ORGANOGRAFIA MICROSCOPICA HUMANA - VeoApuntes.com
ORGANOGRAFIA MICROSCOPICA HUMANA - VeoApuntes.com
ORGANOGRAFIA MICROSCOPICA HUMANA - VeoApuntes.com

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
Página1<br />
MEDICINA<br />
ORGANOGRAFÍA<br />
MICROSCÓPICA<br />
<strong>HUMANA</strong>
M<br />
Página2
Página3<br />
ÍNDICE<br />
INTRODUCCIÓN…………………………………………………………..………………5<br />
SISTEMA CARDIOVASCULAR……………………………………………….….7-18<br />
Corazón……………………………………………………………………………………………………………………………………..……....7-8<br />
Sistema Vascular………………………………………………………………………………………………...…………………………....9-18<br />
Generalidades…………………………………………………………………………………………...…………………………...9<br />
Arterias………………………………………………………………………………………….……………………………....10-12<br />
Capilares…………………………………………………………………………………………..……………....…………....12-13<br />
Venas…………………………………………………………………………………………………………………..………...13-16<br />
Sistema Linfático……………………………………………………………………………………….…………..……....16-17<br />
Estructuras Vasculares Especiales…………………………………………………………………………….……17-18<br />
SISTEMA RESPIRATORIO…………………………………………………...…19-32<br />
Cavidades Nasales y Senos Nasales…………………………………………………………………………………...….…………19-22<br />
Faringe………………………………………………………………………………………………………………………..………………...……22<br />
Laringe………………………………………………………………………………………………………………..…………………………23-24<br />
Tráquea………………………………………………………………………………………………………………..………………………..25-26<br />
Bronquios…………………………………………………………………………………………………………………….……………………..27<br />
Bronquiolos……………………………………………………………………………………………………….……….……………….…28-29<br />
Alvéolos……………………………………………………………………………………………………………….………..…………..…..30-32<br />
Pleura………………………………………………………………………………………………………………………..…..…………………...32<br />
APARATO DIGESTIVO…………………………………………………..…….…33-68<br />
Cavidad Oral………………………………………………………………………………………………...……………………..…..……..33-34<br />
Lengua…..………………………………………………………………………………………………………………………..…..…………35-36<br />
Diente…………………………………………………………………………………………………………………………..…..……………37-39<br />
Glándulas Salivales…………………………………………………………………………………………………………………………39-41<br />
Faringe…………………………………………………………………………………………………………………………..…..……………….42<br />
Generalidades del Tubo Digestivo……………………………………………………………………………………………...…...42-44<br />
Esófago……………………………………………………………………………………………………………..…..………………….……45-46<br />
Estómago……………………………………………………………………………………………………………..…..……………………47-51<br />
Intestino Delgado…………………………………………………………………………………………………………..…..………..…52-56<br />
Intestino Grueso, Recto y Conducto Anal………………………………………………………………………………...………57-60<br />
Hígado……………………………………………………………………………………………………………..…..……………………..…61-67<br />
Vesícula Biliar……………………………………………………………………………………………………………..…..……….....………68<br />
Páncreas Exocrino……………………………………………………………………………………………………………..…......……69-70<br />
SISTEMA LINFÁTICO………………………………………………………..…71-86<br />
Células Linfoides……………………………………………………………………………………………………..…..………....………71-73<br />
Vasos Linfáticos….………………………………………………………………………………..…..……….....……..……………………….74<br />
Tejido Linfoide Difuso, Nódulos Linfáticos y Amígdalas….….…………………………………………………………….74-76<br />
Ganglios Linfáticos….….………………………………………………………………………………..…..……….....………………....77-80<br />
Timo….….………………………………………………………………………………..…..……….....……………………………………...80-83<br />
Bazo….………………………………………………………………………………..…..……….....……………………………………...…..84-86<br />
Médula Ósea….………………………………………………………………………………..…..……….....…………………………….….….86
Página4<br />
SISTEMA TEGUMENTARIO…………………………………………………87-100<br />
Epidermis………………………………………………………………………………………………………………………………………87-92<br />
Células de la Epidermis…………………………………..…………………………………………………………………….88-92<br />
Dermis…………………………………………………………………………………………………………………………………………..93-94<br />
Pelo…………………………………………………………………………………………………………………………………………...…..95-97<br />
Glándulas Sebáceas………………………………………………………………………………………………………………………..……98<br />
Glándulas Sudoríparas…..………………………………………………………………………………………………………...……98-100<br />
Uñas………………………………………………………………………………………………………………………………………………....100<br />
SISTEMA ENDOCRINO………………………………………………………101-123<br />
Hipófisis……………………………………………………………………………………………………………………….…………....101-109<br />
Glándula Pineal o Epífisis…………………………………………………………………………………………………...……….109-110<br />
Glándula Tiroides……………………………………………………………………………………………………………………….111-113<br />
Glándulas Paratiroides……………………………………………………………………………………………………….………114-115<br />
Glándulas Suprarrenales………………………………………………………………………………………………………...…..116-120<br />
Páncreas Endocrino……………………………………………………………………………………………………………………121-123<br />
APARATO REPRODUCTOR FEMENINO………………………...……125-150<br />
Genitales Externos………………………………………………………………………………………………...……………………126-127<br />
Ovario………………………………………………………………………………………………………………..………………………127-133<br />
Oviducto o Trompas de Falopio………………………………………………………………………………..…………………134-135<br />
Útero…………………………………………………………………………………………………………………………………………135-139<br />
Vagina…………………………………………………………………………………………………………………….………….………139-140<br />
Placenta……………………………………………………………………………………………………………………..………………140-144<br />
Mama……………………………………………………………………………………………………………………………………...…144-150<br />
APARATO REPRODUCTOR MASCULINO………………………..…151-167<br />
Escroto y Testículos…………………………………………………………….……………………………………………………..151-159<br />
Espermatogénesis…………………………………..…………………………………………………..……………………155-157<br />
Vías Espermáticas………………………………………….…………………………………………………………………………..159-162<br />
Pene……………………………………………….………………………………………………………………………………...………..163-164<br />
Vesículas Seminales…………………………………………………………………..………………………………………………..164-165<br />
Prostáta…………………………………………………………………………………………………………………………….……….165-166<br />
Glándulas de Cowper……………………………………………………………………………………………………...………………….166<br />
Semen…………………………………………………………………………………………………………………………………….…………167<br />
SISTEMA URINARIO……………………………………………...…………169-182<br />
Riñón…………………………………………………………………………………………………………………………………………169-180<br />
Uréter…………………………………………………………………………………………………………………………..…………………...180<br />
Vejiga………………………………………………………………………………………………………………...………………………180-181<br />
Uretra…………………………………………………………………………………………………………………..……………………181-182<br />
ANEXO.……………………………………………...………………………….…183-193<br />
Exámenes USP-CEU……………………………………………………………………………………………………………….……183-193<br />
Estas páginas recogen una visión detenida de los distintos tejidos que <strong>com</strong>ponen el cuerpo humano. La información<br />
ha sido obtenida del libro ‘Histología’ de Ross y Pawlina (4ª y 5ª Ed.) y de apuntes de la Universidad Complutense de<br />
Madrid (www.veoapuntes.<strong>com</strong>) <strong>com</strong>pletados con la teoría del profesor Miguel Ángel Sáez García de la Universidad<br />
San Pablo CEU de Madrid obtenidos de sus clases, diapositivas y apuntes de Laura del Olmo.<br />
Miguel Argüello de Tomás
Página5<br />
INTRODUCCIÓN<br />
La Organografía estudia el ensamblaje de las estructuras de orden superior que traducen la integración<br />
morfológica funcional de las partes que <strong>com</strong>ponen al organismo.<br />
Existe una serie de conceptos que permiten estudiar los diversos tejidos del organismo:<br />
ÓRGANO HUECO: órgano con una luz tapizada de epitelio; fuera de este se encuentran otros tejidos <strong>com</strong>o<br />
cartílago, hueso, músculo o tejido conjuntivo.<br />
ÓRGANO MACIZO: carece de luz y sus células se disponen en grupos, cordones, nidos… Tienen una capa de<br />
tejido conectivo que emite tabiques al interior dando consistencia al órgano.<br />
PARÉNQUIMA: células funcionales del órgano, es decir, aquellas células que desarrollan la función<br />
características del órgano.<br />
ESTROMA: células que dan soporte físico o metabólico al parénquima.<br />
La mayoría de los órganos huecos tienen en su primera capa el parénquima y el resto suele ser estroma,<br />
aunque tenga su propia función (no significa que sean células estáticas; por ejemplo, pueden aportar<br />
nutrientes o contraerse).<br />
En los órganos macizos la cápsula externa <strong>com</strong>pone el estroma y el parénquima está en el interior.<br />
Los tumores se distribuyen por el parénquima y por el estroma para conseguir el aporte necesario.<br />
MUCOSA ¡ !<br />
Una mucosa es la capa interna de un órgano hueco; es un epitelio cuya lámina basal se encuentra apoyada<br />
en tejido conjuntivo laxo.<br />
Hay tantas mucosas en el organismo <strong>com</strong>o órganos huecos.
Página6
Página7<br />
SISTEMA CARDIOVASCULAR<br />
El sistema cardiovascular transporta sangre y linfa hacia los tejidos del organismo y desde allí de retorno al corazón.<br />
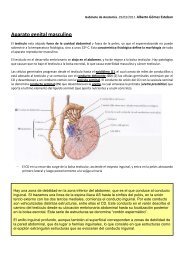
CORAZÓN<br />
El corazón es un órgano hueco que tiene 4 cavidades (dos Au y dos V) a través de los cuales bombea sangre<br />
(600L/día) desde las venas a las arterias.<br />
Se encuentra formado por 3 tejidos organizados en 3 capas, que, de dentro a fuera, son: endocardio, miocardio y<br />
pericardio.<br />
Pesa 250-300g en mujeres y 300-359g en hombres y su espesor varía según la cavidad: el VD tiene 0,3-0,5cm de<br />
espesor y el VI uno más (1,3-1,5cm).<br />
Endocardio<br />
Está formado por un epitelio simple plano (endotelio) asentado en una membrana basal apoyada en un tejido<br />
conjuntivo laxo; es AVASCULAR.<br />
Tejido Subendocárdico<br />
Es el endocardio presente en los sistemas de conducción y está VASCULARIZADO.<br />
Miocardio<br />
Presenta mayoritariamente miocitos, pero también se encuentran fibroblastos y otras células propias del tejido<br />
conjuntivo. Es más grueso en el V que en la Au para dar una gran P a la sangre a su salida a las arterias.<br />
El miocito es una célula fusiforme ligeramente estrellada que forma una red tridimensional y se une a otros miocitos<br />
por uniones tipo GAP llamadas uniones ocluyentes para formar un sincitio funcional y asegurar la contracción<br />
conjunta de todos los miocitos. La contracción se da gracias a las P contráctiles de su citoplasma y al<br />
R.Sarcoplásmico, que asegura la concentración de Ca.<br />
Miocitos de las orejuelas<br />
Secretan la hormona natriurética auricular, que disminuye la P sanguínea, aumenta la secreción de agua, Na y K y<br />
disminuye la secreción de renina y aldosterona (por lo que disminuye el volumen sanguíneo).<br />
Pericardio<br />
Es la capa más externa del corazón y se pueden distinguir 2 capas:<br />
<br />
<br />
Visceral o Epicardio: está formado por un mesotelio apoyado en una membrana basal apoyada en un tejido<br />
conjuntivo laxo. Presenta pequeños grupos de capilares y de adipocitos que discurren por las arterias<br />
coronarias, así <strong>com</strong>o fibras nerviosas y vasos linfáticos.<br />
Parietal: es un mesotelio más gruesa por un mayor porcentaje de fibras de colágeno y elásticas.<br />
Entre las dos capas queda un espacio virtual de unos 50cm 3 que se conoce <strong>com</strong>o cámara cardíaca.<br />
Contiene una película de líquido que disminuye el rozamiento e impide que se separen las dos hojas<br />
pericárdicas.
Página8<br />
La pericarditis conlleva el aumento de volumen de esta cavidad virtual.<br />
Esqueleto fibroso<br />
Está formado por tejido conjuntivo denso y permite el anclaje del corazón. Consta de:<br />
- Trígono fibroso: de él salen los anillos valvulares de las 4 válvulas<br />
- Anillos valvulares<br />
- Septo membranoso: tercio superior del tabique interventricular<br />
Músculos papilares y Válvulas<br />
Los músculos papilares están formados por endocardio y miocardio; es VASCULAR.<br />
De su extremo parten proyecciones de endocardio con tejido conjuntivo denso que forman las cuerdas tendinosas;<br />
las valvas están formadas por endotelio con tejido conjuntivo laxo; son AVASCULARES.<br />
Nódulo de Arancio: borde muy fibroso de cada una de las valvas en el punto donde se juntan.<br />
Sistema de conducción<br />
Las células del sistema de conducción son miocitos transformados que poseen menos P contráctiles (y en<br />
consecuencia menor estriación) y menos uniones oclusivas. En contra, son más grandes que el resto de miocitos.<br />
Está formado por el Nódulo S-A, el Nódulo Au-V y el Haz de Hiss (derecho e izquierdo).<br />
Vascularización e Inervación<br />
Las arterias derecha e izquierda (descendente A y circunfleja) llevan sangre al corazón, desde donde drena al seno<br />
coronario. También llegan vasos linfáticos y está inervado por los nervios vagos, esplácnico y terminaciones<br />
sensitivas.
Página9<br />
SISTEMA VASCULAR<br />
El Sistema Vascular permite distribuir, <strong>com</strong>unicar e integrar funcionalmente al cuerpo. La distribución de<br />
nutrientes y recogida de desechos es imprescindible. Es un circuito cerrado integrado por 2 subcircuitos: el<br />
periférico o sistémico y el pulmonar.<br />
Los vasos que lo integran se encuentran influidos en su estructura histológica según factores mecánicos (<strong>com</strong>o<br />
la P, que determina la rigidez) y factores metabólicos (permeabilidad).<br />
En general, una estructura grande tiene un diámetro>100μm y es independiente del medio en el que se<br />
encuentre; si su diámetro es menor es microcirculación y depende del medio.<br />
Existen 3 tejidos básicos en el sistema vascular:<br />
Tejido Endotelial<br />
Es un tejido plano simple que forma la capa de revestimiento interno de los vasos, Es de origen mesenquimal y<br />
procede de los angioblastos.<br />
Según el tipo de vaso y su localización presentan unas u otras características visiblemente diferenciadas según<br />
qué función predomine de:<br />
o Coagulación sanguínea<br />
o Intercambio molecular entre tejidos y sangre<br />
o Receptores hormonales y de otras moléculas<br />
o Procesado antigénico y presentación antigénica<br />
Tejido Conectivo<br />
Se encuentra en las 3 capas generales de los vasos aportando soporte a cada una de ellas.<br />
Capas o Túnicas<br />
CAPA ÍNTIMA: endotelio + membrana basal + tejido conjuntivo laxo/subendotelial<br />
CAPA MEDIA: músculo liso + tejido conectivo con fibras reticulares/elásticas según el vaso.<br />
CAPA ADVENTICIA: tejido conectivo laxo + anejos; dentro de estos anejos puede haber capilares sanguíneos<br />
que se conocen <strong>com</strong>o “vasa vasorum”, fibras nerviosas conocidas <strong>com</strong>o “nervi vascularis” (cambia nombre<br />
según libro) y/o vasos linfáticos.<br />
Tejido Muscular Liso<br />
Consta de células musculares sin estriación cuya contracción mantiene la P homeostática del sistema vascular. Se<br />
dispone únicamente en la capa media (con excepciones) entremezclándose con las fibras y recibe inervación del SN<br />
Vegetativo, que llega a través de la adventicia a la media.
Página10<br />
ARTERIAS<br />
Arterias Elásticas o de Conducción<br />
Las arterias elásticas son de conducción y tienen un diámetro aproximado de 1cm. Dentro de este grupo se<br />
engloban a las arterias aorta, ilíaca <strong>com</strong>ún o la subclavia.<br />
CAPA ÍNTIMA<br />
Tiene un endotelio grueso cuyas células están estrechamente unidas por zonulae occludens para evitar el paso<br />
de sangre al interior del tejido; también hay uniones tipo GAP Junctions.<br />
Presentan muchas vesículas de transcitosis (generalmente entran en la célula por pinocitosis) y se pueden<br />
percibir en sus citoplasmas unas inclusiones bastiniformes llamadas Cuerpos de Weibel-Palade, unas vesículas<br />
electrodensas que contienen el factor VIII de la coagulación<br />
Se encuentran apoyadas en una lámina basal fina apoyada en tejido conectivo subendotelial que puede<br />
presentar colágeno. En las arterias elásticas de gran calibre aparecen fibras de colágeno y elásticas que son<br />
sintetizadas por fibras musculares lisas (también presentes en esta capa) que permiten una mayor contracción<br />
de la arteria).<br />
Aunque no se percibe muy bien, existe una Lámina Elástica Interna (LEI).<br />
CAPA MEDIA<br />
Se dispone en unas 50-70 láminas de fibras elásticas cuyo diámetro es de hasta 3μm. Estas fibras son<br />
sintetizadas por las células musculares lisas (NO hay fibroblastos en esta capa), que se anclan a las fibras para<br />
producir la contracción. En total, el grosor de esta capa puede ser de hasta 2mm. Además, estas laminillas<br />
tienen fenestraciones que permiten el paso de sustancias a su través.<br />
También aparece colágeno y sustancia fundamental.<br />
En personas con HTA (HiperTensión Arterial) hay un aumento en la cantidad y espesor de las lamnillas elásticas.<br />
Si muta algún gen de las fibras (<strong>com</strong>o la fimbrina I) puede aumentar el riesgo de aneurismo (dilatación arterial<br />
que si revienta produce la muerte instantánea). De todos modos, la principal causa de aneurisma es la úlcera<br />
resultante del desprendimiento de una placa de ateroma.<br />
CAPA ADVENTICIA<br />
Está formada por tejido conjuntivo laxo con fibras colágenas y elásticas poco organizadas con “vasa vasorum”,<br />
“nervi vascularis”, vasos linfáticos, fibroblastos y macrófagos.<br />
La arteria se encuentra nutrida sus dos tercios internos por difusión de la sangre que circula a través de ella<br />
(vesículas pinocíticas), y el tercio externo por los vasa vasorum.<br />
Arterias Musculares o de Distribución<br />
Presentan un predominio del tejido muscular sobre el resto, <strong>com</strong>o en las arterias braquiales, femorales, radiales<br />
y poplíteas. Uno de los rasgos característicos que las diferencias de las elásticas es que presentan una LEI y una<br />
LEE (Lámina Elástica Externa) muy prominentes.<br />
Según su diámetro hay dos subtipos:
Página11<br />
Arteria Muscular Grande<br />
Tienen un diámetro de entre 2 y 10mm.<br />
Su capa íntima está formada por células endoteliales fuertemente unidas con una membrana basal apoyada en<br />
un tejido conjuntivo laxo con fibras elásticas y de colágeno. Acaba en la LEI.<br />
La capa media consta de 10 a 60 láminas de músculo liso dispuestas helicoidalmente en torno al eje de la<br />
arteria, con lo que consiguen una contracción más efectiva. Esta contracción depende de los canales de Ca,<br />
controlados por estímulos electromecánicos o adrenérgicos que se convierten en el blanco del tratamiento de la<br />
HTA.<br />
Termina en una lámina elástica externa o LEE.<br />
La capa adventicia es gruesa y presenta “vasa vasorum” y nervios vegetativos. También se pueden encontrar<br />
fibras colágenas, elásticas, macrófagos y adipocitos dispersos.<br />
Arteria muscular pequeña<br />
Tienen un diámetro de 0,1 a 2mm. El endotelio descansa directamente sobre la LEI (no suele existir tejido<br />
subendotelial), la capa media tiene 3-4 capas celulares de músculo liso y contacta con la adventicia (no hay<br />
LEE), que carece de vasa vasorum.<br />
Arterias Mixtas o de Transición<br />
Como las arterias carótidas externa e interna, la axilar, las ilíacas externa e interna y las viscerales. Presentan<br />
islotes de fibras musculares que inerrumpen las láminas elásticas y forman 2 capas: capa muscular interna y<br />
elástica externa.<br />
La capa media tiene un grosor variable y adaptable a la P sanguínea y a la fuerza externa.<br />
Arterias Especializadas<br />
<br />
<br />
<br />
<br />
<br />
<br />
Arteria Coronaria<br />
Arterias cerebrales y durales<br />
Arterias de flexión: poplítea y axilar<br />
Arterias del MI: son más gruesas que las de igual calibre<br />
Arteria Pulmonar: tienen paredes finas y distensibles; de hecho, histológicamente son más parecidas a<br />
una vena normal de la circulación sistémica que a una arteria <strong>com</strong>ún.<br />
Arteria Umbilical: su capa íntima tiene sólo endotelio, carece de LEI, su capa media tiene pocas fibras<br />
elásticas con 2 capas musculares. Su capa adventicia se encuentra en contacto con la gelatina de<br />
Wharton, un tejido conjuntivo que rodea a las arterias umbilicales y a la vena umbilical, que con el<br />
tiempo se transforma en fibroblastos maduros.<br />
Arteriolas<br />
Son microcirculación porque su diámetro está entre 15 y 100μm. Su función está relacionada con la regulación<br />
del flujo sanguíneo a los tejidos. En su capa más interna hay un revestimiento endotelial sin tejido subendotelial<br />
y la LEI sólo aparece cuando el diámetro es entre 50 y 100μm.<br />
Las uniones mioendoteliales son uniones entre las células endoteliales y células musculares lisas para la<br />
transmisión de información de señales hormonales o bioquímicas recibidas por el endotelio a la fibra muscular<br />
lisa, que es la efectora.<br />
Las uniones mioendoteliales son GAP Junctions cuyos canales están formados por hexámeros de conexina que<br />
en conjunto forman el conexón. A través de ellos viajan iones o AMPc para regular la contracción muscular.
Página12<br />
Metarteriolas<br />
Son vasos que <strong>com</strong>parten características con la arteriola y el capilar. Regulan a modo de esfínter el flujo<br />
sanguíneo por regulación del SN Vegetativo. Así se puede permitir un aumento del flujo con un diámetro grande<br />
o disminuirlo contrayendo el músculo liso de las paredes vasculares.<br />
CAPILARES<br />
Los capilares se organizan en redes tridimensionales anastomosadas que aseguran el riego sanguíneo evitando<br />
la necrosis por falta de sangre.<br />
El diámetro total del lecho capilar es 800 veces el de la aorta, aunque su velocidad es muy inferior: en los<br />
capilares la velocidad es de 0,3cm/seg y en la aorta es de 32cm/seg.<br />
Las células endoteliales realizan diversas funciones:<br />
Eliminación de desechos y aportan nutrientes<br />
Mantienen la membrana basal con síntesis de colágeno y proteoglucanos<br />
Intervienen en la coagulación:<br />
‣ Favoreciéndola: Factor VIII<br />
‣ Dificultándola:<br />
• Prostaciclina<br />
• Trombomodulina<br />
• Óxido Nitroso (NO); inhibe la adherencia y la agregación plaquetaria<br />
Control del flujo sanguíneo<br />
‣ Acción vasoactiva<br />
• NO<br />
• Prostaglandinas y prostaciclinas, <strong>com</strong>o el TxA 2<br />
• Serotonina o 5-HT, bradiquinina (BK) y endotelina, <strong>com</strong>o ET-1<br />
• Angiotensina I (AI) y Angiotensina II (AII).<br />
Intervienen en la reacción inflamatoria agua<br />
‣ Interleucinas 1, 6 y 8<br />
‣ Moléculas de adherencia celular<br />
Producen algunos factores de crecimiento<br />
‣ Factor de Crecimiento de Fibroblastos<br />
‣ Factor de crecimiento derivado de las plaquetas<br />
‣ Factor estimulante de colonias de células sanguíneas<br />
Intervienen en el metabolismo de lipoproteínas
Página13<br />
Tipos<br />
TIPOS ENDOTELIO LÁMINA<br />
PROPIA<br />
Somáticos o<br />
Continuos Continuo; necesita mediadores para el<br />
transporte transendotelial<br />
Continua<br />
Fenestrados<br />
Sinusoides o<br />
Discontinuos<br />
Con poros de 60-80nm separados 50nm;<br />
permiten un rápido intercambio<br />
Continua<br />
LOCALIZACIÓN<br />
Tejidos muscular, conectivo, nervioso,<br />
adiposo, pulmones, riñones (en vasos<br />
rectos), canales de Havers del hueso,<br />
glándulas exocrinas, testículos, cerebro y<br />
barreras hematoencefálica/testicular/tímica<br />
Con diafragma (8 fibrrillar radiales):<br />
Glándulas endocrinas e intestino<br />
Sin diafragma:<br />
Glomérulo Renal<br />
Fenestrado Fenestrada Órganos linfáticos, pars distalis de<br />
adenohipófisis e hígado<br />
Los sinusoides son conductos tortuosos con un diámetro de entre 30 y 40μm que suelen estar asociados a<br />
macrófagos, <strong>com</strong>o las células de Kupffer en el hígado o las células reticulares en los órganos linfáticos.<br />
Pericitos<br />
Aparecen en los capilares continuos y en las vénulas, <strong>com</strong>partiendo lámina basal con las células endoteliales. Su<br />
principal función es la contracción, por lo que presentan P contráctiles (tropomiosina, isomiosina, actina y miosina);<br />
incluso puede diferenciarse a una célula muscular lisa.<br />
Presentan prolongaciones primarias y secundarias (uniones <strong>com</strong>unicantes).<br />
Además, suelen tener receptores H2 de la histamina.<br />
Los tumores que se desarrollan por la proliferación de pericitos se denominan hemangiopericitomas, y pueden ser<br />
benignos o malignos.<br />
VENAS<br />
Las venas no tienen unas túnicas tan bien definidas <strong>com</strong>o en las arterias.<br />
Estos vasos se pueden dividir en 4 subtipos generales en función de su tamaño y características. Por lo general, las<br />
venas de gran y mediano calibre transcurren junto a las arterias del mismo nombre y suelen aparecer en los cortes<br />
histológicos con luces más grandes y paredes más finas que las de las arterias que a<strong>com</strong>pañan.<br />
Según el autor existen 3 o 4 grupos, según se consideren las vénulas y las venas de pequeño calibre en una misma<br />
categoría o en dos distintas. Así, son:<br />
Vénulas (Pericíticas) o Vénulas Postcapilares<br />
Son vasos que se constituyen por la fusión de varios capilares. Tienen una longitud de unos 700μm y un diámetro de<br />
15 a 20μm.<br />
Tienen una túnica íntima formada por un endotelio que <strong>com</strong>parte lámina basal con pericitos que, al tener receptor<br />
H2, son sensibles a la acción de la histamina; también responden a la serotonina. Así, la vénula puede variar su<br />
diámetro y además permite una rápida marginación y diapédesis de leucocitos.<br />
En la capa media hay pericitos y en la adventicia hay fibras colágenas, elásticas y fibroblastos.
