<br />
Se encuentra en el límite entre la capa molecular y la granular y es la capa más<br />
estrecha del cerebelo.<br />
Aquí encontramos las CÉLULAS DE PURKINJE, con un núcleo grande en el que<br />
podemos apreciar un nucléolo evidente (núcleo en ojo de búho).<br />
Estas células son de tipo piramidal grande (neuronas gigantopiramidales) y tienen una<br />
arborización dendrítica con numerosas espinas, lo que permite que cada célula de<br />
Purkinje reciba aproximadamente 2000 sinapsis con axones de la capa molecular.<br />
Tienen un axón largo que se dirige a la sustancia blanca y finaliza en los núcleos<br />
profundos del cerebelo, o bien a núcleos fuera del cerebelo (sólo en el vestibulocerebelo).<br />
La célula de Purkinje utiliza GABA <strong>com</strong>o neurotransmisor, y por tanto es una célula<br />
inhibidora.<br />
Existen tres tipos de células de la glía asociadas a esta capa:<br />
• CÉLULAS DE FANJANA<br />
• ASTROCITOS DE BERGMANN<br />
• OLIGODENDROCITOS<br />
102
Alberto Gómez Esteban<br />
3. Capa granulosa<br />
Se trata de la capa más profunda del cerebelo, siendo limítrofe con la sustancia blanca.<br />
Contiene un alto número de células pequeñas (≈ 6 μm) que son de dos tipos:<br />
• CÉLULAS GOLGI TIPO II. Se localizan en las zonas más superiores de la capa<br />
granulosa.<br />
Son de mayor tamaño que las células granulares (35-30 μm), pero se encuentran<br />
en menor cantidad; tienen forma estrellada.<br />
Sus dendritas son largas (se dirigen a la capa molecular) menos una de ellas que<br />
se dirige al glomérulo cerebeloso para participar en sus sinapsis.<br />
Su axón es corto y se localiza en la granulosa para terminar en el glomérulo<br />
cerebeloso.<br />
La conexión de estas células es inhibidora.<br />
• CÉLULAS GRANULOSAS (granos). Son células pequeñas (6-7 μm) de soma<br />
esférico.<br />
Sus dendritas son pequeñas y se encuentran solo en la capa granulosa.<br />
Su axón es muy fino y se ramifica en forma de T en la capa molecular, siendo el<br />
origen de las FIBRAS PARALELAS que discurren paralelas a la corteza cerebelosa<br />
para sinaptar con células de Purkinje.<br />
Además de con las células de Purkinje sinaptan con las siguientes células:<br />
Células estrelladas<br />
Células en cesta<br />
La sinapsis de las células granulosas es excitatoria.<br />
103
Alberto Gómez Esteban<br />
Estas células forman el GLOMÉRULO CEREBELOSO que está formado por las siguientes<br />
estructuras:<br />
• FIBRA MUSGOSA. Las fibras musgosas son fibras que llegan al cerebelo desde<br />
procedencias externas.<br />
En la capa granulosa forman un ensanchamiento que se denomina ROSETA DE<br />
LA FIBRA MUSGOSA.<br />
La fibra musgosa forma el centro del GLOMÉRULO CEREBELOSO.<br />
• CÉLULAS GRANULOSAS. Intervienen en el glomérulo con dos estructuras:<br />
DENDRITAS DE LOS GRANOS<br />
SOMA DEL GRANO LIBRE<br />
Lógicamente debido a las partes de la célula con las que interviene, las células<br />
granulosas reciben conexiones del glomérulo, pero no influyen en él de forma<br />
directa.<br />
• NEURONA DE GOLGI TIPO II. Interviene en el glomérulo cerebeloso con las<br />
siguientes partes:<br />
AXÓN. Se trata de una conexión inhibitoria.<br />
DENDRITA<br />
Estas células reciben una conexión excitatoria con su dendrita, pero envían una<br />
conexión inhibitoria con su axón.<br />
Además asociadas a estos glomérulos cerebelosos existen ASTROCITOS EN VELO<br />
que envuelven el espacio donde se da la convergencia de axones.<br />
104
Alberto Gómez Esteban<br />
105
Alberto Gómez Esteban<br />
Tema 9. Histología del cerebro<br />
Introducción<br />