Página14<br />
Venas de Pequeño Calibre<br />
En general, tienen un diámetro de 0,1-1mm y están formadas por las tres túnicas vasculares:<br />
Túnica Íntima: endotelio y tejido subendotelial con fibras reticulares; tienen receptores para algunas células,<br />
<strong>com</strong>o los linfocitos por receptores específicos.<br />
Túnica Media: formado por 1-3 estratos de células musculares<br />
Túnica Adventicia: formada por fibroblastos entre fibras colágenas y elásticas.<br />
No hay LEI ni LEE.<br />
Las venas de pequeño calibre incluyen a las Vénulas Musculares y a las Vénulas de Endotelio Alto (VEA).<br />
VÉNULAS MUSCULARES: están situadas más allá de las vénulas postcapilares en la red venosa de retorno y se<br />
diferencian de las anteriores en que tienen ya generalmente 2 capas de fibras musculares lisas y una fina capa<br />
adventicia.<br />
VEA: se encuentran los órganos linfoides y se caracterizan porque tienen un endotelio cilíndrico o cúbico de gran<br />
altura. De este modo permiten el paso de linfocitos por diapédesis pero impiden el paso de agua hacia el tejido en el<br />
que sen encuentre el vaso. Así, permiten que los linfocitos B y T abandonen el torrente sanguíneo hasta el órgano,<br />
<strong>com</strong>o por ejemplo un ganglio linfático, y desarrollen allí su acción (en este caso, madurar a plasmoblastos y<br />
linfoblastos).<br />
Venas de Mediano Calibre<br />
Las venas de mediano calibre presentan 3 túnicas más diferenciadas que en las anteriores, de diámetros menores.<br />
Dentro de este grupo se engloban a un conjunto de venas con un diámetro de entre 1mm y 1cm en las que drenan<br />
las vénulas, y son características algunas <strong>com</strong>o las venas de las extremidades que son distales a la vena braquial en el<br />
MS (es decir, las venas cefálica y basílica) y a la poplítea en el MI (venas safena magna mayor y menor), las venas<br />
viscerales, las de la piel y las de la cabeza (no las de los troncos primarios).<br />
Van siempre a<strong>com</strong>pañadas de la arteria correspondiente y se diferencian de esta por tener paredes finas y una luz<br />
mayor (en las muestras suele colapsarse y aparecer sangre artefactada y luz ovalada).<br />
Las diferentes túnicas presentan:<br />
TÚNICA ÍNTIMA: está formada por células endoteliales cuya membrana basal descansa en un subendotelio con<br />
fibroblastos, fibras reticulares y algunas elásticas.<br />
TÚNICA MEDIA: formada por tejido conjuntivo con fibroblastos y células musculares lisas. Está más desarrollada en<br />
las venas del MI, del útero grávido y en la vena umbilical, pero es más pequeña en las venas de la retina, meníngeas,<br />
placenta, pene y bazo.<br />
TÚNICA ADVENTICIA: está muy desarrollada y contiene fibroblastos, fibras colágenas y elásticas y algunas fibras<br />
nerviosas amielínicas.<br />
Válvulas<br />
Además, al igual que las Venas de Gran Calibre presentan una serie de válvulas que impiden el retorno venoso. Estas<br />
valvas son evaginaciones o repliegues semilunares que tienen un eje central de tejido conectivo revestido por células<br />
endoteliales que están en contacto con la sangre. En una válvula venosa se pueden distinguir, además, dos<br />
estructuras: el seno valvular (espacio <strong>com</strong>prendido entre la pared vascular distal a la válvula y la propia válvula) y la<br />
<strong>com</strong>isura (zona engrosada de la válvula donde se inserta el tejido conjuntivo laxo).<br />
La apertura de las válvulas se da por la contracción de los músculos que la rodean.
Página15<br />
En este dibujo (que muestra a una vena de mediano calibre del MI)<br />
se observa cómo se abre una válvula:<br />
En un momento t0 las dos válvulas están cerradas y los músculos que<br />
rodean la vena están relajados.<br />
En un momento t1 los músculos se contraen y <strong>com</strong>primen la luz de la<br />
vena, por lo que la sangre superior al punto de contracción sigue una<br />
trayectoria hacia arriba y la que es inferior al punto de contracción va<br />
hacia abajo.<br />
En un momento t2, el aumento de P en cada válvula tiene dos efectos<br />
contrarios: la válvula proximal se abre y permite el paso de la sangre<br />
en dirección hacia el corazón, pero la válvula distal permanece<br />
cerrada e impide el flujo retrógrado de la sangre.<br />
Venas de Gran Calibre<br />
Son venas con un gran diámetro (>1cm), <strong>com</strong>o es el caso de las venas pulmonares, la vena porta, las cavas, la yugular<br />
o la subclavia.<br />
Sus diferentes túnicas se encuentran mucho menor delimitadas y presentan:<br />
TÚNICA ÍNTIMA: formada por un endotelio típico con subendotelio formado por fibroblastos, fibras colágenas y<br />
elásticas, que forman una LEI no continua.<br />
TÚNICA MEDIA: formada por fibras musculares lisas sostenidas por un tejido conjuntivo mantenido por fibroblastos;<br />
tiene un mayor desarrollo en las venas pulmonares.<br />
TÚNICA ADVENTICIA: está muy desarrollada porque aparecen fibroblastos, fibras colágenas y elásticas, células<br />
musculares lisas y anejos <strong>com</strong>o vasa vasorum, nervi vascularis y vasos linfáticos. Está aún más desarrollada en la vena<br />
cava inferior y en la suprarrenal, pero menos en la cava inferior, venas yugulares, subclavias y el tronco<br />
braquiencefálico.<br />
Características Especiales de algunas Venas<br />
Fibras musculares subintimales longitudinales en venas ilíaca, femoral, poplítea y safenas.<br />
Células musculares cardíacas en la íntima de las venas cavas y pulmonares (drenan en las aurículas cadíacas).<br />
Ausencia de la capa media: vena cava<br />
inferior (principalmente de la capa muscular).<br />
Capa media muy desarrollada con fibras musculares circulares: vena pulmonar.<br />
Además existen algunas venas atípicas, <strong>com</strong>o los senos venosos o durales que aparecen en la cavidad craneana;<br />
estos son espacios en la duramadre revestidos por células endoteliales.<br />
En las venas es frecuente, generalmente en mujeres, una patología llamada varices o dilataciones varicosas. Se deben<br />
a debilitamientos de la pared vascular por múltiples causas (bipedestación excesiva, <strong>com</strong>presión por el parto o una<br />
patología sistémica <strong>com</strong>o una cardiopatía) que por la degradación del colágeno (por proteasas) produce dilataciones<br />
a nivel de un segmento venoso e impide la vuelta correcta de la sangre al corazón.
Página16<br />
VASOS LINFÁTICOS<br />
Los vasos linfáticos recogen y redistribuyen la linfa a lo largo del organismo en un sentido unidireccional: de los<br />
tejidos al corazón.<br />
Son muy frecuentes en el tejido conectivo subyacente a la piel y a las mucosas (<strong>com</strong>o en el intestino delgado),<br />
aunque son inexistentes:<br />
<br />
<br />
<br />
Cartílago y Hueso<br />
Epidermis<br />
Tejido Nervioso<br />
<br />
<br />
<br />
Órbita<br />
Médula ósea roja<br />
Oído interno<br />
<br />
<br />
<br />
Placenta<br />
Diente<br />
Timo<br />
El sistema linfático <strong>com</strong>ienza en unos fondos de saco ciego que conforman los capilares linfáticos, que drenan en los<br />
vasos linfáticos (de mayor calibre), que finalmente drenan en los dos grandes conductos linfáticos del organismo: el<br />
conducto torácico y la gran vena linfática derecha.<br />
Capilar Linfático<br />
Tiene en su capa íntima un endotelio unido por uniones estrechas (menos que en los vasos sanguíneos), nunca<br />
aparecen poros ni fenestraciones, no hay cuerpos de Weibel-Palade (no intervienen en la coagulación) ni hay células<br />
pericíticas.<br />
En cambio, su lámina basal es discontinua y presenta unos filamentos de anclaje o ligadura hacia las fibras de<br />
colágeno subyacentes. Estos filamentos impiden que la luz vascular se colapse en los momentos de mayor presión<br />
tisular, <strong>com</strong>o en la inflamación.<br />
Una de las principales características de los capilares linfáticos es su elevada permeabilidad, mucho mayor que la de<br />
los vasos sanguíneos. Así son mucho más eficaces en extraer el líquido rico en P del LEC. En el caso del intestino<br />
delgado se conocen <strong>com</strong>o vasos quilíferos centrales, y también recogen otras biomoléculas <strong>com</strong>o los lípidos,<br />
colaborando en la absorción.<br />
Vasos Linfáticos<br />
En los vasos linfáticos ya se pueden observar 3 capas mal delimitadas: túnicas íntima, media y adventicia (poco<br />
desarrollada).<br />
Tienen un mayor diámetro (>0,2mm) y su pared se engruesa, por lo que se pueden observar mayor agregado de<br />
tejido conectivo y haces de músculo liso, así <strong>com</strong>o de válvulas que impiden el reflujo linfático y contribuyen al<br />
sentido unidireccional. De todos modos, el movimiento linfático se da gracias a la contracción de los músculos que<br />
rodean al vaso, y son las válvulas las que impiden que la linfa vaya en el sentido incorrecto. También interviene la<br />
musculatura lisa de la pared.<br />
Por último, estos vasos linfáticos drenan en los grandes vasos linfáticos, de mayor grosor pero características<br />
similares al resto de vasos linfáticos.<br />
El sistema linfático tiene una gran importancia en la distribución de las células malignas en caso de tumoración. Pero<br />
también hay patologías producidas por un aumento en el flujo linfático, <strong>com</strong>o el edema (<strong>com</strong>o consecuencia de un<br />
aumento en la permeabilidad del capilar sanguíneo y la P sanguínea, pero disminución de la P coloidosmótica), o por<br />
una obstrucción de los vasos linfáticos, lo que ligado a otros factores <strong>com</strong>o la inflamación puede producir un<br />
hidrotórax o un linfedema.
Página17<br />
ESTRUCTURAS VASCULARES ESPECIALES<br />
En general, en el Sistema Vascular las arterias se van ramificando hasta que en los capilares se produce el<br />
intercambio con los tejidos y la sangre resultante drena en las venas. Pero existe una serie de excepciones cuyo curso<br />
es diferente y cumplen diferentes funciones en el organismo.<br />
Sistema Portal<br />
Comprende a dos redes de capilares conectadas por un vaso de mayor calibre, generalmente una vena. Existen<br />
muchos ejemplos y en cada uno todos tienen diferentes funciones:<br />
<br />
<br />
<br />
<br />
Sistema Porta Hepático: permite el transporte de nutrientes desde el punto de absorción (capilares<br />
intestinales) al centro de procesamiento metabólico del organismo (el hígado) a través de la Vena Porta.<br />
Sistema Porta Hipofisiario: conecta una red de capilares en la porción nerviosa de la hipófisis (Neurohipófisis)<br />
con la porción endocrina (Adenohipófisis), de modo que regula con diversas sustancias liberadas en los<br />
capilares de la Neurohipófisis la secreción de hormonas en la Adenohipófisis.<br />
Sistema Porta Ínsulo-Acinar: sistema porta que conecta un conjunto de capilares que irrigan a los Islotes de<br />
Langerhans pancreáticos (glándula endocrina) con las glándulas acinares pancreáticas (glándulas exocrinas).<br />
De este modo, según qué hormona se secrete en el Islote de Langerhans se activará o inhibirá la secreción de<br />
jugo pancreático por la porción exocrina pancreática.<br />
Sistema Porta Renal: está formado por una arteria aferente que forma una red de capilares, el glomérulo,<br />
donde se produce el ultrafiltrado de la sangre y la producción de la primera orina, y una arteria eferente que<br />
lleva la sangre filtrada a un segundo <strong>com</strong>plejo capilar donde hay intercambio con el tejido renal.<br />
Cuerpo Aórtico, Cuerpo y Seno carotídeo<br />
Los Cuerpos Aórticos y Carotídeo son dos estructuras que tienen quimiorreceptores que detectan la <strong>com</strong>posición<br />
química de la sangre para responder eficazmente; el seno carotídeo detecta cambios en la P gracias a<br />
barorreceptores.<br />
Cuerpo Carotídeo<br />
El cuerpo carotídeo o Glomus Carotídeo es una estructura par que se localiza en la bifurcación de la arteria carótida<br />
<strong>com</strong>ún en sus ramas externas e internas; su diámetro mayor tiene un tamaño de 0,5cm.<br />
Presenta un quimiorreceptor que controla la presión parcial de O 2 , CO 2 y H + , de modo que controla estados<br />
patológicos <strong>com</strong>o la hipoxia, la hipercapnia o la acidosis y permite responder con cambios en la frecuencia<br />
respiratoria. Está inervado por los nervios vago y glosofaríngeo.<br />
Está formado por un ovillo de sinusoides con dos tipos de células endoteliales:<br />
<br />
<br />
Células Epiteloides tipo I o Células Glómicas de Masson: se asocian a las terminaciones nerviosas aferentes<br />
del nervio de Hering. Contienen gránulos que liberan catecolaminas, principalmente dopamina (DA) que<br />
estimulan los receptores dopaminérgicos D2 de este nervio en situación de hipoxia. Estas células tienen<br />
canales de K sensibles a O 2 cuya conductancia disminuye durante la hipoxia, por lo que disminuye la pO 2<br />
estos canales permiten que se despolarice la célula, entre Ca 2+ y se desencadena un V A que permite la<br />
liberación de los gránulos de catecolaminas.<br />
Células Epiteliodes tipo II o en Vaina: actúan <strong>com</strong>o células de glía que actúan <strong>com</strong>o soporte.<br />
Tiene un gran desarrollo en niños, personas jóvenes y aquellas que viven a grandes alturas; disminuye con la edad.
Página18<br />
Cuerpo Aórtico<br />
El cuerpo aórtico es una estructura par con dos localizaciones diferentes: el derecho entre la carótida <strong>com</strong>ún derecha<br />
y la subclavia derecha, y el izquierdo es medial al origen de la arteria subclavia izquierda.<br />
Tiene una estructura y función semejante al cuerpo carotídeo.<br />
Seno Carotídeo<br />
Es también una estructura par de 3-5mm que se sitúa en la dilatación de la bifurcación de la arteria carótida primitiva<br />
derecha e izquierda. Contienen barorreceptores que permiten medir la P arterial y responder ante cambios en esta.<br />
Tiene una túnica media muy fina con una túnica adventicia altamente inervada por el IX par craneal.<br />
Anastomosis Arteriovenosas (AV)<br />
Son <strong>com</strong>unicaciones directas entre arterias y venas que desvían sangre a los capilares. Se sitúan <strong>com</strong>únmente en la<br />
piel de la punta de los dedos, la nariz y los labios, así <strong>com</strong>o en los tejidos eréctiles del pene y clítoris. La arteriola de la<br />
<strong>com</strong>unicación AV suele estar enrollada y tiene en su capa media una gruesa capa de tejido muscular liso muy<br />
inervada por el SN Vegetativo. Esta capa muscular permite la termorregulación según estímulos neurológicos y<br />
hormonales (su contracción es pulsátil):<br />
<br />
<br />
Contracción del músculo liso: la sangre pasa al lecho capilar, de modo que se pierde calor (aumenta la<br />
superficie de la sangre con el medio).<br />
Relajación del músculo liso: la sangre pasa a la vena y no se pierde calor.<br />
En los tejidos eréctiles, <strong>com</strong>o el pene, el cierre de la anastomosis dirige el flujo sanguíneo hacia los cuerpos<br />
cavernosos y se produce la respuesta eréctil.<br />
Glomus o Canal de Sucquet-Hover<br />
El Glomus es un canal contorneado por anastomosis AV que están tapizados por células endoteliales. Es<br />
característica su capa media, formada por células musculares lisas muy inervadas por el SN Vegetativo; existen 2<br />
tipos celulares:<br />
- Células Epiteloides tipo I o Glómicas de Masson: aparecen de color claro.<br />
- Células Epiteloides tipo II: aparecen de color oscuro.<br />
Son abundantes en zonas acras (<strong>com</strong>parten localización don las anastomosis AV) y según los estímulos se produce:<br />
<br />
<br />
Activación SN S: apertura de la anastomosis por relajación de las células glómicas; se conserva calor.<br />
Activación SN PS: se cierra la anastomosis por la contracción de las células glómicas; se pierde calor.
Página19<br />
SISTEMA RESPIRATORIO<br />
El aparato respiratorio está formado por dos pulmones y una serie de vías aéreas que los <strong>com</strong>unican con el exterior.<br />
Funciones<br />
<br />
<br />
<br />
Conducción del aire<br />
Filtración y calentamiento del aire<br />
Intercambio gaseoso<br />
Además, el paso del aire a través de la laringe permite la fonación y el epitelio olfatorio permite percibir olores.<br />
Partes<br />
El Sistema Respiratorio se puede dividir en una porción conductora, que conducen el aire a los pulmones y no tienen<br />
función de intercambio gaseoso (hay una porción extrapulmonar, que <strong>com</strong>prende de las cavidades nasales a los<br />
bronquios principales, y otra intrapulmonar que <strong>com</strong>prende a los bronquios internos y al árbol bronquial) y una<br />
porción respiratoria (bronquiolos respiratorios, alveolos y conductos y sacos alveolares).<br />
Tipos Epitelio Presente en el Aparato Respiratorio<br />
<br />
<br />
<br />
<br />
Pavimentoso Tegumentario: epitelio estratificado plano que contiene glándulas sebáceas, sudoríparas y<br />
vibrisas<br />
Mucoso: plano estratificado no queratinizado<br />
Respiratorio: epitelio cilíndrico seudoestratificado y ciliado<br />
Olfatorio o neuroepitelio olfatorio<br />
CAVIDADES NASALES<br />
Los orificios A, las narinas, <strong>com</strong>unican con los orificios P, las coanas, que dan paso a la faringe. Entre ambos orificios<br />
aparecen las dos cavidades nasales separadas por un tabique óseo y cartilaginoso. Cada cámara se divide en otras 3<br />
regiones: vestíbulo o narina, segmento olfatorio y segmento respiratorio.<br />
Vestíbulo<br />
El vestíbulo está revestido por un epitelio estratificado plano (pavimentoso tegumentario) que se continúa con la piel<br />
de la cara. Contiene glándulas sebáceas (sin folículo piloso), alguna glándula sudorípara y pelos rígidos llamados<br />
vibrisas.<br />
En la pared posterior del vestíbulo hay epitelio seudoestratificado con células ciliadas y con células caliciformes<br />
intercaladas (epitelio respiratorio). Las células caliciformes segregan producto mucoso (en hipofase, es decir, estado<br />
líquido acuoso) que se extiende sobre el epitelio. El moco se desplaza gracias a los cilios, que lo baten a una<br />
velocidad 1 cm / min. Este epitelio se encuentra en todo el aparato respiratorio excepto en las regiones más<br />
expuestas: cornetes y cuerdas vocales verdaderas, donde el epitelio es plano estratificado.<br />
En la nariz, las vibrisas se encargan de filtrar las partículas. El moco atrapa el polvo e impide la deshidratación. El<br />
moco es transportado por los cilios hacia la faringe.<br />
La enfermedad de Kartagener se produce porque los cilios quedan inmóviles, de modo que los pacientes son<br />
propensos a padecer sinusitis y bronquitis. El tabaquismo altera el patrón del batido ciliar, que pierden la sincronía y<br />
reducen su frecuencia de batido. Además altera el número de células caliciformes.
Página20<br />
Segmento Respiratorio<br />
El segmento respiratorio de las cavidades nasales <strong>com</strong>pone la mayor parte del volumen de las cavidades nasales. Se<br />
puede diferenciar una pared medial lisa que corresponde al tabique nasal y unas paredes laterales con repliegues<br />
debidos a 3 proyecciones óseas: los cornetes o coanas.<br />
La porción respiratoria de la nariz está tapizada por epitelio respiratorio. En esta zona aparecen 5 tipos celulares:<br />
Células ciliadas o Células Columnares Ciliadas (aproximadamente 300 cilios por célula) (30%)<br />
Células Caliciformes (30%)<br />
Células con Ribete en Cepillo (con microvellosidades romas) (5%)<br />
Células de Kulchitsky o Células con Gránulos Pequeños (Small Granule Cells): pertenecen al sistema APUD o<br />
neuroendocrino difuso. Histológicamente son similares a las células basales, pero con gránulos de secreción.<br />
Siguen apareciendo a lo largo del sistema respiratorio (mirar Tráquea).<br />
Células Basales (células madre pluripotenciales/stem cells) (30%).<br />
En la lámina propia de la pared media encontramos glándulas serosas y mucosas y muy pocos vasos sanguíneos<br />
(poco vascularizada) y bajo ella se sitúa el periostio o pericondrio. Aparecen células migradas del sistema inmune,<br />
por lo que en rinitis alérgicas aparecen mastocitos y además se produce un paso del plasma al conjuntivo (conjuntivo<br />
edematoso) y secreción masiva de Ig A, E y G.<br />
Pared Lateral o Pituitaria Roja<br />
El suelo y las paredes laterales de la cavidad nasal es lo que se conoce <strong>com</strong>o pituitaria roja: la mucosa que recubre<br />
los cornetes inferior y medio. Es una mucosa muy vascularizada, por lo que es frecuente que sangre.<br />
Los capilares de la pituitaria roja se sitúan en la lámina propia paralelos entre sí y en perpendicular al flujo del aire.<br />
Son capilares muy superficiales, lo que permite el flujo de aire que llega es directamente calentado por sangre que<br />
corre en sentido opuesto. Encontramos también un plexo venoso superficial y grande, con propiedades eréctiles.<br />
Existen asimismo anastomosis AV de tipo enrollado. Estos vasos pueden aumentar de tamaño y filtrar líquido en las<br />
reacciones alérgicas o infecciones virales, <strong>com</strong>o en el refrío <strong>com</strong>ún. Entonces la lámina propia se distiende con el<br />
líquido y se produce una marcada tumefacción de la membrana mucosa.<br />
Los propios <strong>com</strong>ponentes vasculares permiten grandes variaciones de flujo, debido a haces musculares a modo de<br />
esfínter situados en las paredes venosas (ver anastomosis AV).<br />
Además, está inervada por el SN Vegetativo: la inervación simpática provoca vasoconstricción, mientras que la<br />
parasimpática provoca vasodilatación y secreción glandular. Los aerosoles nasales provocan vasoconstricción por<br />
acción de agentes adrenérgicos y eliminan así las obstrucciones nasales.<br />
Los propios cornetes aumentan la superficie de contacto del aire con la sangre y facilitan el acondicionamiento del<br />
aire y, además, transforman el flujo laminar del aire en uno turbulento de modo que las partículas que contiene el<br />
aire chocan contra el moco de las paredes (secretado por las células caliciformes y las glándulas de la lámina propia)<br />
y se produce la precipitación turbulenta de estas partículas donde quedan adheridas.<br />
También son frecuentes en la lámina propia las glándulas mucosas con semilunas serosas.<br />
En resumen, las funciones de la pituitaria roja son acondicionar aire y absorber vapores hidrosolubles (amoniaco,<br />
formaldehído).<br />
Zona de Kiesselbach: zona muy vascularizada por una <strong>com</strong>unicación AV entre el plexo de la arteria esfenopalatina y<br />
etmoidal y las venas correspondientes; permiten mantener una temperatura constante.
Página21<br />
Segmento Olfatorio<br />
El Segmento Olfatorio es una superficie de 5cm 2 (antiguamente conocido <strong>com</strong>o pituitaria amarilla) que se sitúa en la<br />
cúpula de la cavidad nasal y una pequeña parte de las paredes laterales y mediales, sobre el periostio. Presenta un<br />
color amarillento que se debe acumulaciones de lipofuscina en el epitelio y glándulas olfatorias.<br />
Esquema del Neuroepitelio<br />
Neuroepitelio<br />
El epitelio olfatorio es seudoestratificado, tiene un grosor de 50μm y aparecen 4 tipos celulares: células basales,<br />
células de sostén, en cepillo y olfatorias.<br />
CÉLULAS OLFATORIAS<br />
Las células olfatorias son neuronas de tipo bipolar, con un número de aproximadamente 10 millones en nuestro<br />
organismo.<br />
La prolongación apical es la dendrita, que se abre paso por las células de sostén hasta la superficie, donde se<br />
ensancha formando una estructura llamada maza o vesícula olfatoria, de la que parten quiniocilios (cilios inmóviles<br />
con cuerpos basales típicos) muy largos, dispuestos de manera radial y se disponen sobre las microvellosidades de las<br />
células de sostén, si bien están incluidos dentro de la capa de moco. La membrana plasmática de cada quiniocilio<br />
tiene muchos receptores.<br />
La prolongación hacia basal es el axón, que atraviesa la membrana basal y en la lámina propia se une a otras<br />
neuronas conformando el nervio olfativo o I par craneal, que atraviesa la lámina cribosa del etmoides y se dirige al<br />
bulbo olfatorio.<br />
A pesar de ser neuronas, si se lesionan son reemplazadas. De hecho, su tasa de renovación es de 1 o 2 meses,<br />
aunque este intervalo disminuye con la edad provocando una pérdida de agudeza olfativa.<br />
CÉLULAS DE SOSTÉN<br />
Las células de sostén son las más numerosas. Son altas cilíndricas, con microvellosidades y su función es el apoyo<br />
metabólico y físico a células olfativas. Su núcleo es ovalado y se dispone en la zona apical. Presenta gránulos de<br />
lipofuscina, REL y RER abundantes, así <strong>com</strong>o mitocondrias y uniones estrechas con las células vecinas, excepto con<br />
las olfatorias con las que se une por uniones adhesivas.
Página22<br />
CÉLULAS BASALES<br />
Las células basales son stem cells que permiten la renovación de epitelio porque pueden dar lugar a cualquier tipo<br />
celular.<br />
No alcanzan la luz, su núcleo es más basófilo y se encuentra en la zona basal. Son capaces de emitir prolongaciones<br />
que envainan parcialmente al axón de las células olfativas.<br />
CÉLULAS EN CEPILLO<br />
Se encuentran en mucha menor cantidad; son células cilíndricas con grandes vellosidades romas en su superficie<br />
apical. En cambio, en su superficie basal sinaptan con fibras nerviosas terminales del nervio trigémino (V par craneal)<br />
que interviene en la sensibilidad general, por lo que estas células permiten que la mucosa esté sensibilizada.<br />
Lámina Propia<br />
La lámina propia se continúa directamente con el periostio del hueso adyacente. En su seno hay una gran cantidad<br />
de vasos sanguíneos y linfáticos, nervios olfatorios amielínicos, nervios mielínicos y glándulas olfatorias o de<br />
Bowman.<br />
Las Glándulas Olfatorias o de Bowman son glándulas serosas tubuloalbeolares ramificadas que secretan una<br />
sustancia serosa (proteinácea) que permite atrapar y disolver sustancias con aroma para eliminar aquellas moléculas<br />
ya percibidas.<br />
Las células glandulares tienen gránulos de lipofuscina que, junto a los de las células de sostén, dan el color pardoamarillento<br />
al tejido. Los conductos están formados por una sola capa de células cuboides que según van<br />
acercándose al neuroepitelio se van haciendo planas.<br />
En la lámina propia también hay células propias del sistema inmune, <strong>com</strong>o células plasmáticas, que secretan lisozima<br />
e Ig A.<br />
Mecanismo del Olfato<br />
Existen unos 1000 genes que codifican los receptores olfativos. Cada célula olfativa expresa solo uno de esos genes,<br />
de modo que en cada célula olfativa hay un solo tipo de receptor.<br />
Las sustancias odoríferas se unen a OBP (Odorant Binding Protein), y ese <strong>com</strong>plejo alcanza los receptores olfativos de<br />
los quiniocilios, difundiendo por el moco. Así, un único receptor puede detectar múltiples sustancias odoríferas. Ese<br />
receptor esta acoplado a una proteína G produciendo AMPc, fosfolipasa C y PIP2. Esta reacción puede producir la<br />
despolarización de la membrana, generando así un impulso nervioso. El OBP es reciclado después de cada olfacción.<br />
Senos Paranasales<br />
Son cavidades aéreas excavadas en huesos de la cabeza: maxilar, etmoidal, frontal, esfenoidal. Son estructuras pares,<br />
en <strong>com</strong>unicación directa con cavidad nasal.<br />
Los senos paranasales están tapizados por mucosa respiratoria, con epitelio menos grueso y con gran cantidad de<br />
células caliciformes.<br />
El moco de los senos paranasales se desplaza hacia la cavidad nasal y en caso de inflamación la <strong>com</strong>unicación puede<br />
obstruirse, de modo que se retiene el moco provocando sinusitis.<br />
FARINGE<br />
Conduce el aire a la laringe (ver Aparato Digestivo).
Página23<br />
LARINGE<br />
La laringe es un conducto de 4-5cm del aparato respiratorio que une la orofaringe con la tráquea, aunque no<br />
libremente porque en su extremo superior está cubierta por la epiglotis.<br />
La laringe se divide en:<br />
La supraglotis consta de la epiglotis, el primer par pliegues vocales y los ventrículos laríngeos o de Morgagni.<br />
La glotis la forman los segundos pliegues vocales, y las <strong>com</strong>isuras anterior y posterior.<br />
La subglotis es la zona de la laringe que se extiende bajo los segundos pliegues vocales hasta el cartílago cricoides.<br />
Mucosa laríngea<br />
El epitelio respiratorio de la laringe es de dos tipos:<br />
Epitelio Mucoso: cuerdas vocales verdades & cara lingual y tercio superior de la cara laríngea de la epiglotis.<br />
Epitelio Respiratorio: resto de la laringe.<br />
Además, presenta una capa de moco que es batida hacia la faringe.<br />
Debajo encontramos una lámina propia laxa; es frecuente que se den edemas con asfixia y falso crup por infecciones<br />
virales.<br />
Ya más en profundidad, la lámina propia es más densa y presenta glándulas seromucosas y abundantes folículos<br />
linfoides. Al final de la lámina propia encontramos fibras elásticas que conforman la membrana fibroelástica, bajo la<br />
que está el hueso, cartílago o músculo esquelético.<br />
Cartílagos<br />
En la faringe encontramos cartílagos rodeados por abundante pericondrio de conjuntivo denso que, además, permite<br />
la unión entre las placas cartilaginosas.<br />
Algunos de los cartílagos son pares (aritenoides, corniculados y cuneiformes) y otros impares (tiroides, cricoides,<br />
epiglotis). De todos modos, todos son cartílagos hialinos, excepto la epiglotis, que es cartílago elástico.<br />
Todos estos cartílagos calcifican con la edad.<br />
Músculos esqueléticos<br />
Los músculos de la laringe pueden ser intrínsecos o extrínsecos:<br />
- Los músculos intrínsecos se sitúan entre las placas de cartílago y tensan los pliegues, permitiendo regular la<br />
apertura y cierre de la glotis, es decir, la fonación.<br />
- Los músculos extrínsecos se sitúan uniendo la laringe con estructuras vecinas y evitando el paso del aire al<br />
esófago y de los alimentos a la tráquea.<br />
Esta musculatura está implicada en el reflejo de la tos: cuando un agente extraño se introduce en la laringe,<br />
se produce una inspiración forzada de manera que se cierra la glotis y se retraen los pliegues vocales. Para<br />
expulsar este agente irritante, la musculatura intercostal y la abdominal se contraen, produciéndose de<br />
manera refleja la apertura de la epiglotis; así el aire que se había inspirado sale proyectado abruptamente<br />
hacia fuera a una velocidad de 180 km/h.<br />
Pliegues vocales<br />
Cuerdas vocales falsas o ventriculares<br />
El primer par de pliegues vocales son las cuerdas vocales falsas o ventriculares. Están cubiertas por epitelio<br />
respiratorio, si bien es frecuente encontrar parches de epitelio mucoso.<br />
La lámina propia es laxa y presenta glándulas seromucosas que pueden introducirse en la epiglotis y lubrican los<br />
pliegues. No hay musculatura intrínseca, por lo que no está implicado en la fonación y su función es eminentemente<br />
protectora.
Página24<br />
Entre estos pliegues y los siguientes, encontramos una zona hendida conocida <strong>com</strong>o Ventrículo Laríngeo o de<br />
Morgagni, que se prolonga con forma de sáculo. Está tapizado por epitelio respiratorio, con una lámina propia laxa<br />
con glándulas seromucosas. Participa en la resonancia, intensificando el sonido en las cuerdas vocales verdaderas.<br />
Cuerdas vocales verdaderas<br />
El segundo par de pliegues vocales son las cuerdas vocales verdaderas, tapizadas por epitelio mucoso, con presencia<br />
de melanocitos. Este epitelio ofrece mejor protección a esta zona frente a la abrasión que supone el paso del aire.<br />
Encontramos melanocitos.<br />
La lámina propia se suele dividir en una zona superficial o espacio de Reinke y unas capas media y profunda que<br />
conforman el ligamento vocal.<br />
Bajo la lámina propia encontramos el musculo vocal o tiroaritenoideo.<br />
El espacio de Reinke es un conjuntivo cuyo <strong>com</strong>ponente fundamental es la matriz extracelular rica en fibras elásticas,<br />
pero muy pobre en células y vascularización y carente en glándulas ni vasos linfáticos.<br />
Los ligamentos vocales son conjuntivo con predominancia total de fibras elásticas que forman haces paralelos. En<br />
ellos encontramos mastocitos, por lo que en esta zona se pueden dar reacciones de hipersensibilidad (falso crup), con<br />
edemas y obstrucción laríngea<br />
Estos segundos pliegues vocales se dividen en una zona cubierta, <strong>com</strong>puesta del epitelio y el espacio de Reinke, y un<br />
centro, formado por el ligamento vocal y el músculo tiroaritenoideo. El centro es rígido, mientras que la cubierta es<br />
flexible y lubricada por las secreciones glandulares.<br />
Hay a lo largo de la laringe células de Kultchisky, de tipo neuroendocrinas que forman parte del sistema APUD (mirar<br />
Tráquea).<br />
Esquema estructura de la laringe
Página25<br />
TRÁQUEA<br />
La tráquea es un tubo corto de alrededor 2,5cm de diámetro y 10cm de longitud que permite conducir el aire desde<br />
la laringe a los bronquios. Se pueden distinguir 4 capas muy bien diferenciadas: mucosa, submucosa, cartílago y<br />
adventicia.<br />
Mucosa Traqueal<br />
Epitelio Traqueal<br />
El epitelio traqueal es un epitelio seudoestratificado ciliado (respiratorio) que presenta varios tipos celulares:<br />
CÉLULAS CILIADAS<br />
Es el más numeroso de los tipos celulares y aparecen en el espesor de todo el epitelio. Son células epiteliales con<br />
queratina K8 y una serie de cilias que se proyectan sobre la superficie apical, donde aparece una línea oscura<br />
formada por los cuerpos basales ciliares agregados.<br />
Las cilias permiten un movimiento de barrido de la capa mucosa hacia la faringe.<br />
CÉLULAS MUCOSAS<br />
Aparecen a lo largo de todo el epitelio dispersas entre las células ciliadas. Poseen microvellosidades más o menos<br />
escasas, carecen de cilios y presentan gránulos de mucinógeno acumulados en su citoplasma apical que con la<br />
tinción de HE aparecen <strong>com</strong>o zonas claras, y <strong>com</strong>o PAS+ con la tinción del ácido periódico de Schiff. Son muy<br />
similares a las células caliciformes intestinales, por lo que también se las llama en ocasiones células caliciformes<br />
(traqueales).<br />
CÉLULAS EN CEPILLO<br />
Se encuentran en mucha menor cantidad; son células cilíndricas con grandes vellosidades romas en su superficie<br />
apical y sinapsis con fibras nerviosas aferentes en su superficie basal, de modo que intervienen en la sensibilidad<br />
general permitiendo la sensibilidad de la mucosa.<br />
CÉLULA CON GRÁNULOS PEQUEÑOS O DE KULCHITSKY<br />
Son células que forman parte del sistema APUD y se suelen distinguir por tinción argéntica, por lo que en ocasiones<br />
también se las conoce <strong>com</strong>o células argentófilas o argentofines.<br />
Tienen el núcleo en posición basal y poseen una serie de pequeños gránulos que contienen hormonas tales <strong>com</strong>o<br />
catecolaminas (adrenalina y noradrenalina), serotonina, calcitonina, somatostatina, ADH o ACTH, que secretan al<br />
exterior Algunas de estas células están inervadas formando agrupaciones con las fibras nerviosas llamadas cuerpos<br />
neuroepiteliales, de modo que se cree que intervienen en reflejos que regulan las vías aéreas o los calibres<br />
vasculares.<br />
Los cuerpos neuroepiteliales tienen una gran inervación aferente y se <strong>com</strong>portan <strong>com</strong>o quimiorreceptores que<br />
detectan cambios en la pO2, de manera que esta información es conducida hasta el SNC. Así, estos cuerpos<br />
intervienen en la regulación de la perfusión pulmonar y en el control del estado de contracción del musculo a nivel<br />
bronquial. También son importantes durante el desarrollo pulmonar, porque regulan el crecimiento de los lobulillos<br />
pulmonares.<br />
Estas células son las que con mayor frecuencia se malignizan formando carcinoides y carcinomas: crecen aumentando<br />
el tamaño de los cuerpos neuroepiteliales y se liberan después, metastatizando.<br />
Su presencia en el sistema respiratorio tiene un explicación embriológica: el aparato respiratorio (incluidos los<br />
pulmones) derivan de una evaginación del intestino A embrionario, donde son abundantes las células del sistema<br />
APUD.
Página26<br />
CÉLULAS BASALES<br />
Son stem cells que mantienen el epitelio traqueal. Tienden a ser células prominentes y se sitúan en la parte basal del<br />
epitelio. Suponen el 30% del epitelio y su marcador es el K14.<br />
Membrana Basal y Lámina Propia<br />
Por debajo del epitelio traqueal aparece una lámina propia muy rica e fibras elásticas y células del sistema inmune,<br />
<strong>com</strong>o linfocitos, plasmocitos, mastocitos, eosinófilos y fibroblastos.<br />
En todo el sistema respiratorio de conducción, y especialmente en la lámina propia y en la submucosa de la tráquea,<br />
aparece una presencia constante de tejido linfoide de manera difusa y nodular que forman el BALT o MALT<br />
específico de la mucosa respiratorio (MALT = Tejido Linfoide Asociado a Mucosas).<br />
La parte más superior de esta lámina propia conforma la llamada membrana basal, una capa de aspecto vítreo u<br />
homogéneo que apenas se colorea. Tiene un grosor de 25-40μm y a la microscopía electrónica se revela una gran<br />
cantidad de fibras colágenas tipo IV (o fibras reticulares) densamente agrupadas. Así, esta zona es una capa reticular<br />
muy gruesa y densa, pero sigue formando parte de la lámina propia. En fumadores esta membrana basal se hace aún<br />
más gruesa de lo normal <strong>com</strong>o respuesta a la irritación de la mucosa.<br />
En la zona final de la lámina propia aparece una zona con abundantes fibras elásticas llamada membrana elástica;<br />
marca el límite entre la lámina propia y la submucosa.<br />
Submucosa Traqueal<br />
Es diferente al resto de órganos porque es de carácter laxo, similar a la lámina propia en aspecto. En su seno aparece<br />
el BALT, vasos sanguíneos y linfáticos y glándulas submucosas formadas por acinos secretores de carácter mucoso<br />
con semilunas serosas (más frecuentes en la brecha cartilaginosa).<br />
Cartílagos Traqueales y Adventicia Traqueal<br />
En la tráquea humana aparecen de 16 a 20 cartílagos hialinos con forma de C, quedando una brecha en P orientada<br />
hacia el esófago. Esta brecha está cubierta por tejido fibroelástico, una musculatura lisa que <strong>com</strong>pone el músculo<br />
traqueal y los ligamentos anulares, de tejido conjuntivo denso rico en fibras elasticas dispuestas longitudinalmente.<br />
La función de estos cartílagos es evitar el colapso de la estructura en la espiración, dotándola de flexibilidad. Con la<br />
edad suelen osificarse, perdiendo flexibilidad, o también pueden anastomosarse unos con otros.<br />
La adventicia traqueal es periférica a los anillos cartilaginosos y al músculo traqueal y permite la fijación de este tubo<br />
a estructuras adyacentes del cuello y mediastino. También contiene a vasos sanguíneos y nervios mayores, que<br />
irrigan la pared P traqueal, así <strong>com</strong>o nódulos linfáticos de gran calibre que drenan la pared de linfa.
Página27<br />
BRONQUIOS<br />
La tráquea se divide en 2 ramas que forman los bronquios principales o extrapulmonares, que a nivel anatómico se<br />
diferencian <strong>com</strong>o bronquio derecho e izquierdo; el bronquio derecho es más ancho y corto que el izquierdo.<br />
Al ingresar en los pulmones se denominan bronquios intrapulmonares y rápidamente se dividen en los bronquios<br />
secundarios o lobulares. El bronquio intrapulmonar izquierdo se divide en 2 bronquios lobulares (tantos <strong>com</strong>o<br />
lóbulos tiene su pulmón) y el derecho en 3. A su vez, cada bronquio lobular izquierdo se divide y aparecen un total de<br />
8 bronquios segmentarios o terciarios, y en el lado derecho 10 correspondientes. Cada bronquio segmentario con el<br />
parénquima pulmonar asociado forman un segmento broncopulmonar, de gran importancia en la cirugía pulmonar<br />
para la resección, si es necesario, sólo de los segmentos afectados con sus anejos vasculares, nerviosos y de sostén<br />
correspondientes.<br />
Esquema de la división del Árbol Bronquial<br />
El conjunto de las divisiones consecutivas de los bronquios conforman el árbol bronquial; cada nueva división se<br />
conoce <strong>com</strong>o generación. En un <strong>com</strong>ienzo, los bronquios guardan la misma organización tisular que la tráquea, pero<br />
según se va dividiendo van apareciendo diferentes características según se avanza por los bronquios.<br />
Mucosa<br />
La mucosa es muy similar a la traqueal, con un epitelio respiratorio formado por células ciliadas, caliciformes, basales<br />
(células madre) y endocrinas, así <strong>com</strong>o una importante cantidad de linfocitos y mastocitos. Esta estructura se<br />
mantiene en las sucesivas ramificaciones bronquiales, aunque la altura de las células va disminuyendo.<br />
En la lámina propia hay abundantes glándulas seromucosas, con células secretoras y mioepiteliales. Segregan un<br />
conjunto de sustancias mucosas para recubrir la mucosa bronquial, así <strong>com</strong>o lisozimas y defensinas de carácter<br />
inmunitario. Es igual que la traqueal en el resto de características.<br />
Muscular, Submucosa, Capa Cartilaginosa y Adventicia<br />
La capa muscular es continua, y los miocitos están dispuestos en espiral. Puede ocurrir que en algunas secciones,<br />
sobre todo en ramificaciones avanzadas, parezca que es discontinua por la presencia de pliegues en la luz. Además,<br />
en las nuevas generaciones aparece más adelgazada y con organización más laxa.<br />
La submucosa se mantienen <strong>com</strong>o un tejido relativamente laxo y que en los bronquios de mayor diámetro presenta<br />
glándulas y tejido adiposo.<br />
La capa cartilaginosa consiste en placas discontinuas de cartílago hialino que disminuyen de tamaño a medida que<br />
decrece el diámetro bronquial. Tienen forma cilíndrica y según avanzamos en el árbol bronquial se hacen más<br />
pequeñas y escasas hasta que finalmente desaparecen en el punto que el diámetro es de 1mm (bronquiolo).<br />
La adventicia es un tejido conectivo de densidad moderada que aparece de forma continua con el de estructuras<br />
adyacentes <strong>com</strong>o la arteria o el parénquima pulmonares.
Página28<br />
BRONQUIOLOS<br />
Los bronquiolos son las vías respiratorias de diámetro inferior a 1 mm. Resultan de la subdivisión de cada segmento<br />
broncopulmonar en los lóbulos pulmonares, cada uno de ellos ventilados por un bronquiolo. Cada lóbulo se<br />
encuentra formado a su vez por varias unidades de estructura menores llamadas acinos pulmonares, formadas por<br />
un bronquiolo terminal que airea a los bronquiolos respiratorios y alveolos. En consecuencia la unidad funcional y<br />
más pequeña del pulmón es la estructura formada por un bronquiolo respiratorio con su alveolo.<br />
Mucosa<br />
Su mucosa consta de un epitelio respiratorio, que se transforma en epitelio simple cilíndrico ciliado según se<br />
estrecha el conducto, hasta que en los bronquiolos pequeños el epitelio es finalmente simple cúbico.<br />
En este tejido aparecen células endocrinas, que forman cuerpos neuroepiteliales, escasas células caliciformes (más<br />
abundantes en fumadores) y células de Clara.<br />
CÉLULAS DE CLARA<br />
La cantidad de células de Clara aumenta según avanzamos en el árbol bronquial, de modo que en los bronquiolos<br />
terminales alcanzan ya el 80%. Son células prismáticas, no ciliadas, con una zona apical abultada. Son células<br />
metabólicamente muy activas, con abundante RE (REL en la zona apical y RER en la zona basal) y gránulos de<br />
secreción.<br />
Estas células de Clara sintetizan un producto de naturaleza lipoproteica, no pegajoso, que recubre la superficie del<br />
bronquiolo de manera que rebaja la tensión superficial, evitando el colapso en la espiración.<br />
El marcador de estas células es CC16.<br />
Las células de Clara presentan en su membrana plasmática abundancia de canales de Cl llamados CFTR, de manera<br />
que puede que tengan un papel en la regulación del transporte de los iones Cl. En la fibrosis quística, el transporte de<br />
Cl- está afectado: las células no tienen canales o estos fallan por alguna mutación. Este canal permite que la célula<br />
transporte hacia fuera Cl- y HCO3-. Al estar ausente o deficiente este canal, la célula no puede eliminar el cloro de<br />
manera que capta exceso de Na y segrega un moco espeso. La fibrosis quística puede aparecer tambien en otros<br />
órganos.<br />
Esquema de una Célula de Clara<br />
La lámina propia de la mucosa bronquiolar es parecida a la de los bronquios: conjuntivo laxo con fibras elásticas, si<br />
bien en los bronquiolos NO encontramos glándulas seromucosas.
Página29<br />
Capa muscular y Adventicia<br />
La capa muscular la <strong>com</strong>pone el musculo de Reisseisen, resultando una capa gruesa pero no continua. Los miocitos<br />
de esta capa son sensibles a los cambios en la presión de O2, de manera que si baja, los mioitos se relajan; si la P de<br />
O 2 aumenta los miocitos se contraen. Ello contribuye a la distribución homogénea del aire inspirado.<br />
La inervación de los bronquiolos es idéntica a la de los bronquios: el efecto bronco constrictor es más notable debido<br />
a la importancia de la capa muscular.<br />
El bronquiolo ya no presenta cartílago, pero si capa adventicia. Según avanzamos en los bronquiolos, disminuye la<br />
altura del epitelio y aumenta la cantidad de musculo y de células de Clara.<br />
EPOC<br />
La enfermedad pulmonar obstructiva cronica (EPOC) incluye enfisema y asma.<br />
El enfisema es una patología en la cual un estímulo persistente (por ejemplo, el tabaco) provoca una atracción sobre<br />
neutrófilos y macrófagos, que llegan a la lámina propia bronquiolar y liberan elastasa, una proteína que rompe las<br />
fibras elásticas. Además, disminuye la secreción de P-antitripsina, otra proteína que neutraliza la elastasa. Esto lleva<br />
a un colapso bronquiolar durante la espiración y una obstrucción del flujo del aire.<br />
El asma es una inflamación de las vías respiratorias por aumento del número de linfocitos Tc y eosinófilos,<br />
hipersecreción de las glándulas y aumento de la permeabilidad vascular, que produce edema. Se produce asimismo<br />
una bronco-constricción exagerada, produciendo disnea.<br />
Bronquiolo terminal<br />
El bronquiolo terminal es la última de las vías conductoras. Presenta un epitelio cúbico bajo, con células de Clara y<br />
células ciliadas, pero sin células caliciformes. Se observan haces de fibras elásticas dispuestas longitudinalmente y<br />
miocitos lisos dispuestos en espiral, inervados por el SN Autónomo en su división parasimpática. El bronquiolo<br />
terminal se ramifica dando lugar al bronquiolo respiratorio.<br />
Bronquiolo respiratorio<br />
El bronquiolo respiratorio tiene un diámetro inferior a 0.2 mm y se encuentran en número de 130000.<br />
Su pared está interrumpida por la presencia de alveolos. La pared bronquiolar muestra aquí un epitelio cúbico con<br />
células ciliadas, si bien la altura y el número de cilios van decreciendo. También aparecen células de Kulchitsky y<br />
células con ribete en cepillo de manera ocasional.<br />
También encontramos miocitos lisos dispersos, que ya no forman una capa continua. Se ramifica hasta 3 veces para<br />
dar paso a los conductos alveolares.<br />
Conducto alveolar<br />
Este conducto tiene, asociada a su pared, una enorme cantidad de alveolos, de modo que tiene <strong>com</strong>o pared solo los<br />
bordes libres de los tabiques interalveolares. Se ramifica 2 o 3 veces para dar lugar al atrio o saco alveolar, vestíbulo<br />
del que se abren numerosos alveolos.