La CORTEZA CEREBRAL es la parte más externa del cerebro, <strong>com</strong>puesta por<br />
sustancia gris.<br />
Desde 1909, el anatomista Brodmann realizó una división de la corteza cerebral según su<br />
citoarquitectura en las llamadas ÁREAS DE BRODMANN.<br />
El ISOCORTEX o NEOCÓRTEX es la capa evolutivamente más nueva que consta de 6<br />
capas celulares.<br />
Las partes filogenéticamente más antiguas de la corteza son conocidas de dos formas:<br />
• PALEOCORTEX<br />
• ARQUICORTEX<br />
Estas áreas presentan una citoarquitectura primitiva con 3 capas (arquicorteza).<br />
Se vio que la corteza normalmente se dividía en 6 capas celulares claramente<br />
diferenciadas en la cual todas ellas se encuentran proporcionadas en cuanto a grosor;<br />
esto lo llamó ISOCORTEX HOMOTÍPICO.<br />
También vio que había zonas donde algunos estratos faltaban, y otras zonas donde<br />
había capas excesivamente amplias, lo que le hizo pensar que existía alguna clase de<br />
especialización.<br />
Estas cortezas las llamó ISOCORTEZAS HETEROTÍPICAS, y las hay de dos tipos:<br />
• HETEROCORTEZA GRANULAR<br />
• HETEROCORTEZA AGRANULAR<br />
También existen otros tipos de cortezas intermedias, según la clasificación de Von<br />
Economo.<br />
106
Alberto Gómez Esteban<br />
Tinciones<br />
Cuando se desea ver histológicamente la corteza cerebral, se realizan las siguientes<br />
tinciones en función de lo que deseemos ver:<br />
• TINCIÓN DE WEIGERT. Se trata de una tinción para mielina.<br />
Su función es apreciar la arquitectura microanatómica de axones, es decir, la<br />
MIELOARQUITECTURA.<br />
• AZUL DE METILENO/TOLOUDINA. Se trata de una tinción para somas<br />
neuronales.<br />
Esta tinción nos permitirá apreciar la CITOARQUITECTURA de la corteza cerebral.<br />
• TINCIONES ESPECÍFICAS VASCULARES. Se suelen corresponder a anticuerpos<br />
monoclonales.<br />
Nos permiten apreciar la ANGIOARQUITECTURA del encéfalo, es decir, sus vasos<br />
y capilares sin interferencia de tractos neuronales.<br />
Una tinción específica para células vasculares son los anticuerpos anti-CD31 que<br />
interactúan con el receptor PECAM-1.<br />
• TINCIONES ESPECÍFICAS QUÍMICAS. Se suelen corresponder también a<br />
anticuerpos monoclonales, pero en vez de ir dirigidos contra estructuras<br />
vasculares, van dirigidos contra otras sustancias.<br />
Nos permiten apreciar la QUIMIOARQUITECTURA del sistema nervioso, es decir,<br />
neurotransmisores específicos de cada zona, factores de crecimiento, etc…<br />
Una tinción específica para estudiar la quimioarquitectura son anticuerpos contra la<br />
acetilcolinesterasa, que detectaría grupos celulares colinérgicos.<br />
107
Alberto Gómez Esteban<br />
Citoarquitectura<br />
La corteza cerebral normalmente tiene 6 capas:<br />
1. CAPA MOLECULAR. Se trata de la capa más superficial, y apenas tiene células.<br />
Contiene dendritas de neuronas más profundas que reciben axones de otras<br />
cortezas, por lo que tiene FUNCIÓN ASOCIATIVA.<br />
2. CAPA GRANULAR EXTERNA. Se trata de una capa con células pequeñas.<br />
Estas células son de tipo piramidal y estrellado cuyo diámetro es de unos 10 μm,<br />
es decir, son muy pequeñas.<br />
Estas células envían dendritas ascendentes que van a la capa molecular, y por<br />
ello tienen FUNCIÓN RECEPTORA.<br />
3. CAPA PIRAMIDAL EXTERNA. Contiene neuronas pequeñas y medianas (con un<br />
diámetro máximo de 40 μm).<br />
Sus axones salen para formar una banda que se denomina ESTRÍA DE KAES-<br />
BECHTEREV cuyos axones se dirigen a núcleos subcorticales.<br />
Esta capa también tiene dendritas ascendentes que se dirigen a las capas<br />
superiores.<br />
Esta capa cortical también tiene FUNCIÓN ASOCIATIVA.<br />