Página30<br />
ALVEOLOS<br />
Generalidades<br />
El alveolo es la unidad funcional del pulmón, en un número aproximado de unos 100-300 millones. Es un espacio<br />
aéreo en forma de saco de forma poligonal y pared delgada de 0,2mm de diámetro que se abre a los bronquiolos<br />
respiratorios a través de un conducto alveolar y un saco alveolar.<br />
Un saco alveolar es un espacio rodado por cúmulos de alveolos.<br />
En la pared alveolar encontramos los poros de Kohn, con un diámetro de 8μm, que permiten que el aire se<br />
<strong>com</strong>unique con los alveolos adyacentes, dando lugar a la circulación colateral del aire. A través de estos poros<br />
pueden diseminar microorganismos.<br />
Embriología<br />
Los alveolos se desarrollan entre los meses 4 y 6, siendo espacios llenos de líquido amniótico. En el feto son<br />
redondeados (aún no han adquirido su forma poligonal), pero antes de nacer son más poliédricos y presentan redes<br />
capilares en el interior de sus paredes.<br />
En el recién nacido hay 20 millones de alveolos que se llenan de aire gracias a los movimientos respiratorios que<br />
dilatan los espacios y mejoran el aporte sanguíneo a las paredes alveolares. Los alveolos crecen en número y tamaño<br />
durante la infancia, llegando a los 300 millones a los 8 años.<br />
Epitelio Alveolar<br />
El alveolo esta tapizado por un epitelio simple plano formado por neumocitos tipo I y II, bajo el hay una lámina basal<br />
y bajo ella un intersticio de tejido conjuntivo.<br />
El epitelio tiene muchas funciones, todas muy activas e importantes en los aproximadamente 72m 2 de extensión<br />
total que tiene:<br />
<br />
<br />
<br />
Intercambio Gaseoso Pulmón-Aire<br />
Metabolismo y transformación de la angiotensina I en II por acción de la enzima ECA.<br />
Degrada moléculas circulantes en la sangre, <strong>com</strong>o la serotonina o la bradiquinina.<br />
En los alveolos encontramos una capa de surfactante, una secreción grasa que evita el colapso alveolar. Es un agente<br />
tensoactivo sintetizado por los neumocitos tipo II.<br />
NEUMOCITOS TIPO I<br />
Los neumocitos tipo I suponen un 95% de la superficie del alveolo y un 40% del número de células epiteliales.<br />
Son células aplanadas, con un grosor de 0,2 μm y escasos orgánulos. Están unidos mediante uniones estrechas entre<br />
sí y con neumocitos tipo II. No tienen capacidad de división.<br />
NEUMOCITOS TIPO II<br />
Los neumocitos tipo II suponen un 60% del total de células del epitelio alveolar.<br />
Son células de morfología cubica o redondeada de modo que sobresalen sobre el resto del epitelio (plano simple). Se<br />
sitúan sobre todo en los ángulos de los tabiques alveolares<br />
Presentan microvellosidades apicales cortas, un núcleo basófilo y redondeado, así <strong>com</strong>o abundancia de RER, REL y<br />
aparato de Golgi, puesto que son células muy activas sintéticamente, así <strong>com</strong>o mitocondrias. Presentan en la<br />
superficie apical está llena de orgánulos que se organizan <strong>com</strong>o pilas membranosas paralelas llamadas cuerpos<br />
laminares, que contienen el surfactante.
Página31<br />
Surfactante<br />
Componente lipídico<br />
El surfactante es una sustancia lipoproteica, con una riqueza del 90% en lípidos.<br />
Está <strong>com</strong>puesto principalmente de colesterol y otra serie de fosfolípidos: dipalmitoil fosfatidilcolina (DPPC),<br />
fosfatidilcolina, fosfatidilglicerol, fosfatidilinositol, fosfatidilserina, fosfatidiletanolamina, esfingomileina y<br />
glucolípidos.<br />
Componente proteico<br />
La parte proteica del surfactante la <strong>com</strong>ponen tres proteínas principales: SP-A, SP-B, SP-C y SP-D. Estas proteínas son<br />
importantes en el metabolismo y la función inmunológica del surfactante.<br />
SP-A forma parte del surfactante y es decisiva en formar una estructura de mielina tubular. Modula las respuestas<br />
inmunitarias frente a virus, bacterias y hongos. Promueve fagocitosis, opsoniza y regula la producción de citosinas.<br />
SP-B está implicada en la maduración de la capa de surfactante. Participa en la adsorción de lípidos y extiende la<br />
capa sobre la superficie alveolar. Participa en la lisis del surfactante.<br />
SP-C estabiliza la monocapa de surfactante.<br />
SP-D estabiliza el fosfatidilinositol. Tiene función inmunológica en respuestas alérgicas e inflamatorias. Participa en<br />
opsonización, interactuando con SP-A fuera de la célula.<br />
Las proteínas del surfactante se sintetizan en el neumocito tipo II y se liberan por exocitosis.<br />
SP-B y SP-C, ya fuera de la celula, interactúan con SP-A e iones Ca2+ para formar una monocapa por interacción con<br />
DPPC.<br />
Tejido Intersticial<br />
El intersticio es un tejido conjuntivo delgado con abundantes fibroblastos, leucocitos y mastocitos. Encontramos en<br />
él fibras colágenas y reticulares de sostén, así <strong>com</strong>o fibras elásticas que permiten la expansión de la pared alveolar en<br />
inspiración y contribuyen a la espiración, evitando el colapso bronquiolar y alveolar. Se destruyen en enfisema, lo que<br />
provoca una destrucción de las paredes alveolares y un incremento de los espacios aéreos.<br />
MACRÓFAGOS ALVEOLARES<br />
Los macrófagos alveolares proceden de monocitos y se encuentran dentro del intersticio o en la superficie alveolar.<br />
Son células libres y móviles capaces de atravesar el epitelio, situadas en la superficie alveolar. Entonces migran hacia<br />
la tráquea gracias a su capacidad móvil y el movimiento de la capa de moco batida por los cilios. Se eliminan por tos y<br />
deglución a un ritmo de 2 millones por hora. Se piensa que también tienen capacidad de retornar al intersticio,<br />
puesto que aparecen en la pleura y la pared de bronquios cargados de carbón que ha sido inhalado por las vías<br />
superiores.<br />
Los macrófagos alveolares tienen una alta tasa de recambio. Sus marcadores son CD68 y CD74 y presentan un núcleo<br />
irregular con nucléolo marcado. Hay un alto desarrollo del aparato de Golgi, mitocondrias y ribosomas, e inclusiones<br />
de glucógeno y lisosomas heterogéneos.<br />
Los macrófagos alveolares tienen una función defensiva frente a restos celulares en el intersticio, polvo, polen,<br />
tóxicos y microorganismos del aire, manteniendo la superficie alveolar estéril. Si reconoce un antígeno o toxina,<br />
segrega quimiocinas que atraen a los leucocitos hasta el epitelio alveolar.<br />
También fagocitan a los eritrocitos que puedan entrar en el pulmón por insuficiencia cardíaca, de modo que se puede<br />
percibir en estos casos hemosiderina. Existe la posibilidad de que el macrófago fagocite el bacilo de la tuberculosis<br />
(Mycobacterium Tuberculosis), que no es digerido por la célula, si no que permanece retenido en la célula y en caso<br />
de rotura macrocítica se produce recurrencia de la enfermedad.
Página32<br />
Vascularización<br />
En el intersticio no encontramos vasos linfáticos, pero sí una extensa vascularización de redes capilares de endotelio<br />
continuo.<br />
En la red de capilares es donde se produce el intercambio de gases por difusión pasiva a través de la barrera de<br />
difusión.<br />
La barrera de difusión es una barrera muy delgada (0.2 μm) <strong>com</strong>puesta, desde la luz hacia la profundidad, de<br />
surfactante, epitelio alveolar, laminas basales alveolar y capilar fusionadas y un endotelio. Es aquí donde se produce<br />
la mayor parte del intercambio hemato-gaseoso.<br />
Existe una parte más gruesa, la barrera de difusión gruesa, donde el intercambio se da en menor medida. En esta<br />
zona las láminas basales del alveolo y el capilar no están fusionadas porque entre ambas media un intersticio, que<br />
puede acumular líquido que no puede ser drenado (no hay vasos linfáticos), por lo que circula libremente hasta<br />
llegar a los capilares linfáticos próximos, en los bronquiolos terminales.<br />
La pared alveolar presenta, pues, dos zonas funcionales.<br />
El líquido acumulado no interfiere con el intercambio, salvo en situaciones alteradas <strong>com</strong>o el edema pulmonar por<br />
aumento de presión y salida del plasma hacia intersticio y luz alveolar. Esto sucede frecuentemente en insuficiencia<br />
ventricular izquierda.<br />
A través de la red de capilares se pierden de 800 ml de agua al día.<br />
PLEURA<br />
La cara interna de la caja torácica y la externa de los pulmones está recubierta por serosas que conforman la pleura,<br />
con sus dos hojas: parietal y visceral. Entre ambas queda un espacio pleural lleno de líquido que actúa <strong>com</strong>o<br />
lubricante.<br />
La pleura parietal está formada por tejido adiposo, que se asienta directamente bajo la serosa, y un conjuntivo<br />
fibroso vascularizado e inervado, que se asienta sobre el periostio o el perimisio.<br />
La pleura visceral está formada por un mesotelio con microvellosidads apicales cortas y un conjuntivo dividido en dos<br />
zonas separadas por una membrana elástica externa. Bajo el conjuntivo encontramos la membrana elástica interna y<br />
después el intersticio alveolar.<br />
La pleura visceral sella la superficie pulmonar evitando la fuga de aire en caso de que se produzca una rotura.<br />
Si por cualquier motivo saliese aire de los pulmones a la cavidad torácica, se produce un neumotórax. Pueden<br />
producirse también el vertido de agua (hidrotórax), sangre (hemotórax) o alimentos (quilotórax).<br />
El mesotelio puede maliginizarse dando lugar a un mesiotelioma por exposicion a amianto.
Página33<br />
SISTEMA DIGESTIVO<br />
El aparato digestivo está formado por el tubo digestivo y sus glándulas anexas: lengua, dientes, glándulas salivales,<br />
páncreas, hígado y vesícula biliar.<br />
Cumple varias funciones en el organismo:<br />
<br />
<br />
<br />
<br />
Función absortiva de sustratos metabólicos<br />
Función de barrera contra sustancias nocivas, antígenos y microrganismos patógenos.<br />
Función inmunológica ante las sustancias nocivas que hayan atravesado la mucosa.<br />
Función secretora de moléculas de protección del tubo digestivo o destinadas a la digestión, entre otras.<br />
CAVIDAD ORAL<br />
Vestíbulo<br />
El vestíbulo es la parte de la cavidad oral delimitada, anteriormente, por los labios, lateralmente, por las mejillas, y<br />
posteriormente, por las encías y los dientes.<br />
En él drenan las glándulas salivales labiales, yugales, molares y las parótidas.<br />
Cavidad Oral<br />
La cavidad oral propiamente dicha está limitada anteriormente por los dientes y las encías, superiormente por el<br />
paladar duro y blando, caudalmente por el piso de la boca y la lengua y posteriormente, por el istmo de las fauces.<br />
En el suelo de la cavidad oral se abren los conductos excretores de las glándulas salivales sublinguales y<br />
submaxilares.<br />
En la cavidad oral se distinguen tres áreas histológicas: la mucosa masticatoria, la mucosa no masticatoria y la<br />
mucosa especializada.<br />
Mucosa Masticatoria<br />
Esta mucosa presenta un epitelio plano estratificado queratinizado, con cuatro estratos y sin estrato lúcido. Aparece<br />
en el paladar duro y en las encías, donde es posible encontrar zonas en donde este epitelio esté paraqueratinizado<br />
(similar al queratinizado, pero las células del estrato córneo no pierden sus núcleos).<br />
Debajo del epitelio, encontramos una lámina propia, similar a la dermis, con una zona papilar y otra reticular. Las<br />
crestas epiteliales y las papilas dérmicas están muy marcadas. Las papilas presentan muchos capilares, terminaciones<br />
nerviosas y corpúsculos.<br />
Esta mucosa está adherida directamente al hueso, por lo que presenta una mayor resistencia frente a las tracciones.<br />
No es una mucosa elástica y es más espesa que la mucosa no masticatoria.<br />
Encías<br />
En las encías, la mucosa masticatoria está adherida al diente o al hueso alveolar, es decir, no hay submucosa bajo la<br />
lámina propia, que además carece de glándulas.<br />
El epitelio de la mucosa de las encías puede ser de fijación o cervicular:<br />
En el epitelio de fijación, parte de la mucosa está unida al diente, permitiendo sellarlo; cuando se rompe, favorece la<br />
entrada de gérmenes y la aparición de gingivitis (inflamación de la encía).<br />
Los dientes están insertos en una zona excavada, el alveolo dentario, de modo que existe un epitelio que tapiza esta<br />
cavidad, conocido <strong>com</strong>o epitelio cervicular. Es un epitelio no queratinizado y más fino que el del resto de la mucosa<br />
de la encía.
Página34<br />
Paladar Duro<br />
El paladar duro conforma la parte anterior del techo de la cavidad oral, y se encuentra dividido por el rafe medio en<br />
dos mitades, izquierda y derecha, simétricas.<br />
La mucosa masticatoria del paladar duro sí presenta submucosa, excepto en el rafe. Esta submucosa es un tejido<br />
conjuntivo fibroso de haces colágenos, con la misión de unir la mucosa masticatoria con el colágeno asociado al<br />
hueso (fibras de Sharpey). En la submucosa encontramos adipocitos, sobre todo en la parte anterior del paladar<br />
duro. En la parte posterior, esta submucosa presenta glándulas mucosas.<br />
Mucosa de Revestimiento<br />
La mucosa de revestimiento es una mucosa más fina que la masticatoria. Presenta un epitelio plano estratificado no<br />
queratinizado, con tres estratos (basal, espinoso y superficial). En el borde libre del labio, este epitelio puede<br />
queratinizarse.<br />
La lámina propia suele ser rica en fibras elásticas, así <strong>com</strong>o en nervios y fibras nerviosas. Esto es sumamente<br />
importante, porque gracias a ello la mucosa no masticatoria presenta cierta movilidad y adaptación a los<br />
movimientos de la cavidad oral. El hecho de que el epitelio sea más fino contribuye además a que sea algo más<br />
permeable, en distintos grados, habiendo zonas más permeables <strong>com</strong>o el suelo de la boca.<br />
Este epitelio aparece en los labios, las mejillas, la superficie mucosa alveolar, el piso de la boca, las superficies<br />
inferiores de la lengua y el paladar blanco.<br />
Su lámina propia puede descansar sobre periostio, músculo estriado o glándulas.<br />
Labio<br />
Histológicamente, el labio presenta tres zonas: una externa o cutánea, una interna o mucosa oral, y otra roja o del<br />
bermellón.<br />
La zona externa o cutánea es piel con glándulas sebáceas, sudoríparas y folículos pilosos.<br />
La zona interna o mucosa oral está tapizada por epitelio plano estratificado y presenta glándulas salivales menores<br />
en su lámina propia, bajo la cual encontramos los músculos orbiculares de los labios.<br />
La zona roja o del bermellón se trata de piel, con células epiteliales a modo de gránulos eosinófilos de eleidina,<br />
haciendo que sean células translúcidas. Presenta glándulas sebáceas (gránulos de Fordyce*). Este epitelio está<br />
anclado a su lámina propia, que presenta un firme irregular y papilas muy vascularizadas, lo que dota a esta zona de<br />
su color rojo característico. No presenta glándulas sudoríparas (se seca con facilidad) ni folículos pilosos.<br />
*Gránulos de Fordyce<br />
Las gránulos de Fordyce son glándulas sebáceas especializadas no asociadas a folículo piloso cuyas células poseen<br />
unos gránulos llamados gránulos de Fordyce.<br />
Mejillas<br />
Las mejillas presenta, bajo su mucosa no masticatoria, una submucosa rica en fibras colágenas y elásticas que evitan<br />
que la mucosa se pliegue, ya que le aplican cierta tensión. En la submucosa encontramos glándulas salivales (yugales<br />
y molares) y glándulas sebáceas, de tamaño considerable y visibles macroscópicamente (con granos de Fordyce), en<br />
la parte posterior de la mejilla. Estas glándulas sebáceas no están asociadas a folículos pilosos.<br />
La submucosa se asienta sobre el músculo buccinador.<br />
Paladar Blando<br />
El paladar blando conforma la parte posterior del techo de la cavidad oral. Es móvil debido a la ausencia de hueso.<br />
Termina en una extensión posterior conocida <strong>com</strong>o la úvula.<br />
El paladar blando presenta gran cantidad de papilas en su mucosa. En la zona nasal, presenta mucosa respiratoria.<br />
Bajo la mucosa no masticatoria de la zona oral hay una submucosa con glándulas mucosas y una capa muscular.
Página35<br />
LENGUA<br />
La lengua es un órgano muscular esquelético, con abundantes miocitos agrupados en haces orientados en todas las<br />
direcciones del espacio, que otorgan a la lengua una gran movilidad y flexibilidad. Esta musculatura se divide en<br />
extrínseca e intrínseca: la musculatura extrínseca la <strong>com</strong>ponen miocitos que se insertan fuera de la lengua y son<br />
responsables de mover la lengua, mientras que la musculatura intrínseca corresponde a miocitos que se insertan en<br />
la propia lengua y permiten cambiar la forma de la misma.<br />
La cara ventral de la lengua está tapizada por mucosa no masticatoria, fina y sin submucosa. La cara dorsal está<br />
tapizada por mucosa especializada. Podemos dividir esta superficie en tres tercios: los dos tercios anteriores<br />
presentan abundantes papilas linguales y se separan, mediante el surco terminal del tercio posterior (raíz o base)<br />
donde encontramos las amígdalas linguales. El surco terminal, con forma de V, divide la parte anterior de la raíz y<br />
contiene el foramen ciego, que corresponde a los restos del conducto tirogloso.<br />
Presentan en la parte posterior de la lengua un tejido linfoide asociado: la amígdala lingual.<br />
Papilas<br />
Las papilas son irregulares elevaciones de la mucosa dorsal (dos tercios anteriores) formadas por un núcleo central<br />
conjuntivo cubierto por un epitelio plano estratificado. Se asocian con los botones gustativos formando la mucosa<br />
especializada de la lengua.<br />
Existen cuatro tipos de papilas: filiformes, fungiformes, foliadas y caliciformes.<br />
Papilas Filiformes<br />
Las papilas filiformes son las más abundantes, y se encuentran por toda la parte anterior de la lengua. Habitualmente<br />
se disponen formando hileras, dispuestas paralelamente al surco terminal.<br />
Cada papila filiforme es una proyección de forma cónica. La punta del cono está orientada hacia la base de la lengua.<br />
Tienen un eje conjuntivo y un epitelio plano estratificado muy queratinizado. Al óptico, se ve tan solo una mínima<br />
parte de la proyección porque se pierde la mayor parte al hacer la preparación.<br />
Estas papilas están muy queratinizadas para poder erosionar el alimento, es decir, tienen una función eminemente<br />
mecánica, por lo que no presentan botones gustativos.<br />
Algunos trastornos provocan la alteración de la capa superior de la lengua, que se cubre por una sustancia pastosa<br />
que impide renovar las papilas; esto se conoce <strong>com</strong>o lengua saburral.<br />
Papilas Fungiformes<br />
Las papilas fungiformes se encuentran en menor número y se disponen intercaladas entre las hileras de papilas<br />
filiformes, aunque abundan en la punta de la lengua. Se aprecian a simple vista y tienen un color rosado.<br />
Son proyecciones en forma de hongo, con un eje de conjuntivo recubierto por epitelio plano estratificado no<br />
queratinizado. El eje conjuntivo está muy vascularizado, lo que hace que no se distingan entre ellas, pero sí de las<br />
demás por su tono rojo.<br />
Tienen una función sensitiva, ya que en ellas encontramos botones gustativos.<br />
Papilas Foliadas<br />
Estas papilas son poco abundantes en nuestra especie, si bien son más visibles en niños. Se sitúan en los bordes<br />
laterales de la cara dorsal de la lengua dispuestas <strong>com</strong>o crestas bajas alineadas paralelas entre sí, perpendiculares al<br />
eje mayor de la lengua.<br />
Presentan un eje conjuntivo cubierto por epitelio plano estratificado no queratinizado, con botones gustativos. Están<br />
separadas entre sí por profundas hendiduras donde drenan glándulas serosas.<br />
Papilas Caliciformes o Circunvaladas<br />
Son entre 8 y 12 papilas alineadas justo por delante del surco terminal. Son redondeadas y sumamente grandes, de<br />
modo que son visibles a simple vista.
Página36<br />
Cada papila está alojada en una depresión de la mucosa. El espacio entre la papila y la depresión es el denominado<br />
surco o valle, al fondo del cual se abren los conductos excretores de las glándulas de von Ebner, que permiten que<br />
los botones gustativos respondan rápidamente a estímulos cambiantes.<br />
La papila muestra un eje conjuntivo muy vascularizado y un epitelio plano estratificado que puede queratinizar. En<br />
los bordes de la papila encontramos abundantes botones gustativos.<br />
Botones Gustativos<br />
Son estructuras ovaladas, pálidas, en forma de tonel, ocupando todo el espesor del epitelio en un número de 3000<br />
asociadas a papilas fungiformes, foliadas y caliciformes, así <strong>com</strong>o fuera de la lengua (arco glosopalatino, paladar<br />
blando, superficie posterior de la epiglotis, parte posterior de la faringe)<br />
A MO se distingue en ellos un poro, una fosita y 3 tipos celulares: basales, de sostén (oscuras) y neuroepiteliales<br />
(claras). Las de sostén se disponen en la periferia y las sensoriales en el centro del botón.<br />
La superficie del epitelio del botón tiene una zona hundida, la fosita, donde se observan los pelos gustativos,<br />
inmersos en una matriz amorfa eosinófila.<br />
A MET se establece que hay 4 tipos celulares:<br />
· Tipo I: corresponden a las células oscuras. Son periféricas, de<br />
carácter secretor, por lo que presentan gránulos densos que, cuando<br />
se vierten por exocitosis, permiten liberar el material amorfo.<br />
Presentan abundante RER, mitocondrias y aparato de Golgi, así <strong>com</strong>o<br />
microvellosidades<br />
· Tipo II: corresponden a parte de las células claras. Son células de<br />
soporte, sin gránulos densos, con microvellosidades y REL. Algunos<br />
autores defienden que son células tipo I en un estado funcional<br />
distinto.<br />
· Tipo III: corresponden a otra parte de las células claras. Son los<br />
receptores sensitivos propiamente dichos, es decir, son neuronas con<br />
una prolongación apical que invade el material amorfo y termina en<br />
una expansión o maza que sobresale sobre esa matriz amorfa. En su<br />
base, se aprecian fibras nerviosas de naturaleza sensitiva, que<br />
establecen sinapsis con la zona basal de las células tipo III.<br />
· Tipo IV: corresponden a las células basales. Regeneran y renuevan el<br />
botón<br />
Los botones gustativos son entonces órganos sensitivos dentro del epitelio de la lengua, que se encargan de percibir<br />
sabores, y nos advierten si el alimento es digestible.<br />
Además, cuando los botones gustativos se ponen en marcha, se producen unos fenómenos que tienen que ver con la<br />
secreción de la saliva y del jugo gástrico. Están inervados por las fibras de los pares craneales VII, IX y X.<br />
Los botones gustativos perciben 5 sabores, y todos los botones pueden percibir todos los sabores. Estos cinco<br />
sabores son dulce, salado, amargo, ácido y umami. El umami es un sabor carnoso que se debe a los glutamatos,<br />
presentes en carnes, sopas, conservas…. Para poder saborear las sustancias, éstas deben estar disueltas en agua.<br />
Existen dos tipos distintos de receptores en los botones gustativos, TR1 y TR2, ambos acoplados a gustductina, una<br />
proteína G. Cuando una sustancia es detectada por este receptor, se desencadena la apertura o cierre de canales de<br />
Na+ y Ca2+, lo que puede hiperpolarizar o despolarizar la célula. En este último caso es cuando se produce el impulso<br />
nervioso.<br />
Glándulas Linguales<br />
Aparecen:<br />
Glándulas mucosas, que se abren al fondo de las criptas de las amígdalas linguales.<br />
Glándulas de Nuhn o de Blandin: glándulas mixtas situadas en la zona más proximal de la lengua; drenan a la<br />
cara ventral de la lengua.<br />
Glándulas de von Ebner: glándulas serosas cuyos conductos drenan en el fondo del valle de papilas<br />
caliciformes.
Página37<br />
DIENTE<br />
Los dientes son estructuras esenciales en el proceso digestivo que se insertan en los maxilares superior e<br />
inferior. En la infancia hay un total de 20 dientes deciduos, pero en el adulto el número aumenta a los 32<br />
dientes permanentes.<br />
Los dientes se pueden dividir histológicamente en varios <strong>com</strong>ponentes estructurales: esmalte y cemento (parte<br />
externa), dentina (inferior) y pulpa (capa más interna).<br />
Anatómicamente se pueden dividir en 3 zonas:<br />
<br />
<br />
<br />
La corona anatómica es la zona recubierta por<br />
esmalte; de todos modos también se puede<br />
diferenciar una corona clínica <strong>com</strong>o aquella<br />
expuesta por encima de la línea gingival.<br />
Corona = Esmalte + Dentina<br />
El cuello o cérvix es la unión de la corona con la<br />
raíz.<br />
La raíz es la parte del diente cubierta por el<br />
cemento. Pueden aparecer de 1 a 3, según el<br />
diente.<br />
Raíz = Cemento + Dentina<br />
Esmalte<br />
El esmalte es la sustancia más dura del organismo, <strong>com</strong>puesta en un 96-98% por hidroxiapatita, por lo que<br />
apenas presenta material orgánico.<br />
La hidroxiapatita se dispone en forma de bastoncillos o prismas con un diámetro de entre 4-8μm dispuestos a<br />
lo largo de toda la capa desde la unión amelodentinal hasta la superficie del esmalte. Los bastoncillos forman<br />
unas estriaciones llamadas líneas de contorno de Retzius, en los que se evidencia el ritmo del crecimiento. Una<br />
vez el esmalte es maduro, es acelular y no se reemplaza.<br />
De todos modos, no es un tejido estático, porque sufre la acción de saliva y secreciones que las mantienen<br />
(enzimas digestivas, enzimas antibacterianas, anticuerpos y <strong>com</strong>ponentes inorgánicos, <strong>com</strong>o minerales).<br />
Formación del Esmalte o Amelogénesis<br />
En un primer momento se forma la dentina y, sobre esta, el esmalte. Esta síntesis tiene dos etapas:<br />
Etapa secretora: los ameloblastos secretores producen una matriz orgánica proteinácea por acción del RE, el<br />
aparato de Golgi y los gránulos de secreción. Consiguen formar el espesor total del esmalte . Los ameloblastos<br />
secretores son células cilíndricas estrechas y muy polarizadas que se encuentran inmediatamente adyacentes al<br />
esmalte en formación. En el polo apical de la célula presentan una prolongación llamada proceso de Tomes. Su<br />
polo basal es muy eosinófilo por la alta concentración de mitocondrias que presentan, al ig ual que en la zona<br />
apical, <strong>com</strong>plejos de unión asociados a filamentos contráctiles que permiten el movimiento de la célula a<br />
través del esmalte en formación.<br />
Etapa madurativa: la maduración del esmalte, realizada por los ameloblastos maduros, consiste en la retirada<br />
de la matriz orgánica y el agregado constante de Ca y fosfato a esta. Los ameloblastos maduros se diferencian a<br />
partir de los ameloblastos secretores y se pueden diferenciar en ameloblastos madurativos de borde estriado<br />
(70%) y de superficie lisa (30%). Entre los ameloblastos madurativos aparecen las células papilares estrelladas,,<br />
que, al igual que los ameloblastos madurativos, tienen una gran cantidad de mitocondri as.