4. CAPA GRANULAR INTERNA. Tiene una alta concentración de células estrelladas<br />
pequeñas.<br />
Las ramificaciones terminales horizontales de estas células suelen dirigirse al<br />
tálamo para formar la ESTRÍA EXTERNA DE BAILLANGER.<br />
Esta capa es el máximo exponente de FUNCIÓN RECEPTORA en la corteza<br />
cerebral.<br />
5. CAPA PIRAMIDAL INTERNA. Contiene neuronas piramidales de gran tamaño<br />
(pirámides gigantocelulares de Betz).<br />
Suelen encontrarse en cortezas motoras, y sus axones se agrupan en la ESTRÍA<br />
INTERNA DE BAILLANGER.<br />
Esta capa es el máximo exponente de FUNCIÓN EFECTORA en la corteza<br />
cerebral.<br />
108
Alberto Gómez Esteban<br />
6. CAPA MULTIFORME (DE MARTINOTTI). Se denomina así porque tiene células<br />
muy diferentes morfológicamente entre sí.<br />
Esta capa funcionalmente es poco relevante, pero se le atribuyen FUNCIONES<br />
MIXTAS (efectoras ¿y receptoras).<br />
Existiría por debajo una CAPA ÍNFIMA (VII), que no es considerada normalmente debido a<br />
que es inconstante.<br />
Se trataría de una membrana basal que separaría la sustancia gris presente en la capa<br />
multiforme de la sustancia blanca subyacente.<br />
109
Alberto Gómez Esteban<br />
Patrón de distribución<br />
Como <strong>com</strong>entábamos antes, la corteza evolutivamente más reciente se denomina<br />
ISOCORTEZA o NEOCORTEZA y consta de las 6 capas que acabamos de ver.<br />
Según la clasificación de Von Ecónomo existe una clasificación dentro de la corteza<br />
isocorteza.<br />
• CORTEZA HETEROTÍPICA. Contiene algunas capas desproporcionadamente<br />
grandes frente a otras que se encuentran casi ausentes.<br />
Dentro de esta clasificación existen otros dos subgéneros:<br />
CORTEZA AGRANULAR HETEROTÍPICA (I). Este tipo de corteza carece<br />
de las capas granulares (II y IV), mientras que existe un agrandamiento de<br />
las capas piramidales (III y V).<br />
Se da en cortezas efectoras.<br />
Esta distribución se observa en cortezas motoras, y también en corteza<br />
límbica.<br />
CORTEZA GRANULAR HETEROTÍPICA (V). En esa corteza las capas<br />
piramidales (III y V) se encuentran disminuidas, mientras que existe un<br />
aumento exagerado de capas granulares (II y IV).<br />
Se da en cortezas receptoras.<br />
Lo encontramos sobre todo en cortezas sensitivas (corteza somestésica,<br />
corteza auditiva…).<br />
• CORTEZA HOMOTÍPICA (II, III y IV). Sus capas celulares se encuentran<br />
proporcionadas.<br />
Se encuentra en algunas zonas de la corteza frontal, parietal y temporal (áreas<br />
psicovisuales).<br />
110
Alberto Gómez Esteban<br />
Mieloarquitectura<br />
La mieloarquitectura de la sustancia gris cerebral se trata de la distribución de fibras<br />
mielínicas que encontramos en la misma.<br />
Se realiza aplicándole a la corteza cerebral una tinción para mielina (tinción de Weigert) y<br />
nos permitirá apreciar una serie de estrías paralelas a la superficie:<br />
• LÁMINA TANGENCIAL (I). Se trata de una estría tangencial al cerebro.<br />
• ESTRÍA DE KAES-BECHTEREV (II). Se encuentra en la capa piramidal externa<br />
(III).<br />
Está formada por axones que se dirigen a núcleos subcorticales.<br />
• ESTRÍA EXTERNA DE BAILLANGER (III). Se encuentra en la capa granular<br />
interna (IV).<br />
Está formada por axones que se dirigen al tálamo.<br />
• ESTRÍA INTERNA DE BAILLANGER (IV). Se encuentra en la capa piramidal<br />
interna (V).<br />
Está formada por axones efectores.<br />
• CAPA SUBESTRIADA (V). Se encuentra en la capa multiforme (VI).<br />
• CAPA DE COLUMNAS RADIALES (VI). Se trata de fibras nerviosas que se dirigen<br />