Página38<br />
La matriz del esmalte tiene, inicialmente, amelogeninas, enamelaninas y P de los penachos o flecos ( tuft<br />
protein), pero en el esmalte maduro sólo aparecen enamelinas y P de los penachos (presente en la unión<br />
amelodentinal, presenta un porcentaje mayor que en el resto del esmalte de matriz orgánica).<br />
Cemento<br />
El cemento recubre la raíz del diente y permite la inserción con el alveolo del maxilar. Es una fina capa de<br />
material similar al hueso secretada por los cementocitos (muy similares a los osteocitos).<br />
El cemento tiene una <strong>com</strong>posición de un 45-50% de matriz mineral, y aparecen unas lagunas o canalículos en él<br />
que alojan a los cementocitos y sus prolongaciones. Se diferencia del hueso en que el cemento es avascular y<br />
en que los canalículos no se inter<strong>com</strong>unican.<br />
Junto a los cementocitos aparecen en la parte externa del cemento una capa de cementoblastos, adyacentes al<br />
ligamento periodóntico.<br />
Dentina<br />
La dentina es un material calcificado que <strong>com</strong>pone la mayor parte de la sustancia del diente, entre el esmalte y<br />
el cemento, por fuera, y la pulpa, por dentro.<br />
La dentina está formada por un 70% de hidroxiapatita y es secretada por los odontoblastos, que forman una<br />
capa epitelial por encima de la superficie interna de la dentina (cara en contacto con la pulpa).<br />
Los odontoblastos son células cilíndricas alargadas que contienen un RER bien desarrollado, un gran aparato de<br />
Golgi y otras organelas relacionadas con la síntesis y secreción de P. La superficie apical del odontoblasto está<br />
en contacto con la dentina en formación y se separa de la pulpa por <strong>com</strong>plejos de unión entre los<br />
odontoblastos.<br />
La capa de odontoblastos se retrae a medida que se deposita la dentina, de modo que quedan las<br />
prolongaciones de los odontoblastos apresadas en delgados canalículos llamados túbulos dentinales. El<br />
conjunto de los túbulos y sus prolongaciones forman engrosamientos en la dentina que originan unas líneas de<br />
crecimiento, <strong>com</strong>o las líneas incrementales de von Ebner y las líneas de Owen, más gruesas). Estas líneas<br />
evidencian el crecimiento del diente; hoy en día se usan en la Medicina Forense.<br />
Los odontoblastos proceden de las células subodontoblásticas, que en la dentinogénesis forman unos haces de<br />
fibras colágenas llamadas fibras de von Korff.<br />
La predentina es la matriz orgánica recién sintetizada, por lo que aparece al lado del odontoblasto y no está<br />
mineralizada. En los odontoblastos aparecen los cuerpos en ábaco, unos gránulos que se van condensando<br />
según maduran a gránulos de secreción, y que contienen colágeno, hidroxiapatita y un precursor de colágeno<br />
que se dispone en forma filamentosa y permite la unión a Ca.<br />
Pulpa<br />
La cavidad pulpar es un espacio dentario con la forma del diente correspondiente ocupada por la pulpa, un<br />
tejido conjuntivo laxo con rica vascularización e intervación. Los vasos sanguíneos y los nervios ingresan por el<br />
orificio radicular o apical, y se extienden hasta la corona del diente donde forman redes vasculares y nerviosas<br />
inferiores a la capa de odontoblastos. Algunas fibras nerviosas desnudas también penetran en las porciones<br />
proximales de los túbulos dentinales y hacen contacto con las prolongaciones de los odontoblastos.<br />
Proceso alveolar y Hueso Alveolar<br />
El hueso alveolar es una fina capa de hueso <strong>com</strong>pacto que forma la pared del alveolo; permite la inserción del<br />
ligamento periodóntico
Página39<br />
Ligamento Periodóntico<br />
El ligamento periodóntico es el tejido conectivo fibroso que une al diente con el hueso con lo rodea, aunque<br />
también permite la remodelación ósea, la nutrición de estructuras adyacentes, la propiocepción y la erupción<br />
dentaria.<br />
Se encuentra formado por zonas un tejido conjuntivo laxo y otras de tejido conjuntivo denso,que en ambos<br />
casos constan de fibras colágenas y fibroblastos de aspecto alargado y paralelos al eje mayor de las fibras<br />
colágenas. Los fibroblastos se mueven hacia adelante y atrás sintentizando colágeno, pero también lo fagocitan<br />
y degradan por enzimas colagenasas. Además, en el tejido conjuntivo laxo contiene vasos sanguíneos y<br />
terminaciones nerviosas.<br />
De forma longitudinal se pueden apreciar, junto con el resto de <strong>com</strong>ponentes, fibras de oxitalán, que se unen al<br />
hueso y al cemento en ambos extremos; histológicamente son parecidas a las fibras elásticas y permiten<br />
movimientos ligeros del diente dentro de la cavidad alveolar.<br />
Fibras de Sharpey<br />
Se extienden desde el ligamento periodóntico hasta el cemento, y consisten en unas fibras colágenas, que en la<br />
porción que se adentra en el cemento está mineralizada.<br />
GLÁNDULAS SALIVALES<br />
Las principales glándulas salivales, cada una en un número par, son: parótida, submaxilar y sublingual.<br />
Las glándulas parótidas y submaxilares están localizadas por fuera de la cavidad oral, por lo que transportan<br />
sus secreciones a través de largos conductos.<br />
Las glándulas salivales menores o accesorias se ubican en la submucosa de distintas partes de la cavidad oral;<br />
son las glándulas linguales, labiales, bucales, molares y palatinas.<br />
El origen de las glándulas salivales está en un epitelio oral embrionario que <strong>com</strong>ienza siendo un cordón celular<br />
macizo que penetra en el mesénquima, que poco a poco se va ramificando y ensanchando en sus extremos.<br />
Finalmente se degeneran las células del interior del cordón celular y las ramificaciones ensanchadas forman lso<br />
acinos secretores.<br />
Cada glándula se puede dividir en dos partes: acino y conducto excretor.<br />
Cápsula<br />
Las principales glándulas salivales principales están rodeadas por una cápsula de tejido conectivo<br />
moderadamente denso que emite tabiques a la porción secretora o adenómero, que queda dividida en lóbulos<br />
y lobulillos. Los tabiques contienen el paquete vasculonervioso así <strong>com</strong>o los conductos de excreción. En las<br />
glándulas salivales menores no hay una cápsula <strong>com</strong>pleta, pero sí aparece tejido conectivo que rodea al<br />
parénquima y que, al igual que en las principales, contiene linfocitos y plasmocitos que permiten la formación<br />
de anticuerpos salivales.<br />
Acino<br />
Los acinos de las glándulas salivales pueden contener células mucosas, serosas o ambas; así hay acinos<br />
mucosos, serosos o mixtos.<br />
Por lo general, los acinos serosos son esféricos, mientras que los mucosos son tubulares. Los acinos mixtos son<br />
acinos mucosos con un casquete de células serosas que forman una semiluna serosa.
Página40<br />
CÉLULAS SEROSAS<br />
Las células serosas contienen una gran cantidad de RER, ribosomas libres, un gran desarrollo del aparato de<br />
Golgi y abundantes gránulos de secreción que contienen cimógeno y se almacenan en el dominio apical; las<br />
organelas, en cambio, se localizan a nivel perinuclear. Así, la zona apical es eosinófila por la presencia del<br />
cimógeno, mientras que la zona basal es basófila.<br />
La base de la célula serosa puede mostrar invaginaciones de la membrana plasmática y pliegues baso-laterales<br />
que se interdigitan con las células vecinas. Se unen por el dominio apical por <strong>com</strong>plejos de unión, a menos que<br />
la luz del canalículo se extienda entre células vecinas a modo de canalículo tortuoso y tortuoso, de modo que<br />
el <strong>com</strong>plejo de unión se localiza al fondo del canalículo.<br />
CÉLULAS MUCOSAS<br />
Esquema Células Serosas<br />
Las células mucosas tienen gránulos de mucinógeno que no se tiñen con HE (con esta tinción aparece en la<br />
zona apical espacios sin teñir que corresponden a estos gránulos), pero contienen muchos hidratos de carbono<br />
por lo que son PAS+. Estas células responden a ciclos determinados a un estímulo hormonal o neural por el que<br />
secretan todos los gránulos, tras el que sintetizan nuevos gránulos y almacenan en la zona apical.<br />
Estas células tienen <strong>com</strong>plejos de unión apicales, más <strong>com</strong>unes en acinos mixtos mucosos con semilunas<br />
serosas.<br />
CÉLULAS MIOEPITELIALES<br />
Las células mioepiteliales son células contráctiles con numerosas prolongaciones y filamentos de actina y<br />
miosina que se tiñen con eosina. Estas células se disponen entre la membrana plasmática basal de las células<br />
epiteliales y la lámina basal del epitelio, aunque también aparecen en la unión del acino con el conducto<br />
excretor.<br />
Sistema de Conductos<br />
La luz de los acinos salivales se continúa con la de un sistema de conductos que puede tener hasta tres<br />
segmentos: conductos intercalares, estriados y excretores. Estos segmentos no son meramente un medio de<br />
transporte de la saliva, sino que permiten la formación de la saliva definitiva con reabsorciones y excreciones.<br />
A grandes rasgos los acinos serosos tienen conductos intercalares y estriados muy desarrollados que modifican<br />
la secreción serosa, mientras que en los acinos mucosos no se modifica la secreción, de modo que los<br />
intercalares están poco desarrollados y los estriados están ausentes.<br />
Conductos Intercalares o de Böll<br />
Los conductos intercalares están revestidos por células epiteliales cuboides bajas con actividad anhidrasa<br />
carbónica y que modifican la secreción salivar secretando ión bicarbonato y absorbiendo ión cloruro. Cuanta<br />
más acuosa sea la secreción de la glándula serosa más notable es este segmento.
Página41<br />
Conductos Estriados<br />
Los conductos estriados están formados por un epitelio cuboide simple que gradualmente se transforma en<br />
cilíndrico según se acerca al conducto excretor. La estriación se debe a una serie de repliegues laterales que<br />
contienen mitocondrias alargadas con orientación longitudinal. Estos repliegues con mitocondrias ap arecen en<br />
otras estructuras relacionadas con la reabsorción de líquido y electrolitos (<strong>com</strong>o en la nefrona). También<br />
aparecen abundantes repliegues baso-laterales con células vecinas; el núcleo es central.<br />
En este conducto se reabsorbe Na y se secreta K; <strong>com</strong>o se absorbe más Na del K que se excreta, la saliva final<br />
es hipotónica.<br />
Estos conductos suelen tener un gran diámetro, y se sitúan en el parénquima (son intralobulillares), pero<br />
pueden estar rodeados por pequeñas cantidades de tejido conectivo, que también suele contener vasos y<br />
nervios.<br />
Conductos Excretores<br />
Son los conductos más importantes de las glándulas salivales porque conducen la saliva final hasta la cavidad<br />
oral. El epitelio de estos conductos varía según aumenta el diámetro del conducto:<br />
Comienza siendo simple cuboide y gradualmente se transforma en estratificado o seudoestratificado cuboide;<br />
según aumenta el diámetro del conducto, aparece un epitelio estratificado cilíndrico que en la cavidad oral se<br />
convierte en estratificado plano.<br />
El conducto de la glándula parotídea (conducto de Stensen) y el de la glándula submaxilar (conducto de<br />
Wharton) transcurren por el tejido de la cara y el cuello, respectivamente, hasta que finalmente llegan a la<br />
cavidad oral.<br />
Glándulas Salivales Principales<br />
La glándula parótida es totalmente serosa, con conductos estriados grandes y conspicuos y una gran cantidad<br />
de tejido adiposo. Está atravesada por el VII par craneal; la parotiditis puede afectar a este nervio.<br />
La glándula submaxilar es mixta, predominantemente con glándulas serosas y alguna mucosa con semiluna<br />
serosa.<br />
La glándula sublingual es predominantemente mucosa, con glándulas mucosas con semilunas serosas y aisladas<br />
glándulas serosas.<br />
Saliva<br />
Las glándulas salivales producen cada día unos 1200mL de saliva, que permiten algunas funciones no<br />
metabólicas <strong>com</strong>o humedecer la mucosa oral, los alimentos secos para mejorar la deglución, disolver los<br />
alimentos para que puedan estimular a los botones gustativos, y otras funciones metabólicas <strong>com</strong>o la digestión<br />
de hidratos de carbono por la enzima digestiva α-amilasa y por una lisozima que controla la cantidad de flora<br />
bacteriana en la boca.<br />
La saliva también provee de minerales al esmalte para regenerarlo y también recubre los di entes con una<br />
cubierta proteica protectora.<br />
La saliva se encuentra formada por agua, P, glucoproteínas (enzimas y anticuerpos, <strong>com</strong>o la IgA) y electrolitos.<br />
La secreción de saliva responde a estímulos del SN Autónomo, pero también ante hormonas <strong>com</strong>o la<br />
aldosterona, que regula la reabsorción de Na y la expulsión de K a nivel de la nefrona, pero también actúa<br />
sobre los conductos estriados.
Página42<br />
FARINGE<br />
La faringe es un órgano hueco que histológicamente se puede estudiar según los dos epitelios que la<br />
<strong>com</strong>ponen:<br />
<br />
<br />
Epitelio plano estratificado no queratinizado que se continúa con la cavidad oral y el esófago, y que lo<br />
hace resistente a la abrasión.<br />
Epitelio cilíndrico ciliado que contiene células caliciformes y es continuo con el epitelio respiratorio.<br />
El tejido conectivo de la faringe es fibroelástico, con una gran cantidad de fibras elásticas, y que se asienta<br />
sobre los músculos constrictores de la faringe y otros músculos faríngeos.<br />
A cada lado y en la parte superior de la faringe aparecen las amígdalas faríngeas y palatinas, y se relaciona con<br />
el oído interno mediante la trompa de Eustaquio, que se abre a la nasofaringe cerca de los molares.<br />
GENERALIDADES DEL TUBO DIGESTIVO<br />
La pared del tubo digestivo presenta cuatro capas: mucosa, submucosa, muscular y adventicia.<br />
Mucosa<br />
La mucosa del tubo digestivo está formada por epitelio, lámina propia y la capa muscular de la mucosa ,<br />
llamada muscularis mucosae. La mucosa es la parte más variable a lo largo de las distintas zonas del tubo y se<br />
expone a luz del tubo es el medio externo.<br />
Funciones<br />
· Protección frente a entrada de patógenos y antígenos, por lo que encontramos tejido linfoide difuso y GALT.<br />
· Protección frente a abrasión física y química.<br />
· Protección frente a permeabilidad (zonas occludens).<br />
· Absorción selectiva de sales, agua y nutrientes. Depende de la superficie, por lo que, principalmente en el<br />
intestino delgado, hay una serie de mecanismos que aumentan la superficie de absorción:<br />
-Las válvulas conniventes o de Kerkring son pliegues circulares, con un eje de submucosa.<br />
-Las vellosidades o plicas están dispuestas en torno al eje de la válvula de Kerkring, con un eje de<br />
lámina propia.
Página43<br />
- Las microvellosidades están presentes en las células del epitelio<br />
· Digestión y procesamiento de los alimentos, ya que el glicocálix de las células epiteliales contiene enzimas.<br />
· Difusión de los nutrientes absorbidos a los vasos sanguíneos y linfáticos.<br />
· Secreción de moco protector, que lubrica y actúa a modo de tampón; enzimas y anticuerpos.<br />
A lo largo del tubo digestivo aparecen unas glándulas que derivan de invaginaciones del epitelio luminal y<br />
corresponden a:<br />
• Glándulas mucosas: en el seno de la lámina propia.<br />
• Glándulas submucosas: las secreciones pueden verterse en las glándulas mucosas o a la luz.<br />
• Glándulas extramurales: están fuera del tubo digestivo y entregan sus secreciones mediante conductos<br />
que atraviesan determinadas zonas de este. Así, englobamos a glándulas <strong>com</strong>o el páncreas o el hígado.<br />
Epitelio<br />
Es un epitelio de origen endodérmico. Forma el epitelio de revestimiento y es también el origen de todas las<br />
glándulas, que están tanto en la mucosa <strong>com</strong>o en la submucosa, e incluso forma todos los anejos que quedan<br />
fuera de la pared del tubo.<br />
Lámina Propia<br />
Es un conjuntivo laxo especialmente fibroso, con fibras colágenas, reticulares y elásticas. Está muy<br />
vascularizada e inervada y en ella se encuentra GALT.<br />
La presencia de más o menos lámina propia se asocia a la presencia de glándulas: cuántas más glándulas,<br />
menos lámina propia observamos, ya que su espacio es ocupado por las glándulas.<br />
Muscularis Mucosae<br />
Presenta un estrato interno, con los miocitos dispuestos circularmente; y otro externo, con los miocitos<br />
dispuestos longitudinalmente. Estos miocitos presentan un endomisio rico en fibras elásticas. Esta capa<br />
muscular es responsable de los pliegues de la mucosa, de modo que facilitan la absorción de nutrientes pero<br />
NO intervienen en el peristaltismo.<br />
Submucosa<br />
Es un conjuntivo laxo muy vascularizado, con abundantes GALT. Es muy flexible debido a la presencia de fibras<br />
elásticas.<br />
En la zona más externa aparece el plexo nervioso submucoso o de Meissner, que contiene fibras nerviosas<br />
simpáticas y parasimpáticas.<br />
Capa Muscular<br />
Está formada por miocitos lisos, excepto en la porción proximal del esófago.<br />
Presenta, <strong>com</strong>o la muscularis mucosae, un estrato interno, con los miocitos dispuestos circularmente (estrato<br />
circular interno o CI), y otro externo, con los miocitos dispuestos longitudinalmente (estrato longitudinal<br />
externo o LE). A veces puede encontrarse un tercer estrato. Cumplen 3 funciones:<br />
La capa circular interna es responsable de que en determinadas zonas se originen los distintos esfínteres 1 del<br />
tubo, <strong>com</strong>o el faringo-esofágico, que impide entrada de aire, el esófago-gástrico, que impide el reflujo gástrico,<br />
el pilórico, que controla la liberación del quimo, la válvula íleocecal, que retrasa el paso del contenido al ciego y<br />
controla el reflujo del colon, o el esfínter anal interno, que retiene las heces.<br />
Esta capa es responsable de que el contenido avance hacia el ano, gracias a los movimientos peristálticos 2 . Por<br />
estos movimientos, la contracción de los miocitos genera ondas de avance. Son movimientos espontáneos y<br />
rítmicos.
Página44<br />
Existe en esta capa un tono muscular basal y sostenido 3 , responsable del diámetro de la luz del tubo.<br />
Entre los dos estratos de esta capa muscular encontramos el plexo mientérico o de Auerbach, con fibras<br />
parasimpáticas (activadoras de la actividad muscular) y simpáticas (inhibidoras).<br />
Adventicia<br />
La adventicia es un conjuntivo laxo rico en adipocitos. Se asocia con el mesotelio peritoneal, formando serosas.<br />
Es una zona importante porque por ella entran y salen venas, arterias y nervios<br />
Vascularización<br />
Vascularización Sanguínea<br />
Los vasos sanguíneos llegan a la adventicia y atraviesan la muscular, la submucosa y la lámina propia, dando<br />
vasos que irrigan los miocitos, las glándulas, las criptas intestinales y las células del estrato basal del epitelio.<br />
Los vasos que encontramos en la mucosa del tubo son capilares fenestrados.<br />
El drenaje venoso se produce paralelamente el suministro arterial, drenando hacia la adventicia. Las venas del<br />
tubo digestivo presentan válvulas.<br />
Vascularización Linfática<br />
En el estómago, los capilares linfáticos forman un plexo entre las glándulas. Este plexo va por debajo del plexo<br />
capilar sanguíneo y drenan a vasos linfáticos de la submucosa. Estos vasos presentan válvulas y drenan hacia la<br />
muscular.<br />
En el intestino delgado, parte de los nutrientes se absorben vía linfáticos, <strong>com</strong>o las grasas.<br />
Inervación<br />
La inervación aferente está regentada por fibras aferentes viscerales generales, que vehiculan hambre, dolor,<br />
etc.<br />
La inervación eferente está formada por fibras pertenecientes al SNA, es decir, simpáticas y parasimpáticas.<br />
La inervación intrínseca, que conforma el sistema nervioso entérico, es una serie de ganglios y nervios que se<br />
extienden desde el esófago hasta el esfínter anal. Son independientes del control simpático y parasimpático,<br />
aunque están en conexión con éste: recibe información de axones del sistema simpático y parasimpático, de<br />
modo que es capaz de responder a estímulos locales y además integra señales que llegan desde el SNC.<br />
Este sistema nervioso entérico se dispone en dos plexos: el plexo submucoso o de Meissner, situado entre los<br />
estratos de la muscularis mucosa; y el plexo mientérico o de Auerbach, situado entre los estratos de la capa<br />
muscular del tubo. En estos plexos se observan ganglios con neuronas asociadas a células gliales, todo rodeado<br />
de elementos conjuntivos. En los haces nerviosos de los plexos encontramos las células intersticiales de Cajal.<br />
Funciones<br />
· Control del peristaltismo.<br />
· Control de los movimientos de la muscularis mucosae.<br />
· Control de la secreción de las glándulas.
Página45<br />
ESÓFAGO<br />
Generalidades<br />
El esófago es un tubo muscular de 25 cm situado entre la faringe y el estómago, que <strong>com</strong>ienza a nivel del cartílago<br />
cricoides y acaba 2-4 cm por debajo de diafragma<br />
El traspaso de material a través del esófago no puede interferir con la respiración ni con la circulación pulmonar<br />
Mucosa<br />
La mucosa esofágica presenta pliegues longitudinales. Es de tonalidad rosácea excepto en la zona distal, donde<br />
adquiere un color más mas blanquecino, y acaba en la línea Z.<br />
Epitelio<br />
La mucosa esofágica está tapizada por un epitelio plano no queratinizado<br />
Las células epiteliales cambian según la dieta, que tienden a aplanarse y acumular glucógeno. Entre ellas<br />
encontramos células de Langerhans.<br />
La función de este epitelio es eminentemente protectora. En la parte más distal es menos resistente, porque puede<br />
tener características no esofágicas, <strong>com</strong>o parches de epitelio gástrico o intestinal.<br />
El esófago de Barrett puede deberse a que en la persona que lo posee hay un fallo en el esfínter esófagogástrico que<br />
produce reflujo gastroesofágico. El esófago de Barrett condiciona una tendencia al reflujo y una reacción de la<br />
mucosa esofágica, que tiende a inflamarse. A largo plazo, estos parches son precursores de metaplasias.<br />
Lámina Propia<br />
Es un conjuntivo laxo con fibras elásticas y células linfoides, tanto dispersas <strong>com</strong>o en forma de nódulos (GALT), y con<br />
glándulas esofágicas cardiales.<br />
Muscularis Mucosae<br />
La forman miocitos lisos dispuestos longitudinalmente, y a su alrededor una red de fibras elásticas, aunque según<br />
avanzamos, aparecen miocitos que se disponen circularmente. La muscularis mucosae facilita el aplanamiento de los<br />
pliegues y con ello la deglución.<br />
Submucosa<br />
Es un conjuntivo denso con fibras elásticas y GALT. Es característica la presencia de glándulas esofágicas propiamente<br />
dichas en esta capa<br />
Aparece el plexo de Meissner y vascularización, tanto sanguínea <strong>com</strong>o linfática.<br />
Como consecuencia de otras patologías, puede producirse la rotura de venas esofágicas, por ejemplo en cirrosis<br />
hepática que, produce hipertensión. A veces se da la rotura de varices por otras razones producen hemorragias que<br />
drenan al estómago.<br />
Capa Muscular<br />
La capa muscular presenta dos estratos: un CI y un LE. En el tercio más proximal, las dos capas son miocitos<br />
estriados, no lisos, en el tercio medio, los miocitos estriados se <strong>com</strong>binan con miocitos lisos y en la parte más distal<br />
solo hay ya miocitos lisos.<br />
Los miocitos esqueléticos están inervados por neuronas motoras que están situadas en el núcleo ambiguo; mientras<br />
que los miocitos lisos reciben los impulsos de neuronas viscerales del núcleo motor dorsal. En esta capa muscular<br />
encontramos el plexo de Auerbach.
Página46<br />
Esfínteres<br />
En el esófago encontramos dos esfínteres: un esfínter superior o cricofaríngeo, que controla el inicio de la deglución,<br />
y un esfínter inferior o gastroesofágico. El primero es un esfínter físico, y el segundo es funcional. Ambos esfínteres<br />
están cerrados en condiciones normales, de manera tónica<br />
Los esfínteres presentan inervación a cargo de fibras parasimpáticas estimuladoras, mediadas por ACh y sustancia P;<br />
así <strong>com</strong>o por fibras del sistema nervioso entérico, mediadas por VIP y NO.<br />
Si los esfínteres no funcionan correctamente, el estómago tiende a deslizarse hacia arriba, provocando una hernia de<br />
hiato. La acalasia se produce si el esfínter no funciona correctamente (no se abre) porque durante el desarrollo<br />
embrionario se han perdido neuronas inhibidoras del plexo de Auerbach.<br />
Glándulas<br />
Glándulas Esofágicas Propiamente Dichas: se encuentran en la submucosa de todo el esófago; son túbuloacinares<br />
<strong>com</strong>puestas, de secreción mucosa, con función lubricante y túbulo excretor con epitelio estratificado plano<br />
generalmente dilatado. Es relativamente frecuente que esta secreción viscosa quede retenida en los conductos<br />
excretores y den lugar a quistes.<br />
Glándulas Esofágicas Cardiales: aparecen, generalmente, en la lámina propia del tercio distal del esófago. Producen<br />
un moco neutro que protegen al esófago de las secreciones estomacales (ácidas).<br />
Adventicia<br />
La adventicia esofágica es un conjuntivo laxo. En su porción más distal se asocia con el peritoneo, formando una<br />
serosa.<br />
Microfotografía de una glándula esofágica propiamente dicha (submucosa)
Página47<br />
ESTÓMAGO<br />
El estómago es una región dilatada del tubo digestivo, con una capacidad de 1,5L.<br />
Desde el punto de vista anatómico, el estómago está dividido en<br />
cuatro regiones: el cardias 1 , que rodea el orificio de desmbocadura de<br />
esófago, el fundus o techo 2 , el cuerpo 3 y la región pilórica o<br />
antropliórica 4 .<br />
Pero histológicamente, se distinguen 3 grandes regiones según el tipo<br />
de glándulas que presenten:<br />
1. Región cardial o cardias: contiene glándulas cardiales;<br />
corresponde al cardias.<br />
2. Región pilórica o antro: contiene glándulas pilóricas o<br />
cardiales y corresponde a la región pilórica.<br />
3. Región fúndica o fundus: es la parte más extensa y contiene glándulas fúndicas o gástricas; engloba al fundus<br />
y al cuerpo estomacales.<br />
Funciones<br />
· Digestión (homogeneización de alimentos)<br />
· Secreción de enzimas y HCl<br />
· Secreción de factor intrínseco<br />
· Almacenamiento del quimo<br />
Estructura Histológica General<br />
Al igual que todo el tubo digestivo, el estómago presenta una mucosa con epitelio, lámina propia y muscularis<br />
mucosae, una submucosa, una capa muscular y una adventicia.<br />
El estómago presenta arrugas gástricas con orientación longitudinal que se denominan rugae, cada una de las cuales<br />
presenta un eje submucoso. Son más profundas en la región pilórica y fúndica Estas arrugas permiten adaptarse al<br />
contenido estomacal, porque pueden distenderse al llenarse.<br />
Estas arrugas presentan a su vez elevaciones o surcos, que aumentan ligeramente la superficie de la mucosa para la<br />
secreción; estos se conocen <strong>com</strong>o regiones mamiladas o mamelones.<br />
Con un estudio a mayor aumento se ven unos orificios que corresponden a las fovéolas, criptas gástricas, fositas o<br />
fosetas. Se trata de invaginaciones profundas del epitelio gástrico que se continúan con glándulas. Son más<br />
profundas en el antro.<br />
Mucosa<br />
Es un epitelio simple cilíndrico (en ocasiones cúbico) formado por unas células mucosas llamadas células mucosas<br />
superficiales.<br />
CÉLULAS MUCOSAS SUPERFICIALES<br />
Son células dispuestas en el epitelio estomacal que segregan mucógenos, por lo que son células PAS +.<br />
Liberan a la luz iones bicarbonato procedentes de las células oxínticas, de modo que protegen al estómago con su<br />
basicidad de la acidez de su propio jugo. El moco es muy viscoso y turbio, y se sitúa sobre la mucosa, formando una<br />
capa de más de 100 μm de grosor evitando que los alimentos que se mueven en el estómago rocen y dañen la<br />
mucosa.<br />
Este epitelio presenta zónulas occludens y adherens, haciendo que sea un epitelio hermético.<br />
Son células capaces de absorber agua, sales y productos lipososolubles. Lo que explica el rápido efecto de algunos<br />
fármacos (aspirina) y del alcohol.
Página48<br />
Lámina Propia<br />
Es un tejido conjuntivo que parece muy escaso por la abundancia de glándulas que lo ocupan. Presenta muchas<br />
fibras colágenas y reticulares, pero no elásticas. Se encuentran asimismo haces de miocitos lisos en región pilórica<br />
Glándulas Fúndicas<br />
Las glándulas más abundantes son las glándulas fúndicas, por lo que en ocasiones también se las denomina glándulas<br />
gástricas. Es por ello por lo que sirven de referencia para el estudio de los tipos celulares, <strong>com</strong>parando algunas<br />
características con el resto de glándulas del estómago.<br />
Cada glándula fúndica se <strong>com</strong>unica con la fovéola mediante una estructura llamada istmo, un corto segmento celular<br />
poblado por células madre, en número reducido, que aumenta cuando se producen daños en la mucosa. Se encargan<br />
de renovar tanto el epitelio foveolar y de revestimiento, así <strong>com</strong>o las glándulas, es decir, de un solo tipo de célula<br />
madre se pueblan todas las glándulas y el epitelio.<br />
La glándula gástrica es una glándula tubular simple ramificada que se puede dividir en un cuello angosto y bastante<br />
largo que se continúa con una base o fondo de mayor amplitud, que suele dividirse en dos o tres ramas que se<br />
enrollan levemente cerca de la muscularis mucosae.<br />
a. Glándula Fúndica, corte histológico<br />
b. Glándula Fúndica, esquema con principales células<br />
Las glándulas fúndicas están formadas por varios tipos celulares:
Página49<br />
CÉLULAS MUCOSAS O CÉLULAS MUCOSAS DEL CUELLO<br />
Se sitúan en el cuello de las glándulas. Son similares a las células mucosas foveolares y del epitelio de revestimiento,<br />
pero carecen de la dilatación apical prominente; además, son más pequeñas, de citoplasma pálido, con menos<br />
gránulos de mucígeno (PAS+), y con un núcleo redondo y basal.<br />
Segregan un moco más líquido y soluble que el de las células mucosas superficiales, en respuesta a estímulos<br />
vagales.<br />
CÉLULAS OXÍNTICAS, DELOMORFAS O PARIETALES<br />
Se encuentran en cualquier zona de la glándula. Son células altas, prismáticas, con una zona basal ensanchada,<br />
dándoles una forma abombada que las hace distinguibles. Presentan un núcleo redondo central y un citoplasma<br />
eosinófilo.<br />
Por impregnación argéntica se obtiene una preparación en la cual en estas células queda resaltada parte del polo<br />
apical y que corresponde a los canalículos intracelulares. Cuando se les aplica esta tinción se les da el nombre de<br />
células en garra.<br />
No tienen muchos orgánulos, ni gránulos, ni vesículas, pero sí tienen muchas mitocondrias con grandes crestas<br />
mitocondriales, ya que en estas células hay un alto consumo de oxígeno por encima de lo habitual.<br />
Las células oxínticas segregan HCl y factor intrínseco (FI).<br />
La membrana plasmática en su porción apical presenta algunas<br />
microvellosidades y está invaginada. Estas invaginaciones forman los<br />
canalículos intracelulares, muy numerosos y de trayectoria tortuosa que<br />
llegan hasta el citoplasma basal, es decir, son muy profundas. Presenta<br />
un sistema de membranas túbulovesicular.<br />
En su membrana plasmática, encontramos una inmensa mayoría de<br />
bombas ATPasas, concretamente una variante de la bomba Na+/K+ que<br />
saca protones y mete potasio.<br />
La célula oxíntica funciona de manera que por su membrana basal toma<br />
CO 2 del plasma de los capilares de la lámina propia. La célula tiene<br />
enzima anhidrasa carbónica que hace que el CO 2 se una con el agua<br />
formando ácido carbónico, que se escinde formando protones y<br />
bicarbonato.<br />
La membrana basal presenta además canales para cloro que permiten<br />
incorporar iones Cl- a la célula. La célula expulsa estos iones por su membrana apical, de manera que encontramos<br />
un doble mecanismo que permite que la célula tome protones y cloro del exterior para expulsarlos a la luz formando<br />
HCl.<br />
La bomba de protones es la base del funcionamiento del omeprazol, un fármaco diseñado originalmente para<br />
bloquear esta ATPasa en el tratamiento de úlceras.<br />
Los iones bicarbonato formados salen por la membrana basal y alcanzan la sangre, que está circulando hacia la<br />
superficie, arrastrando hacia el epitelio de revestimiento altas concentraciones de bicarbonato. Entonces los iones<br />
difunden atravesando el epitelio revestimiento o foveolar hacia la barrera de moco, incrementando el pH del mismo.<br />
Las células oxínticas muestran cambios en función del estado funcional. Afectan a la cantidad de membrana y al<br />
sistema túbulovesicular. Este sistema es un reservorio de membrana apical: cuando las células segregan HCl,<br />
necesitan mas membrana, fusionándose el reservorio con la membrana apical, aumentando así la superficie de<br />
secreción y el número de microvellosidades. También cambia la densidad de mitocondrias
Página50<br />
La secreción ácida de las células oxínticas presenta un control nervioso y hormonal:<br />
La gastrina, hormona sintetizada por la pared gástrica e intestinal, estimula su secreción.<br />
La acetilcolina o ACh es un neurotransmisor liberado por las fibras del SNA, estimulando las zonas secretoras<br />
de gastrina. Actúa sobre los receptores acetilcolínicos M 3 .<br />
La histamina estimula al SNA y de forma directa incide sobre las células productoras de gastrina.<br />
El síndrome de Zollinger-Ellison supone una secreción permanente de HCl, provocando ulceras gástricas y duodenales.<br />
Esta secreción desregulada se debe a un tumor de las células secretoras de gastrina (gastrinoma).<br />
Además de HCl, las células oxínticas segregan también factor intrínseco (FI), una glicoproteína esencial, porque, si<br />
no se asocia a vitamina B12, ésta no es absorbida en el íleon.<br />
La regulación de la síntesis de FI es igual que la de gastrina.<br />
La aclorhidria es una enfermedad autoinmune en la que se destruyen las células oxínticas, por lo que no se sintetiza FI<br />
y se sufre anemia perniciosa por carencia de vitamina B12. Esta también aparece por la enfermedad ulcerosa péptica,<br />
en la que las úlceras cicatrizadas presentan parches que no sintetizan FI de manera eficaz.<br />
CÉLULAS PRINCIPALES, ZIMOGÉNICAS O ADELOMORFAS<br />
Se localizan en el fondo de las glándulas fúndicas.<br />
Son células altas, prismáticas, de secreción serosa, con un citoplasma muy basófilo que se debe la presencia de un<br />
RER muy desarrollado que se localiza en la zona más basal del citoplasma, junto al núcleo. Es frecuente que a veces<br />
no se observen porque son difíciles de preservar en la preparación.<br />
En el citoplasma apical destaca una enorme cantidad de gránulos que contienen lipasa y pepsinógeno<br />
fundamentalmente. Cuando el pepsinógeno se libera fuera de la célula, se encuentra un ambiente ácido que<br />
promueve su transformación en pepsina. También secreta una lipasa débil.<br />
Las células principales responden a una regulación hormonal a secretina.<br />
CÉLULAS ENDOCRINAS, ENTEROENDOCRINAS O NEUROENDOCRINAS<br />
Se encuentran en cualquier zona de la glándula, aunque tienden a localizarse en la base, y se pueden diferenciar a<br />
MO con tinciones de metales pesados, <strong>com</strong>o la plata, por lo que también se conocen <strong>com</strong>o células enterocromafines,<br />
células argentafines o argentófilas. En la actualidad, las diferentes IHQ permiten diferenciar diferentes tipos de<br />
células enteroendocrinas.<br />
Son células apoyadas en la lámina basal que no siempre alcanzan la luz, aunque en ocasiones presentan<br />
prolongaciones citoplasmáticas muy delgadas con microvellosidades que alcanzan la luz. Se caracterizan porque<br />
presentan una gran cantidad de gránulos en el polo basal. Así, se cree que son capaces de controlar la <strong>com</strong>posición<br />
del jugo gástrico y poder responder ante los cambios que se produzcan en él.<br />
Contienen una amplia variedad de hormonas implicadas en la regulación de las funciones gástricas:<br />
· Serotonina: estimula el peristaltismo.<br />
· Histamina: estimula la secreción de HCl.<br />
· Somatostatina: inhibe la secreción y la motilidad.<br />
· Gastrina: estimula la secreción HCl y el peristaltismo; y activa la liberación de CCK.<br />
· Grelina: estimula secreción de hormona del crecimiento (GH) y estimula los centros del apetito; su secreción está<br />
aumentada durante el sueño, por lo que disminuye el apetito y aumenta la concentración de GH mientras se<br />
duerme.<br />
Las células enteroendocrinas aparecen a lo largo del tubo digestivo, islotes de Langerhans y en el sistema respiratorio<br />
conformando el mayor sistema endocrino del organismo: el sistema neuroendocrino difuso. Este sistema está<br />
formado por las células endocrinas gastroenteropancreáticas o GEP y secretan, de manera similar a las células<br />
neurosecretoras del SNC, hormonas y agentes reguladores. Así, estas células secretan:<br />
<br />
<br />
Hormonas: gastrina, secretina, colecistoquinina (CCK), péptido inhibidor gástrico (GIP), motilina,<br />
somatostonina,…<br />
Neurotransmisores: ACh, Péptido Intestinal Vasoactivo (VIP), bombesina, encefalinas,…
Página51<br />
Jugo Gástrico<br />
El jugo gástrico está formado por dos secreciones:<br />
· Constitutiva: es la capa de moco protectora de la mucosa<br />
· Facultativa: <strong>com</strong>puesta por pepsina y HCl agresivas para con los alimentos y la mucosa<br />
El jugo gástrico contiene agua y sales minerales, por lo que pacientes con vómitos crónicos requieren hidratación y<br />
suministro de K+ suficiente para evitar una acidosis metabólica hipopotasémica. El jugo gástrico contiene además FI.<br />
La capa protectora que recubre la mucosa permite tamponar el HCl y además permite separar la masa que se está<br />
digiriendo de la mucosa. Es frecuente que en la capa de moco habite Helicobacter pylori (95% de pacientes ulcerosos<br />
lo tienen, así <strong>com</strong>o el 20% de gente de 20 años y el 90% de ancianos). Esta bacteria secreta amoniaco y proteasas<br />
citotóxicas. No es detectada por el organismo porque presenta en su superficie oligosacáridos que parecen propios.<br />
El HCl permite digerir alimentos por activación del pepsinógeno y tiene además propiedades bacteriostáticas.<br />
Glándulas Cardiales y Pilóricas<br />
Las Glándulas Cardiales permiten con su secreción conjunta a la de las glándulas esofágicas cardiales proteger el<br />
epitelio esofágico. En general son glándulas tubulares poco tortuosas y a veces ramificadas <strong>com</strong>puestas<br />
principalmente por células secretoras de moco y ocasionalmente células enteroendocrinas.<br />
Las Glándulas Pilóricas son tubulares enrolladas y ramificadas con una luz amplia y gran renovación celular.<br />
Muscularis Mucosae<br />
Presenta un estrato interno, con los miocitos dispuestos circularmente; y otro externo, con los miocitos dispuestos<br />
longitudinalmente. A veces puede encontrarse un tercer estrato.<br />
Encontramos en ella glándulas tubulares ramificadas largas y de luz estreha, de modo que necesitan <strong>com</strong>primiese<br />
para expulsar la secreción.<br />
Submucosa<br />
Es un conjuntivo laxo con gran cantidad de adipocitos y muchos vasos. Es frecuente la presencia de GALT. No hay<br />
glándulas. Aparece en ella el plexo de Meissner.<br />
Capa Muscular<br />
Compuesta por miocitos lisos presenta, además de los estratos habituales (interno circular y externo longitudinal),<br />
un tercer estrato oblicuo más interno.<br />
En este estrato oblicuo los miocitos se disponen de manera helicoidal que ayuda a que el estómago se adapte a<br />
cambios de volumen sin que cambie su presión interna, cambiando el tono muscular.<br />
La capa circular forma el esfínter pilórico, que cuando se relaja permite que el quimo pase hacia el duodeno.<br />
En esta capa muscular encontramos el plexo de Auerbach.<br />
Adventicia<br />
Se haya asociada al peritoneo, formando una serosa.
Página52<br />
INTESTINO DELGADO<br />
El intestino delgado es la porción del tubo digestivo situada entre el estómago y el intestino grueso. Mide 5m,<br />
longitud que aumenta en post mortem. Es una zona donde se segregan enzimas y hormonas, termina la<br />
digestión y se produce la absorción de los nutrientes.<br />
Anatómicamente, el intestino delgado se divide en duodeno, yeyuno e íleon.<br />
El duodeno es la parte que discurre desde el píloro al ángulo duodenoyeyunal en forma de C alrededor de la<br />
cabeza páncreas, y tiene unos 25 cm de longitud.<br />
El yeyuno es la siguiente porción, con una longitud de 2,5 m.<br />
El íleon termina a la altura de válvula íleocecal y tiene una longitud de 3,5 m.<br />
Al igual que todo el tubo digestivo, el intestino delgado presenta una mucosa con epitelio, lámina propia y<br />
muscularis mucosa; una submucosa, una capa muscular y una adventicia.<br />
El intestino delgado tiene una serie de estructuras que permiten aumentar su superficie de absorción en el<br />
mismo espacio. Son:<br />
Pliegues circulares, válvulas conniventes o de Kerkring: son repliegues transversales permanentes que<br />
presentan un eje de submucosa, y son muy abundantes en el yeyuno y el duodeno distal. Tienen forma de<br />
C que se extiende por la mitad o dos terceras partes de la circunferencia de la luz intestinal.<br />
Vellosidades o Plicas: son proyecciones digitiformes (foliáceas en niños)<br />
que se disponen sobre las válvulas conniventes y tienen un eje de<br />
lámina propia. Son muy abundantes en el yeyuno proximal y el<br />
duodeno, pero alcanzan su altura máxima en el yeyuno.<br />
Tienen un centro de tejido conjuntivo laxo cubierto por un epitelio<br />
cúbico simple cilíndrico. El centro de la vellosidad es una extensión de<br />
la lámina propia que contiene abundantes fibroblastos, células<br />
musculares lisas, linfocitos, plasmocitos, eosinófilos, macrófagos y una<br />
red de capilares fenestrados subyacentes a la lámina propia, así <strong>com</strong>o<br />
de un capilar linfática que se origina en el fondo de un saco ciego<br />
llamado vaso quilífero central a donde drena la linfa, impulsada a vasos<br />
mayores por las contracciones de la capa muscular lisa.<br />
Criptas de Lieberkühn: son invaginaciones que corresponden a<br />
glándulas tubulares simples cuya base queda en la zona profunda de la<br />
lámina propia.<br />
Microvellosidades: son diferenciaciones de la membrana apical de los<br />
enterocitos. Son regulares en grosor y longitud, por lo que se ven <strong>com</strong>o chapa estriada a microscopía<br />
óptica.<br />
Estos mecanismos permiten un incremento de la superficie de absorción hasta los 200 m 2 .<br />
Mucosa<br />
En el epitelio de la mucosa intestinal encontramos células epiteliales prismáticas simples con la chapa estriada<br />
que forman las microvellosidades (PAS+) En este epitelio encontramos enterocitos, células caliciformes y<br />
endocrinas, así <strong>com</strong>o linfocitos y células M.
Página53<br />
ENTEROCITOS<br />
Los enterocitos son las células<br />
características del epitelio intestinal.<br />
Presentan una membrana apical con<br />
microvellosidades, en conexión con el<br />
velo terminal, que cuando se contrae<br />
tira de los filamentos de actina que<br />
forman las microvellosidades, haciendo<br />
que éstas encojan. Esto facilita la<br />
exposición de la superficie de las<br />
microvellosidades durante la absorción.<br />
La membrana de las microvellosidades<br />
tiene un glicocálix asociado, con<br />
enzimas que permiten terminar la<br />
digestión al escindir azúcares, pequeños<br />
lípidos, y terminan de digerir péptidos<br />
pequeños, produciendo aminoácidos.<br />
Estas microvellosidades permiten<br />
aumentar la superficie de absorción<br />
apical hasta en 600 veces.<br />
Los enterocitos presentan una gran<br />
cantidad de proteínas a modo de<br />
transportadores.<br />
Los enterocitos presentan zónulas occludens muy herméticas, sobre todo en el íleon, mediante las que se unen<br />
entre ellas y con las células caliciformes y las enteroendocrinas.<br />
Su membrana basolateral presenta una ATPasa que permite sacar iones Na + , lo que atrae agua, creando un<br />
incremento de la presión hidrostática. En el yeyuno y el duodeno, los enterocitos son más permeables y<br />
permiten que se de el <strong>com</strong>o arrastre del solvente: una absorción directa agua y Na+ al espacio extracelular. En<br />
estas zonas el enterocito absorbe mucha agua y Na+ y lo pasa a la zona basolateral, creando una tendencia a<br />
arrastrar agua a la lámina propia.<br />
Su núcleo es alargado y se sitúa en la zona media. Sobre este núcleo se disponen mitocondrias. El enterocito es<br />
una célula con un importante aparato de Golgi.<br />
En los enterocitos encontramos ácidos grasos absorbidos, parte de los cuales son directamente llevados a<br />
membrana basolateral para salir de la célula por la lámina propia hacia la sangre. Los ácidos de cadena más<br />
larga son utilizados por el enterocito para sintetizar triglicéridos. Esta síntesis ocurre en el REL y, una vez que se<br />
forman los triglicéridos, se transforman en quilomicrones. Los quilomicrones se vierten fuera de la célula por la<br />
membrana basolateral y el flujo del agua arrastra los quilomicrones hacia los vasos quilíferos.<br />
Cuando el enterocito absorbe muchos productos, la membrana presenta muchos pliegues, creando espacios<br />
extracelulares más grandes<br />
Los enterocitos segregan además <strong>com</strong>ponentes del glicocálix e IgA-secretor.<br />
Los síndromes de mala absorción se dan por causas variadas: alteración de las enzimas, de modo que no se<br />
digieren bien las alimentos; alteraciones de los transportadores, digestión anómala de proteínas y grasas<br />
(pancreatitis, fibrosis quística) o falta de solubilización de grasas por defecto en la secreción biliar. Estos<br />
síndromes afectan a múltiples sistemas y cursan con diarrea.<br />
CÉLULAS CALICIFORMES<br />
Son células pálidas, con forma de cáliz. Interrumpen la chapa estriada y la reducen a una zona llamada teca<br />
localizada en la porción apicolateral. Presentan un núcleo alargado, heterocromático y basal, donde el<br />
citoplasma es basófilo y hay abundante RER y ribosomas libres.<br />
Aumentan según se avanza hacia posiciones distales del intestino delgado. Presentan gránulos de mucígeno,<br />
que tiene carácter protector frente al contenido ácido.
Página54<br />
CÉLULAS DE PANETH<br />
Se sitúan en el fondo de las criptas y son más numerosas según avanzamos hacia distal. Son células altas de<br />
núcleo redondeado, con una importante dotación de orgánulos, <strong>com</strong>o un gran Aparato de Golgi supranuclear,<br />
lo que hace que citoplasma basal sea basófilo. En la zona apical encontramos gránulos refringentes eosinófilos,<br />
que tienen una mezcla de sustancias: proteínas ricas en arginina, enzimas <strong>com</strong>o la lisozima, TNFα…<br />
Encontramos también α-defensinas o criptidinas, capaces de perforar las membranas de bacterias y parásitos,<br />
creando canales que aumentan la permeabilidad de éstos. Además, son capaces de estimular el acercamiento<br />
de células dendríticas y actúan <strong>com</strong>o mediadores de los linfocitos T CD8+, facilitando la reacción inmunitaria.<br />
Las células de Paneth regulan el nivel de la flora bacteriana intestinal y eliminan protozoos. Presentan picos de<br />
secreción después de una ingesta.<br />
CÉLULAS M<br />
Las células M provienen de células madre epiteliales que expresan CMH-II. Se sitúan sobre las Placas de Peyer y<br />
otros nódulos linfáticos grandes. No tienen microvellosidades, aunque presentan micropliegues en el dominio<br />
apical e invaginaciones o bolsillos en el dominio basolateral, donde se sitúan linfocitos B, T y prolongaciones de<br />
macrófagos y dendríticas. Las células M captan los microorganismos y antígenos por endocitosis y éstos viajan<br />
en vesículas con catepsina B y son expresados en el dominio basolateral para que los linfocitos B puedan<br />
reconocer el antígeno. Algunos virus y bacterias se sirven de las células M para poder entrar en el organismo a<br />
través de las barreras externas. Las células M constituyen además un punto de investigación para la fabricación<br />
de vacunas orales.<br />
Las células inmunitarias salen de los bolsillos y migran hacia a la región donde están los folículos. Los linfocitos<br />
B se transforman allí en células plasmáticas e invaden la mucosa e incluso pueden migran a otros órganos.<br />
Estas células plasmáticas secretan mayoritariamente IgA, aunque también pueden secretar IgG, IgM e IgE.<br />
CÉLULAS ENTEROENDOCRINAS<br />
Se encuentran en la base de las glándulas, y pueden diseminarse por todo el epitelio glandular y de<br />
revestimiento. Son argirófilas y cromafines. Presentan gránulos de imagen variable según el contenido. Forman<br />
hormonas y péptidos neuroendocrinos.<br />
Secretan una gran variedad de hormonas:<br />
• Colecistoquinina (CCK): se encuentra en duodeno y en yeyuno. Tiene efectos diversos: inhibe el vaciado<br />
gástrico, estimula el crecimiento del páncreas, estimula la pared muscular de la vesícula y la secreción de<br />
jugo pancreático.<br />
• Secretina: se libera en duodeno. Estimula el crecimiento del páncreas, la contracción de la vesícula biliar y<br />
la secreción de enzimas pancreáticas y de HCO 3 - . Inhibe la secreción ácida.<br />
• Polipéptido inhibidor gástrico (GIP): se libera en duodeno y yeyuno. Estimula la liberación de insulina e<br />
inhibe la secreción ácida. Actúa sobre los miocitos del intestino y del estómago, estimuland o el<br />
peristaltismo.<br />
• Motilina: se libera en duodeno y yeyuno. Estimula la motilidad gástrica e intestinal.<br />
• Péptido vasoactivo intestinal (VIP): estimula las secreciones intestinales y pancreáticas, inhibe los<br />
esfínteres y la musculatura en general.<br />
• Encefalinas: estimulan la contracción muscular e inhiben las secreciones intestinales.<br />
Asimismo, las células endocrinas segregan una serie de hormonas paracrinas<br />
• Somatostatina: hormona inhibidora de la liberación de gastrina y otras hormonas gastrointestinales, así<br />
<strong>com</strong>o de la secreción ácida gástrica.<br />
• Histamina: hormona estimulante de la ecreción ácida gástrica.<br />
CÉLULAS MADRE<br />
Permiten renovar el epitelio de revestimiento y glandular y están en la base de las criptas. Se dividen<br />
activamente y migran. Tienen una tasa de renovación alta, que es máxima en el yeyuno.