de la corteza a la sustancia blanca cerebral.<br />
La capa de las columnas radiales es perpendicular a la superficie del encéfalo.<br />
111
Alberto Gómez Esteban<br />
Módulos corticales<br />
Si observamos el conjunto de citoarquitectura y axones, vemos que en determinadas<br />
zonas del cerebro existe un patrón repetitivo que da lugar a COLUMNAS CORTICALES.<br />
Las columnas corticales son asociaciones de neuronas que forman columnas<br />
verticales, interconectadas entre sí en una misma área.<br />
Estas columnas corticales son la unidad funcional del cerebro.<br />
Los circuitos que forman las columnas se repiten infinidad de veces para dar lugar a<br />
estructuras que explican hasta cierto punto los fenómenos cognitivos superiores.<br />
Las columnas miden ≈ 300 μm, de modo que en 1 mm caben aproximadamente 3<br />
columnas.<br />
Existen conexiones asociativas entre neuronas de columnas corticales adyacentes, de<br />
modo que cada columna además de ser una entidad propia, también constituye unidades<br />
funcionales con las columnas de al lado.<br />
Esto permite hacernos una idea del nivel de <strong>com</strong>plejidad del cerebro.<br />
Angioarquitectura<br />
El encéfalo, y el cerebro más concretamente es una estructura que restringe mucho el<br />
paso de moléculas a su medio interno.<br />
Esto está permitido gracias a la BARRERA HEMATOENCEFÁLICA que está formada a su<br />
vez por tres tipos de barreras (unidad 2):<br />
• ENDOTELIO. Se trata del epitelio plano simple que encontramos revistiendo<br />
internamente los vasos sanguíneos.<br />
El endotelio encefálico es continuo pero contiene numerosos receptores para la<br />
transcitosis.<br />
• MEMBRANA BASAL. Se encuentra envolviendo al endotelio y a ella se anclan<br />
los pies perivasculares de los astrocitos.<br />
112
Alberto Gómez Esteban<br />
• MEMBRANA GLIAL LIMITANTE PERIVASCULAR. Está formada por las<br />
prolongaciones podocíticas de los astrocitos.<br />
Los astrocitos presentan uniones estrechas entre sí para impermeabilizar<br />
máximamente este espacio, de forma que todo lo que deba pasar al medio interno<br />
perineuronal pase antes por el citoplasma del astrocito.<br />
Esto regula el acceso de iones y metabolitos al espacio perineuronal.<br />
• MEMBRANA GLIAL LIMITANTE SUPERFICIAL. Rodea a la piamadre por debajo.<br />
La integridad de la barrera hematoencefálica está formada sobre todo por las uniones<br />
estrechas del endotelio, las cuales si se resienten darán lugar a la acumulación de líquido<br />
en el parénquima cerebral, es decir, un EDEMA CEREBRAL.<br />
Las uniones estrechas normalmente regulan el acceso de líquido al espacio<br />
perineuronal, y si degeneran, la entrada aumentada de plasma, junto al hecho de que<br />
en el encéfalo no existen vasos linfáticos, causaría el edema.<br />
La membrana hematoencefálica es fundamental para la salud cerebral, pero en situaciones<br />
patológicas puede ser un estorbo; este es el caso del GLIOMA CEREBRAL.<br />
El GLIOMA CEREBRAL es un tumor de las células gliales, y tiene un tratamiento<br />
dificultoso debido a que la quimioterapia no puede pasar a través de la barrera<br />
hematoencefálica.<br />
Además existe la reacción de gliosis cicatricial en torno al tumor, lo cual impide aun más<br />
el acceso de fármacos.<br />
Los capilares cerebrales forman una red anastomótica que rodea las columnas y envía<br />
colaterales al interior de dichas columnas.<br />
113
Alberto Gómez Esteban<br />
Estos apuntes fueron realizados por Alberto Gómez Esteban<br />
Apuntes de la asignatura de Neurociencia<br />
Impartida por el Dr. Fernando Vidal Vanaclocha el curso del 2012<br />
2º de Medicina.<br />
114