Página55<br />
Lámina Propia<br />
Es un conjuntivo laxo rico en fibras colágenas y con abundantes células (fibroblastos, miocitos, linfocitos,<br />
plasmáticas, eosinófilos, macrófagos). Forma el eje de cada vellosidad intestinal y en dicho eje encontramos los<br />
vasos quilíferos, que captan el quilo y la linfa, impulsados por los miocitos. Bajo la lámina basal encontramos<br />
una red de capilares sanguíneos fenestrados.<br />
La lámina propia presenta abundantes células linfoides periglandulares y GALUs, así <strong>com</strong>o las acumulaciones<br />
linfoides conocidas <strong>com</strong>o placas de Peyer.<br />
La placa de Peyer se forma cuando en una zona concreta del tubo digestivo se desarrolla un número muy<br />
elevado de folículos de tejido parafolicular. Aunque pueden aparecer en cualquier parte, se suelen formar en la<br />
pared opuesta a donde se inserta el mesenterio. El número máximo de placas de Peyer es de más de 300 y se<br />
alcanza en la infancia.<br />
La estructura histológica general del tubo digestivo está alterada en las placas de Peyer: apenas se encuentran<br />
criptas de Lieberkuhn y se disgrega la muscularis mucosae, que es parte de la mucosa del digestivo, porque los<br />
folículos son tan grandes que no caben en la mucosa e invaden la submucosa. Por su parte, el epitelio intestinal<br />
que cubre las placas apenas contiene células caliciformes. Los enterocitos dejan de ser células absortivas y<br />
adquieren una alta capacidad para degradar antígenos.<br />
En el epitelio asociado a los folículos (FAE) se localizan células M y linfocitos T.<br />
La región cupular de la placa de Peyer se <strong>com</strong>pone de la cúpula y el epitelio cupular. La cúpula es una banda de<br />
lámina propia con abundancia de células inmunitarias, sobre todo linfocitos B, aunque también hay<br />
macrófagos, células dendríticas… La cúpula rodea o abraza el centro germinal.<br />
El centro germinal se <strong>com</strong>pone de linfocitos B, T CD4+ y células presentadoras de antígenos. Los linfocitos B<br />
acuden atraídos por las células dendríticas y se transforman en células plasmáticas. Éstas, en su inmensa<br />
mayoría, migran hacia otros folículos próximos de la placa o a zonas mas alejadas de la mucosa. La mayoría de<br />
las células plasmáticas secretan IgA, aunque también se encuentra IgM e IgE, que se libera para atraer<br />
mastocitos que liberan histamina. El enterocito capta la IgA después de que se dimeriza por acción de la<br />
cadena J. Este <strong>com</strong>plejo se une a un receptor en la membrana basal del enterocito. Se libera entonces la IgA<br />
unida al <strong>com</strong>ponente secretor (IgA secretor). IgA se une al <strong>com</strong>ponente secretor para dificultar su degradación<br />
y la unión de los microorganismos a los enterocitos.<br />
Durante una enteritis las placas de Peyer crecen tanto en longitud <strong>com</strong>o en grosor.<br />
La lámina propia se encuentra ocupada por profundas criptas de Lieberkühn, en la que encontramos<br />
enterocitos, células caliciformes, endocrinas, madre y de Paneth.<br />
Muscularis Mucosae<br />
Presenta un estrato interno, con los miocitos dispuestos circularmente; y otro externo, con los miocitos<br />
dispuestos longitudinalmente. Permite mezclar mejor el contenido intestinal.<br />
Submucosa<br />
Es un conjuntivo laxo con adipocitos frecuentes y acúmulos linfoides. En el duodeno aparece la glándula de<br />
Brunner.<br />
Glándula de Brunner<br />
Es una glándula tubular ramificada de naturaleza mucosa, cuya secreción es drenada al fondo de las criptas de<br />
Lieberkühn. Es una secreción de glicoproteínas que van a permitir dar un pH neutro o básico al moco duodenal.<br />
Esto es esencial para procesar el quimo y activar las enzimas pancreáticas.<br />
También se detecta en estas glándulas la urogastrona, un factor de crecimiento epidérmico (EGF). La<br />
urogastrona es resistente a enzimas digestivas y actúa en diferentes órganos, puesto que hay receptores suyos<br />
en hepatocitos y glándulas gástricas. La urogastrona inhibe la secreción ácida gástrica. Estimula asimismo a las<br />
células madre, es decir, estimula la renovación y crecimiento de la mucosa digestiva.<br />
Parece ser que interviene en situaciones de reparación o regeneración tras daño tisular.
Página56<br />
Capa Muscular<br />
Presenta un estrato interno, con los miocitos dispuestos circularmente; y otro externo, con los miocitos<br />
dispuestos longitudinalmente. Permite mezclar mejor el contenido intestinal. En ella encontramos el plexo de<br />
Auerbach.<br />
Hay miocitos que se contraen, provocando dos tipos de contracciones:<br />
La capa interna forma contracciones de segmentación que movilizan el contenido luminal, haciendo que se<br />
mezclen con los jugos que se vierten al intestino, y aseguran que el jugo esté en contacto con la mucosa.<br />
La capa externa es responsable del peristaltismo, que permite el avance del contenido.<br />
Las ondas peristálticas se producen también en ayuno para que el intestino delgado se vacíe. La motilina<br />
favorece el peristaltismo. La motilina se segrega permanentemente cuando no <strong>com</strong>emos y se inhibe su<br />
liberación cuando <strong>com</strong>emos.<br />
Si no se dan movimientos peristálticos, se retiene alimento y crece la flora bacteriana.<br />
Esfínter Ileocecal<br />
El esfínter ileocecal se dispone entre el íleon y el ciego. Está parcialmente contraído. Esta contracción tónica<br />
está impidiendo que el contenido avance hacia el ciego. Si el ciego se distiende, entonces se produce, <strong>com</strong>o<br />
reacción refleja, una inhibición del peristaltismo del íleon y una contracción del esfínter.<br />
Tras una ingesta, lo que se produce es la relajación del esfínter, que permite el avance del contenido y que este<br />
alcance el ciego.<br />
No hay que confundir el esfínter con la válvula ileocecal: la válvula son pliegues de la mucosa orientados hacia<br />
el ciego, de tal forma que una vez que el ciego se llena, los pliegues se cierran, impidiendo que el contenido<br />
retroceda hacia el íleon.<br />
Serosa<br />
En el yeyuno y el íleon, pero no en el duodeno, se encuentra asociada al peritoneo, formando una serosa.<br />
Diferencias regionales<br />
Duodeno<br />
Encontramos vellosidades de tipo foliáceo, sobre todo en la infancia. Encontramos glándulas de Brunner. En el<br />
duodeno se produce la llegada de jugo pancreático y la bilis. No se encarga de la digestión final y permite la<br />
emulsión de las grasas.<br />
Íleon<br />
Es la zona donde se da un máximo desarrollo de las válvulas conniventes, vellosidades y microvellosidades. Las<br />
vellosidades son digitiformes. Las células presentan una alta tasa de división y es la zona donde se produce la<br />
máxima absorción.<br />
Yeyuno<br />
Tenemos un desarrollo máximo del tejido linfoide, y encontramos el esfínter y la válvula ileocecales. Se<br />
produce la absorción específica de sales minerales
Página57<br />
INTESTINO GRUESO, RECTO Y CONDUCTO ANAL<br />
El intestino grueso conforma la porción distal del tubo digestivo. Tiene un diámetro mayor que el intestino<br />
delgado (5 cm) y una longitud de 1,5 m. Anatómicamente se divide en colon, recto y ano. El ciego es donde se<br />
sitúa además el apéndice vermiforme.<br />
Entre las funciones del intestino grueso contamos las de absorción, secreción, <strong>com</strong>pactación de heces y<br />
almacenamiento temporal de las mismas.<br />
Las heces se <strong>com</strong>ponen en un 75% de agua, y el 25% restante de grasa, proteínas no digeridas, sustancias<br />
inorgánicas, células muertas y pigmento biliar (estercobilina). La flora bacteriana es anaeróbica, y <strong>com</strong>o<br />
consecuencia se ello se dan reacciones de putrefacción y fermentación que conllevan la producción de gas.<br />
En las heces encontramos vitaminas K y B12, ácido sulfhídrico y mercaptanos.<br />
Al igual que todo el tubo digestivo, el intestino grueso presenta una mucosa con epitelio, lámina propia y<br />
muscularis mucosa; una submucosa, una capa muscular y una adventicia.<br />
En la cara interna del intestino grueso encontramos los pliegues semilunares o válvulas colónicas, pliegues<br />
formados por mucosa que se contrae por acción de la muscularis mucosae. No encontramos válvulas<br />
conniventes ni vellosidades.<br />
En la cara externa del intestino grueso encontramos las haustras y los apéndices epiploicos, unas proyecciones<br />
serosas de tejido adiposo.<br />
Mucosa<br />
El epitelio de la mucosa del intestino grueso es prismático simple. Presenta escasas vellosidades, alguna en la<br />
parte más proximal. Encontramos abundantes criptas de Lieberkühn, más largas que en el intestino delgado. En<br />
el epitelio encontramos diversos tipos celulares: enterocitos, células caliciformes, células madre y células<br />
endocrinas.<br />
CÉLULAS ABSORTIVAS CILÍNDRICAS O ENTEROCITOS<br />
Son las células principales del intestino grueso, sobre todo en el colon (proporción 4:1 con caliciformes). La<br />
cantidad de estas células va disminuyendo, hasta que en el recto se encuentran en el mismo número que las<br />
células caliciformes (proporción 1:1).<br />
Estos enterocitos son muy similares a los del intestino delgado: su membrana apical sigue teniendo<br />
microvellosidades, pero no son tan numerosas y regulares <strong>com</strong>o en el intestino delgado, por lo que no se<br />
observa chapa estriada. Estos enterocitos presentan una mayor proporción de mitocondrias.<br />
El espacio intercelular está muy dilatado. Esta región acoge agua, sales y vitaminas que los enterocitos<br />
absorben. En el intestino grueso, el transporte activo de Na+ arrastra mucho agua, lo que es fundamental para<br />
que las heces sean cada vez más semisólidas. Los enterocitos absorben asimismo determinados fármacos.<br />
Los enterocitos segregan además K + y HCO 3 - , secreción que presenta un control endocrino por aldosterona.<br />
CÉLULAS CALICIFORMES<br />
Las células caliciformes son aquí más numerosas que en el intestino delgado, encontrándose su máxima<br />
densidad en las criptas de Lieberkühn.<br />
Las células caliciformes segregan de manera constante un producto mucoso que juega un papel fundamental<br />
para proteger la mucosa frente al contenido luminal que se solidifica y erosiona la pared. El moco facilita<br />
asimismo el avance de las heces y su <strong>com</strong>pactación, por lo que tienen mayor concentración según avanzamos<br />
hacia el recto.
Página58<br />
Las células con flecos caveolada es una célula caliciforme agotada que ha expulsado casi todo su contenido de<br />
mucinógeno, de modo que son altas y delgadas con escasos gránulos de moco en su interior y un citoplasma<br />
centropical.<br />
CÉLULAS MADRE<br />
Se sitúan en la base de las criptas de Lieberkühn y presentan un índice de renovación similar a las del intestino<br />
delgado<br />
CÉLULAS ENTEROENDOCRINAS<br />
Estas células son menos numerosas que en el intestino delgado, y se encuentran principalmente en el colon, en<br />
la parte inferior de las criptas.<br />
En el intestino grueso, las células endocrinas segregan glucagón, sustancia P, cromogranina, somatostatina,<br />
encefalinas, histamina, péptido Y, VIP, GLP-1 (similar a glucagón 1)…<br />
La somatostatina actúa estimulando la síntesis de gastrina y la secreción ácida gástrica.<br />
GLP-1 inhibe la secreción acida y el vaciado gástrico. Estimula la liberación de insulina.<br />
El péptido Y inhibe el vaciado gástrico y la secreción ácida.<br />
Lámina Propia<br />
Es un conjuntivo laxo con un espacio de conjuntivo: la meseta colágena. Esta meseta tiene un grosor de 5 μm y<br />
está situada entre la lámina basal subeptielial y las láminas basales de los capilares fenestrados. Es una zona<br />
que regula el transporte de agua y sales que llegan hasta la sangre desde los enterocitos. La meseta colágena<br />
está muy hipertrofiada en personas con pólipos en el colon.<br />
La lámina propia presenta una importante cantidad de fibroblastos alrededor de las criptas formando la Vaina<br />
Fibroblástica Pericríptica. Estas células se dividen con gran frecuencia y migran en paralelo a las criptas hacia la<br />
superficie. Lo curioso es que estos procesos ocurren en sincronización con la división de las células madre de<br />
las bases de las criptas: es por ello que se postula que estos fibroblastos que forman la vaina dan lugar a los<br />
macrófagos de la lámina propia.<br />
La lámina propia presenta un desarrollo muy marcado del GALT con grandes nódulos linfáticos que distorsionan<br />
la lámina propia y se extienden hacia la submucosa. No encontramos vasos linfáticos, lo cual se relaciona con el<br />
hecho de que en buena parte del colon hay un bajo índice de metástasis.<br />
Muscularis Mucosae<br />
Presenta un estrato interno, con los miocitos dispuestos circularmente; y otro externo, con los miocitos<br />
dispuestos longitudinalmente. Desaparece bruscamente al llegar al conducto anal. Sí encontramos asociados a<br />
los miocitos una red de linfáticos.<br />
Submucosa<br />
Es un conjuntivo laxo muy vascularizado, con una alta presencia de adipocitos.<br />
Capa Muscular<br />
El estrato circular interno es similar al del intestino delgado. En el conducto anal forma el esfínter anal interno.<br />
El estrato longitudinal externo solo existe en parte del intestino grueso: en el recto, el conducto anal y el<br />
apéndice. En el ciego y el colon no hay capa longitudinal externa y los miocitos longitudinales se agrupan<br />
formando tres bandas: las tenias del colon.<br />
Los miocitos salen de la tenia y pasan al estrato circular interno de techo en trecho, haciendo que se observen<br />
segmentos: los sacos o haustras. Estos sacos tienen unas contracciones locales, independientes entre sí.<br />
La capa longitudinal provoca el peristaltismo, que permite el avance del contenido hacia el ano, normalmente<br />
una vez al día.
Página59<br />
Adventicia<br />
En algunas zonas es una serosa. Presenta unas proyecciones asociadas con adipocitos: los apéndices epiploicos.<br />
Ciego<br />
Es una expansión del intestino grueso que termina cerrado. Es una zona de máxima densidad glandular y de<br />
criptas, con una mayor cantidad de caliciformes. El ciego presenta tenias y tiene un importante desarrollo del<br />
GALT.<br />
Apéndice Vermiforme<br />
Es una expansión que sale del ciego. Mide unos 10 o 15 cm, y está situado a 3 cm. del íleon. Su luz pequeña e<br />
irregular, con restos acumulados, y generalmente está ocluida en adultos. En una preparación de apéndice no<br />
hay chapa estriada y hay pocas criptas, bastante irregulares. Se observan pocas células caliciformes y una<br />
norme cantidad de GALUs y linfocitos dispersos, que parecen formar una capa continua de nódulos, haciendo<br />
del apéndice un órgano linfoide secundario.<br />
La muscularis mucosae está poco definida por el desarrollo masivo del GALT. La submucosa, aunque sea laxa,<br />
tiende con los años a hacerse más densa y fibrosa. La capa muscular es delgada, sin tenias.<br />
El apéndice puede sufrir episodios inflamatorios crónicos, típicos de infancia y la adolescencia, menos probables<br />
en adultos. La apendicitis aguda es una obstrucción de la luz y una ulceración de la mucosa. El aumento de la<br />
presión intraluminal conlleva una dilatación de los vasos, lesionándolos. Se puede producir perforación y<br />
peritonitis.<br />
Colon<br />
Se divide en colon ascendente, colon transverso y colon descendente. En él, el material luminal se hace<br />
semisólido, sobre todo en el colon descendente y el colon sigmoideo. Tiene tenias y apéndices epiploicos.<br />
Funcionalmente, tiene una zona donde se produce una mayor absorción de agua y electrolitos, produciéndose<br />
la <strong>com</strong>pactación de las heces. Esta propiedad se utiliza para administrar fármacos (sedantes, anestésicos…) en<br />
forma de supositorios.<br />
Recto<br />
Va desde el colon sigmoide hasta el diafragma pélvico y suele estar vacío habitualmente. Su mucosa es<br />
bastante gruesa, con criptas de amplia longitud y gran cantidad de células caliciformes. La parte más superior<br />
del recto muestra unos pliegues de la mucosa: los pliegues rectales transversos. En la parte inferior muestra un<br />
mayor diámetro (ampolla rectal) y encontramos los pliegues semilunares.<br />
Conducto Anal<br />
Va desde el diafragma pélvico hasta el orificio anal. Presenta una serie de pliegues longitudinales, las columnas<br />
anales o de Morgagni. En la base de estos pliegues tiene una serie de expansiones, las válvulas anales. Entre<br />
válvula y válvula se observan las depresiones llamadas senos anales.<br />
Los senos y las columnas actúan conjuntamente para impedir fugas: cuando las heces avanzan, las columnas y<br />
válvulas se aplanan para permitir el paso de las heces.<br />
En el conducto anal no hay tenias, y la capa circular interna forma el esfínter anal interno, que cierra el paso a<br />
las heces. Existe otro esfínter, el anal externo, que es de músculo esquelético<br />
Se necesita sincronía entre aplanamiento de columnas y senos, la secrecion y el control sobre lo esfínteres.<br />
La línea pectínea es una línea imaginaria que trazamos uniendo las bases de las columnas anales. Es la zona de<br />
división entre ectodermo y endodermo, es decir, es la zona donde se situó primigenia membrana anal.
Página60<br />
El conducto anal presenta tres zonas diferentes:<br />
‣ Porción Colorrectal o Tercio Superior: tapizada por epitelio prismático simple.<br />
‣ Tercio medio o de Transición: es una región especial que permite el paso del epitelio simple cilíndrico de la<br />
mucosa rectal al epitelio estratificado plano de la piel perianal, de modo que aparece un epitelio cilíndrico<br />
estratificado.<br />
‣ Porción Pavimentosa o Tercio Inferior: tapizado por un epitelio simple plano que se continúa con la piel de<br />
la región perianal.<br />
Las criptas de Lieberkühn <strong>com</strong>ienzan a ser cada vez más cortas y, 2 cm antes del orificio anal, en la línea<br />
pectínea, desaparecen. En el conducto anal no encontramos muscularis mucosae.<br />
Glándulas Anales<br />
Son glándulas tubulares rectas ramificadas, se ubican en la submucosa, y pueden invadir la muscular del<br />
esfínter anal interno. Son glándulas mucosas y drenan por un conducto de epitelio prismático estratificado que<br />
se abre en un seno anal y suelen estar rodeadas de tejido linfoide difuso. Esta secreción se vierte antes de que<br />
pasen las heces, de manera que estimula el vaciado del canal anal. Estas glándulas se infectan generando<br />
abscesos y fistulas.<br />
En la porción distal del conducto anal, encontramos glándulas sebáceas y perianales (sudoríparas apocrinas).<br />
Vascularización<br />
Encontramos estructuras vasculares (glomérulos vasculares) que reciben sangre arterial y venosa. Están entre<br />
la lámina propia y la submucosa. La sangre acaba llegando a una zona que denominamos cuerpo cavernoso<br />
rectal. Cuando se llena de sangre, este cuerpo contribuye a cerrar el paso a las heces.<br />
Si estos vasos sanguíneos se dilatan excesivamente, se conduce a hemorroides internas, asociadas a<br />
hipertensión portal frecuentemente. Las hemorroides externas se producen por la dilatación de venas cutáneas<br />
del orificio anal.
Página61<br />
HÍGADO<br />
El hígado es una glándula mixta, con <strong>com</strong>ponente endocrino y exocrino. Es la mayor glándula corporal, con un<br />
peso aproximado de 1,5 kg, y se la puede dividir en dos lóbulos principales y dos lóbulos menores unidos a<br />
nivel del hilio.<br />
El parénquima es de origen endodérmico, surge de una evaginación del intestino anterior que, además del<br />
divertículo hepático, genera el conducto colédoco, es decir, tanto el conducto cístico <strong>com</strong>o el hepático.<br />
Estroma<br />
El estroma del hígado es de origen mesodérmico<br />
El estroma hepático forma, en el exterior la cápsula de Giesson, un conjuntivo denso y fino cubierto de<br />
mesotelio desde el cual surgen tabiques que forman los lóbulos y lobulillos hepáticos. Dentro de los lobulillos,<br />
encontramos una red de fibras reticulares estromales, pero nunca fibras colágenas<br />
El espacio portal o de Kiernan es un estroma de conjuntivo laxo con fibras de colágeno I y escasas células. Aquí<br />
encontramos vasos linfáticos, fibras amielínicas simpáticas y parasimpáticas y, sobre todo, la tríada portal,<br />
formada por una rama de la vena porta, otra de la arteria hepática y una rama del conducto biliar.<br />
Organización Estructural Hepática<br />
Los hepatocitos se disponen en cordones, formando las trabéculas de Remak, una red tridimensional dispuesta<br />
desde la vena centrolobulillar, paralela a los sinusoides hepático. Al espacio mínimo que queda entre la pared<br />
del sinusoide y la membrana del hepatocito de los cordones se le denomina espacio de Disse.<br />
Lobulillo Hepático o Lobulillo Clásico<br />
Es la división clásica del hígado descrita por Maplighi y se basa en la distribución de las ramas de la vena portal<br />
y la arteria hepática dentro del propio órgano.<br />
Consiste en pilas de trabéculas de hepatocitos anastomosadas separadas por un sistema anastomosado de<br />
sinusoides que perfunden el tejido. Tienen morfología poliédrica o hexagonal, muy fáciles de ver en el hígado<br />
de cerdo, en cuyo centro está la vena hepática terminal o central a la que drenan los sinuosides. En otras<br />
especies <strong>com</strong>o el hombre, los lobulillos no son tan evidentes. En el borde del lobulillo tenemos los canales o<br />
espacios portales o de Kiernan <strong>com</strong>puestos por tejido conectivo laxo en la que se sitúan las triadas portales y<br />
que se continúa con la cápsula fibrosa hepática.<br />
El lobulillo está limitado por los hepatocitos más externos del lobulillo. Además, en los bordes del espacio<br />
portal, entre el estroma de tejido conectivo y los hepatocitos, aparece el espacio de Mall donde se origina,<br />
entre otros sitios, la linfa en el hígado.<br />
Lobulillo Portal<br />
Mall estableció que el hígado se <strong>com</strong>ponía de unas unidades poliédricas, que son los lobulillos portales. Para<br />
Mall, el hígado era una glándula y las subunidades que estableció eran glandulares, es decir, son unidades<br />
secretoras de bilis, de modo que la bilis fluye hacia el centro de este lobulillo portal. Así el eje morfológico es el<br />
conduco biliar interlobulillar y los márgenes externos lo conforman líneas imaginarias trazadas entre las 3<br />
venas centrales más cercanas a la tríada portal. No hay un límite claro de estos lobulillos a MO.<br />
Esquema de las 3 Organizaciones
Página62<br />
Ácino Hepático o de Rappaport<br />
Esta unidad fue establecida por Rappaport. Tiene forma romboidal y una delimitación no muy clara. El eje<br />
menor del acino está definido por las ramas terminales de la tríada portal localizada a lo largo del límite de dos<br />
lobulillos clásicos y el eje mayor es la línea trazada entre las dos venas centrales más cercanas al eje menor.<br />
Esta concepción es más moderna, donde las unidades hepáticas son metabólicas, no anatómicas ni glandulares.<br />
Presentan tres zonas (1, 2 y 3) según la cercanía al eje.<br />
La zona 1 es la más cercana al eje y recibe preferentemente los nutrientes y otros productos de la circulación.<br />
Tardan más en morir ante deficiencia de nutrientes y son las primeras zonas en recuperarse tras un daño<br />
hepático, así <strong>com</strong>o las primeras en mostrar alteraciones biliares.<br />
La zona 3 está más alejada del eje y tarda más en manifestar los síntomas de intoxicación y responder a la<br />
estasis biliar, pero es la que antes se afecta ante una hipoxia (necrosis en isquemia) y en acumular lípidos.<br />
La zona 2 se encuentra entre las zonas 1 y 2 sin límites bien definidos. Presentan diferencias en la actividad<br />
enzimática, cantidad de glucógeno y de organelas.<br />
El ácino hepático es una concepción funcional, que permite interpretar mejor datos de modelos de<br />
degeneración, regeneración e intoxicación hepáticas.<br />
En una insuficiencia cardiaca congestiva, que provoca una baja perfusión y por tanto una hipoxia de los<br />
órganos, la zona 3 del ácino hepático es la que antes se afecta ante una hipoxia. Se desarrolla entonces cirrosis<br />
cardiaca en el hígado y necrosis centrolobulillar isquémica, donde el hepatocito presenta vacuolas lipídicas y<br />
luego muere.<br />
Irrigación<br />
Esquema del ácino hepático<br />
Un 75% de la sangre que llega al hígado es aportada por la vena porta hepática, con una sangre color rojo<br />
oscuro por su gran cantidad de nutrientes. Esta sangre recoge los <strong>com</strong>ponentes endocrinos de la pared del<br />
tubo digestivo y del páncreas. De esta vena porta hepática se dan ramificaciones interlobulares, conductoras,<br />
interlobulillares, hasta llegar a las venas portas, en los espacios portales.<br />
El 25% de la sangre restante lo lleva la arteria hepática, con sangre oxigenada, que se ramifica también en<br />
ramas interlobulares e intralobulares, hasta las ramas terminales, en los espacios portales.<br />
Desde los espacios portales la sangre mezclada pasa a los sinuosides. Estos sinusoides están formados por<br />
células endoteliales, células de Kupffer y células de Ito, lipocitos o almacenadoras de grasa.<br />
La sangre fluye hacia las venas centrolobulilalres, que convergen entre sí para formar venas sublobares o<br />
intercalares, colectoras y hepáticas, hasta alcanzar la cava inferior.<br />
El endotelio está formado por células epiteliales muy delgadas. El endotelio corresponde a capilares<br />
fenestrados con grandes brechas, capilares que se apoyan en la lámina basal, de modo que es un conducto de<br />
pared muy fina por la que se sale la sangre hacia los espacios de Disse. Los elementos formes y los<br />
quilomicrones quedan retenidos en los sinusoides.
Página63<br />
CÉLULAS DE KUPFFER<br />
Se detectan por tinción argéntica y cloruro de oro.<br />
Son células de morfología estrellada que provienen de macrófagos (monocitos). Están formando parte de la<br />
pared del sinusoide, sin unirse a las células endoteliales, y son capaces de emitir pseudópodos y moverse. Se<br />
pueden dividir de forma activa.<br />
Las células de Kupffer se encargan de degradar eritrocitos y almacenan hierro, funciones que resultan<br />
esenciales tras una esplenectomía.<br />
Además, estas células tienen que ver con la función inmunológica del hígado: segregan citocinas ante la<br />
estimulación producida por ciertos agentes (alcohol, infecciones), produciendo una reacción inflamatoria.<br />
CÉLULAS DE ITO<br />
Estas células se ven por tinción de cloruro de oro. Son células estrelladas de origen mesenquimal que<br />
presentan inclusiones con lípidos y vitamina A, la cual almacenan y movilizan cuando es necesaria.<br />
Ciertos estudios demostraron que, en determinadas situaciones, estas células se transforman y cumplen otras<br />
funciones. Al transformarse, se convierten en miofibroblastos, que por una parte son contráctiles y por otra<br />
parte sintetizan fibras colágenas I y III (fibras reticulares). Esto provoca que el hígado se hace cada vez más<br />
fibroso, pudiendo provocar una cirrosis.<br />
En condiciones normales, las células de Ito regulan el diámetro de los sinusoides gracias a su potencial<br />
capacidad contráctil.<br />
Las células de Ito juegan un papel importante cuando, tras una lesión hepática, <strong>com</strong>ienza una regeneración<br />
remodelando la matriz extracelular.<br />
Espacio Perisinusoidal o de Disse<br />
El espacio de Disse queda entre la pared del sinusoide y la membrana plasmática de los hepatocitos. Estos<br />
espacios son continuos y se encuentran en la zona periférica del lobulillo clásico, es decir, en el espacio portal.<br />
Los espacios de Disse convergen formando el espacio de Mall, una de las zonas donde se supone se genera<br />
linfa. Los primeros capilares linfáticos se sitúan en el espacio portal, es decir, la linfa fluye en la misma<br />
dirección que la bilis. La linfa drena finalmente al conducto torácico.<br />
El espacio de Disse es el espacio intercelular que queda entre el endotelio sinusoidal y la membrana plasmática<br />
del hepatocito. Debido a que el sinusoide carece de lámina basal y a que posee grandes brechas, prácticamente<br />
no hay barrera entre el plasma del vaso y este espacio, por lo que la sangre se encuentra en contacto libre con<br />
la cara del hepatocito orientada a este espacio.<br />
En este espacio casi no hay elementos de matriz,<br />
pocas fibras de reticulina y algún axón amielínico.<br />
Hay sobre todo plasma que fluye hacia la zona<br />
periférica, hacia el espacio de Mall. En este espacio<br />
de Disse encontramos las microvellosidades de las<br />
membranas de los hepatocitos. El espacio de Disse<br />
es la zona en donde se produce la hematopoyesis<br />
durante la etapa fetal. En adultos con anemias<br />
crónicas, se ha demostrado que las células<br />
hematopoyéticas de esta zona se recuperan.
Página64<br />
Hepatocitos<br />
Los hepatocitos son células de origen epitelial, están polarizadas, y tienen una vida media de 4-6 meses, a lo<br />
largo de los cuales pueden dividirse, aunque no es habitual. El hepatocito tiene una alta capacidad<br />
regenerativa: en situación de daño, esta capacidad se pone de manifiesto y se recupera la masa hepática<br />
rápidamente.<br />
Los hepatocitos son células grandes (20-30μm), con núcleo grande y posición central. Se encuentran<br />
frecuentemente polinucleados (binucleados) y suponen el 80% de la población celular del hígado. Tienen un<br />
citoplasma eosinófilo con todo tipo de orgánulos.<br />
Presentan inclusiones lipídicas se ponen especialmente de manifiesto en el hígado graso. Presenta asimismo<br />
inclusiones de glucógeno (PAS+), en una cantidad que presenta variación circadiana.<br />
El hepatocito presenta abundantes mitocondrias y un gran desarrollo del citoesqueleto, que forma una red<br />
bastante más densa en algunas zonas, <strong>com</strong>o por ejemplo alrededor de la superficie canalicular. El hepatocito<br />
presenta una gran cantidad de RER, que se pone de manifiesto a microscopía óptica <strong>com</strong>o grumos de Berg.<br />
Su aparato de Golgi es muy grande, con hasta 50 dictiosomas, relacionados con la secreción de bilis y VLDL.<br />
El REL del hepatocito es fundamental en degradación de tóxicos, fármacos, proteínas no hidrosolubles,…<br />
Asimismo, está relacionado con la síntesis de colesterol y sales biliares, la degradación de glucógeno, la<br />
degradación de ácidos grasos paraformar triglicéridos y la de toxificación. Otra de sus funciones es la<br />
eliminación de yodo. El REL es fundamental asimismo en el metabolismo del etanol mediante dos vías:<br />
- Por un lado, ADH es una enzima que actúa sobre el etanol, obteniéndose protones y acetaldehído.<br />
Ambos, cuando se forman en grandes cantidades, producen daño hepático: dañan las mitocondrias y el<br />
citoesqueleto y además generan una respuesta autoinmune.<br />
- La segunda vía es la vía MEOS. En esta vía, el etanol se metaboliza dando acetaldehído y radicales de<br />
oxígeno, muy lesivos para la membrana plasmática del hepatocito. El hepatocito reacciona provocando<br />
una reacción autoinmune y perdiendo su capacidad de resistir los residuos de etanol.<br />
Encontramos en el hepatocito una importante cantidad de pigmentos y lisosomas con Fe, Cu y lipofucsina. La<br />
cantidad de estos lisosomas aumenta en ciertas patologías: anemia, hepatitis, estasis biliar…<br />
Los peroxisomas son orgánulos a los que muchas veces se les da poca importancia, pero que son<br />
fundamentales en el hígado. Metabolizan ciertos radicales y tóxicos, de modo que son fundamentales en la<br />
función de detoxificación, junto con el REL. Intervienen en la β-oxidación, en la gluconeogénesis y en el<br />
metabolismo de las purinas.<br />
Existen algunas enfermedades relacionadas con los peroxisomas:<br />
· Enfermedad de Zellweger: consiste en una ausencia de las enzimas de los peroxisomas, que están vacíos.<br />
· Adenoleucodistrofia: es una enfermedad ligada al cromosoma X, y provoca la degeneración de la mielina.<br />
En el hígado encontramos los hepatocitos adosados en forma de cordones. La mayor parte de la membrana<br />
plasmática de los hepatocitos da al espacio de Disse, formando la superficie del sinusoide hepático. Esta<br />
membrana tiene abundantes microvellosidades, lo que permite aumentar la superficie de intercambio con la<br />
sangre.<br />
El resto de la membrana plasmática se divide entre la superficie intercelular (15%), adosada a otro hepatocito,<br />
regular y continua, con uniones de <strong>com</strong>unicación; y la superficie canalicular (15%), delimitando los canalículos<br />
biliares que se abren entre los hepatocitos.<br />
Estos canalículos tienen una anchura de 1 μm. y muestran microvellosidades. Encontramos abundantes<br />
fosfatasas que se encargan de intercambiar productos y excretar la bilis, puesto que el canalículo es la primera<br />
vía de la secreción biliar. Esta membrana plasmática tiene una serie de transportadores: son cuatro de la<br />
familia ABC, encargados del transporte específico de moléculas. MDR1 transporta colesterol; MDR2,<br />
fosfolípidos; MOAT, bilirrubina conjugada; y BAT, sales biliares. Es fácil encontrar mutaciones en los genes de<br />
estos transportadores, afectando a la secreción de la bilis (la bilis no tiene el <strong>com</strong>ponente que transporta ese<br />
transportador). Se afecta no solo la <strong>com</strong>posición, sino que se producen necrosis focales que provocan la<br />
proliferación de los conductillos biliares e inflamación.<br />
En los hepatocitos se crea un gradiente osmótico de ácidos biliares, por lo que el hepatocito tiende a expulsar<br />
sangre al canalículo, lo que permite el paso de bicarbonato y cloro al mismo.
Página65<br />
Estos canalículos tienen que estar perfectamente sellados, por lo que hay una importante cantidad de<br />
<strong>com</strong>plejos de unión y uniones de adhesión. Rodeando al canalículo encontramos un gran desarrollo de<br />
filamentos de actina y miosina en el hepatocito. Estos filamentos juegan un papel fundamental en la regulación<br />
del diámetro del canalículo (0.5-1 μn.), controlando el avance la bilis. La Amanita phalloides inhibe este sistema<br />
contráctil.<br />
Bilis<br />
Se segrega 1 l. diario de bilis. La bilis se <strong>com</strong>pone de agua, electrolitos (K+, Na+,<br />
Ca2+, Mg2+, Cl-…), fosfolípidos y colesterol, ácidos biliares primarios y secundarios, pigmentos biliares y<br />
bilirrubina.<br />
Funciones:<br />
· Excreción de los <strong>com</strong>ponentes de la bilis<br />
· Excreción de catabolitos que proceden de metales pesados o fármacos procesados por el hepatocito<br />
· Facilita la absorción de las grasas, ya que permite su emulsión<br />
· Transporte de IgA a la mucosa intestinal (circulación enterohepática)<br />
· Inhibición de la flora bacteriana por ácidos biliares<br />
Árbol Biliar<br />
Las vías de excreción de la bilis crecen en diámetro y <strong>com</strong>plejidad de la pared según avanzan.<br />
Canalículos Biliares o Capilares<br />
Son los canales presentes entre los hepatocitos, de luz entre 0,5 y 1,5 μm., y donde podemos encontrar<br />
microvellosidades, la fosfatasa alcalina y uniones de zonula occludens. Cerca del espacio portal se reúnen para<br />
formar los colangiolos. En los canalículos la membrana plasmática de los hepatocitos hay una ATPasa que<br />
sugiere que la secreción de bilis a este espacio es un proceso activo.<br />
Conductillo Biliar, Colangiolo o Conducto de Hering<br />
Es una vía corta (1 μm), con un epitelio cúbico simple, con lámina basal continua, <strong>com</strong>o en el resto del árbol<br />
biliar. Recibe la bilis desde los canalículos y conduce la bilis a los conductos biliares interlobulillares, es decir, a<br />
la triada portal.<br />
Conductor Biliares Interlobulillares<br />
Están dentro de las triadas portales. Es la vía más grande (hasta 40 μm) y está tapizada por un epitelio simple<br />
cilíndrico alto. Su membrana apical tiene microvellosidades, bajo la lámina basal se observa tejido conjuntivo<br />
denso. En algunas zonas próximas al hilio se detectan también fibras musculares.<br />
Vías Extrahepáticas<br />
La conforman los conductos hepáticos izquierdo y derecho y el conducto hepático <strong>com</strong>ún. Este último tiene 3<br />
cm. de longitud, con una pared enteramente similar a la del tubo digestivo, solo que no hay muscularis<br />
mucosa. Está tapizada por epitelio prismático simple que recuerda al de la vesícula biliar.<br />
Conducto Cístico<br />
Conecta la vesícula con el hepático <strong>com</strong>ún. En su pared se encuentran todas las capas de la vesícula biliar. La<br />
mucosa tiene pliegues que constituyen la válvula espiral o de Heister, que controla la entrada y salida de bilis a<br />
la vesícula.
Página66<br />
Colédoco<br />
El colédoco permite la salida de la bilis al duodeno y recoge secreción pancreática del conducto de Wirsung. Se<br />
dilata en el duodeno, formando la ampolla de Vater. La salida está controlada por el esfínter de Oddi.<br />
Esfínter de Oddi<br />
El esfínter de Oddi engloba varias estructuras:<br />
o El esfínter más proximal se denomina esfínter de Boyden, e inhibe el paso de la bilis al duodeno<br />
o El esfínter pancreático es el encargado de inhibir el paso de jugo pancreático al intestino delgado y de la<br />
bilis al conducto pancreático.<br />
o El esfínter de la ampolla se <strong>com</strong>pone de miocitos longitudinales que favorecen la entrada de la bilis y el<br />
jugo pancreático al duodeno.<br />
El flujo biliar depende de los hepatocitos, que segregan continuamente bilis, generando una presión; y del<br />
esfínter de Oddi. En ayuno, el esfínter está cerrado, por lo que la bilis es conducida a la vesícula. La ingesta<br />
produce incremento de CCK que tiene dos efectos: estimula la contracción de la musculatura vesical y relaja el<br />
esfínter de Oddi. Esto permite que la bilis almacenada llegue al duodeno.<br />
Funciones<br />
El hígado es una glándula que surgió evolutivamente <strong>com</strong>o reservorio de moléculas energéticas, y<br />
posteriormente pasó, en los mamíferos, a secretar bilis y convertirse en el órgano central del metabolismo y la<br />
homeostasis corporal. Sus funciones son:<br />
· Captación, almacenamiento y distribución de sustancias nutritivas.<br />
· Almacenamiento de vitaminas<br />
· Almacenamiento, metabolismo y homeostasis del hierro<br />
· Síntesis de proteínas plasmáticas circulantes<br />
· Función inmunológica<br />
· Degradación de proteínas<br />
· Secreción de bilis<br />
· Función endocrina<br />
En el hígado se producen una serie de reacciones con respecto a distintas hormonas y reguladores:<br />
· La tiroxina T4 se convierte en T3 o forma activa<br />
· Se produce la síntesis de somatomedina C o IGFI, implicada en la síntesis de GH<br />
· La insulina y el glucagón cumplen su función y son degradados<br />
· La vitamina D (hormona) se transforma en 25-hidroxicolecalciferol (forma funcional).<br />
Por un lado, el hígado se encarga del mantenimiento de la glucemia, ya que almacena glucógeno (por lo que es<br />
PAS+), lo degrada en situaciones de ayuno y, en caso de una necesidad aún mayor de glucosa, realiza el proceso<br />
de gluconeogénesis a partir de precursores aminoacídicos.<br />
Además, el hígado juega un papel fundamental en el metabolismo lipídico. Por ejemplo, es capaz de actuar<br />
sobre los ácidos grasos captados mediante la reacción de β-oxidación, formando cuerpos cetónicos. Un exceso<br />
de estos cuerpos cetónicos (por ejemplo, en casos de inanición), causa problemas en el sistema nerviosos en lo<br />
que se conoce <strong>com</strong>o cetosis.<br />
Asimismo, el hígado es responsable de la síntesis y captación del colesterol, la síntesis de sales biliares y su<br />
excreción a la bilis, así <strong>com</strong>o de la síntesis de lipoproteínas transportadoras <strong>com</strong>o VLDL, HDL o LDL. También<br />
regula los niveles de VLDL en sangre.<br />
El hígado sintetiza urea <strong>com</strong>o mecanismo de eliminación del amonio que surge de la desaminación de los<br />
aminoácidos y del metabolismo bacteriano en el tubo digestivo.<br />
Una alteración de esta síntesis de urea puede provocar un <strong>com</strong>a hepático.<br />
Finalmente, el hígado tiene la capacidad de sintetizar todos los aminoácidos esenciales y de convertir los no<br />
esenciales en esenciales, incrementando así la capacidad del organismo de producir proteínas.
Página67<br />
El hígado constituye un reservorio de vitaminas A (para 10 meses), D (para 4 meses) y B12 (para un año).<br />
En el hígado se produce el almacenamiento, captación y mantenimiento de los niveles circulantes de vitamina<br />
A, precursora del retinal. Este retinal se transforma en retinol unido a la proteína RBP gracias a las células de<br />
Ito. Un fallo de esta función puede provocar ceguera nocturna y trastornos de la piel.<br />
En cuanto a la vitamina D, aunque el hígado no la almacena es fundamental en su metabolismo, en el que son<br />
cruciales los metabolismos del calcio y del fósforo. En el hígado la vitamina D3 se convierte en 25-<br />
hidroxicolecalciferol. Un fallo de esta función produce raquitismo y trastornos de mineralización.<br />
Finalmente, en lo que respecta a la vitamina K, en el hígado se capta, se almacena y se regulan sus niveles en<br />
sangre. La vitamina K es fundamental en la síntesis de protrombina y otros factores de coagulación. La<br />
alteración de esta función genera hipoprotrombinemia y trastornos hemorrágicos.<br />
El hígado sintetiza las proteínas implicadas en el metabolismo y transporte del hierro, tales <strong>com</strong>o la<br />
transferrina, la haptoglobina y la hemopexina. Además, almacena el hierro en forma de ferritina o <strong>com</strong>o<br />
gránulos de hemosiderina.<br />
El hígado se encarga de la síntesis de globulinas no inmunológicas (_ y _) y albúmina, importantes para el<br />
mantenimiento de la presión coloidosmótica, del control del volumen plasmático y, en última instancia, del<br />
equilibrio hídrico corporal.<br />
Asimismo, en el hígado se generan proteínas de transporte: lipoproteínas (VLDL, HDL, LDL), transportadores del<br />
hierro (haptoglobinas, transferrina y hemopexina), transportadores de proteínas y ácidos grasos (albúmina) y<br />
proteínas implicadas en la cascada de coagulación <strong>com</strong>o la protrombina y el fibrinógeno.<br />
En el hígado se produce la síntesis de <strong>com</strong>plejos de IgA y <strong>com</strong>ponente secretor por parte de los hepatocitos,<br />
secretándolos a la bilis y a la sangre.<br />
Además, el hígado tiene la capacidad de reconocer y fagocitar hasta el 90% de las bacterias que circulan por la<br />
sangre portal, debido a la acción de las células de Kupffer, las cuales también eliminan células muertas y tienen<br />
cierta capacidad de destruir eritrocitos dañados.<br />
En el hígado se produce la degradación de fármacos, toxinas y proteínas extrañas, no hidrosolubles y por tanto<br />
no eliminables en la orina. Esta función corre a cargo del hepatocito, para lo que llevan a cabo un proceso de<br />
detoxificación en el que se produce la oxidación de estos <strong>com</strong>puestos en el REL y las mitocondrias, función en<br />
la que es muy importante el citocromo p450; y posteriormente se produce la conjugación mediante la unión a<br />
ácido glucurónico, glicina o taurina en los peroxisomas, de modo que estas proteínas ya pueden ser eliminadas<br />
por la orina.<br />
El consumo continuado de fármacos, sobre todo de barbitúricos, reduce la eficacia de estos fármacos, lo que<br />
hace que las dosis necesarias para que tengan efecto sean mayores. Del mismo modo, se produce<br />
progresivamente un daño del hepatocito, en el cual se ha hipertrofiado el REL e incrementado el número de<br />
oxidasa, lo que genera una tolerancia a fármacos.<br />
En el hígado se produce también la degradación de hemoglobina tras su destrucción en el bazo y en el hígado,<br />
gracias a las células de Kupffer. Posteriormente, la bilirrubina proveniente del grupo hemo se transporta unida<br />
a la albúmina hasta el hepatocito, el cual la conjuga y la elimina vía orina y vía heces.<br />
La ictericia se produce debido a niveles altos de bilirrubina libre o conjugada. Esto puede deberse a una<br />
hepatitis hemolítica, infecciosa o destructiva.<br />
La bilis es un medio de excreción digestiva en la que se secretan las sales biliares, sintetizadas por el propio<br />
hepatocito. La ausencia de sales biliares impide la emulsión y absorción de grasas, generando heces grasas<br />
(esteatorrea).
Página68<br />
VESÍCULA BILIAR<br />
La vesícula biliar es un órgano hueco, alargado, derivado secundario del intestino anterior embrionario, con<br />
capacidad de 50-70mL, que anatómicamente se divide en cuerpo, fondo y cuello, porción esta última que se<br />
continúa con el conducto cístico. La válvula de Heister se inicia en el cuello e impide que, ante cambios bruscos<br />
de presión, las paredes vesicales y del conducto se dañen.<br />
La bilis se almacena temporalmente en la vesícula, pero la vesícula además concentra la bilis 10 veces. La<br />
vesícula libera la bilis en respuesta a un estimulo, siendo el principal estímulo la ingesta. Asimismo, la vesícula<br />
cumple un importante papel en al regulación hidrostática del árbol biliar.<br />
La vesícula biliar consta de tres capas: mucosa, muscular y adventicia<br />
Mucosa<br />
La mucosa tiene una superficie irregular con muchas proyecciones llamadas arrugas o pliegues, cubiertas por<br />
una capa de moco protector. Estos pliegues se modifican, respondiendo al estado de distensión. En<br />
contracción, encontramos pliegues paralelos, altos y numerosos. En distensión, la atura y el número de<br />
pliegues se reduce.<br />
La mucosa se invagina, formando los senos de Rokitansky-Aschoff. Estos senos presentan un epitelio similar al<br />
de revestimiento. Los senos son abundantes en la zona del cuello y no hay que confundirlos con glándulas.<br />
Pueden invadir la muscular Hay quien piensa que los senos de Rokitansky son preludios de situaciones<br />
patológicas, porque en patologías se incrementa su número, si bien una vesícula con senos no indica patología.<br />
En los senos se acumulan bacterias.<br />
Es un epitelio simple cilíndrico, sin células caliciformes. Tiene células principales y células endocrinas.<br />
Las células principales presentan una membrana plasmática con muchas microvellosidades, con un glicocálix<br />
asociado (PAS+). Esta membrana apical tiene una importante cantidad de transportadores y bombas ATPasa.<br />
Presentan <strong>com</strong>plejos de unión apicales, citoplasma eosinófilo, núcleos alargados y basales, muchas<br />
mitocondrias y chapa estriada. Presentan muchos gránulos de secreción con glicoproteínas vertidas por<br />
exocitosis formando la capa protectora de la mucosa.<br />
Estas células principales son capaces de absorber agua y electrolitos. Estos elementos se excretan por la<br />
membrana basolateral, de manera que se genera una alta presión hidrostática y el agua fluye hacia la lámina<br />
propia, llegando a la sangre.<br />
Funcionan pues de manera similar a los enterocitos del colon.<br />
La lámina propia carece de vasos linfáticos, pero es muy rica en capilares y vénulas pequeñas fenestradas.<br />
Submucosa<br />
Es similar a la del colon. Encontramos glándulas específicas, tubuloalveolares, de tipo mucoso, sobre todo en<br />
cuello.<br />
Capa Muscular<br />
Está muy desarrollada. Encontramos haces de miocitos en todas direcciones, con un endomisio de fibras<br />
elásticas y colágenas. La disposición helicoidal de las fibras musculares permite una contracción máxima de la<br />
pared que estimula la salida masiva de la bilis. Esta salida está controlada por CCK y ACh.<br />
Adventicia<br />
Se encuentra asociada a peritoneo, formando una serosa. Es muy gruesa, siendo un conjuntivo denso con fibras<br />
de todo tipo y adipocitos. Se encuentra en continuidad con el conjuntivo asociado al hígado. En esta adventicia<br />
hay vasos de gran calibre, sanguíneos y linfáticos. Encontramos un plexo que recibe la linfa hepática.<br />
Encontramos ganglios del parasimpático y, a veces, los conductos de Luschka: vestigios de conductos biliares<br />
del desarrollo embrionario pero que no culminaron su desarrollo.
Página69<br />
PÁNCREAS EXOCRINO<br />
El páncreas es una glándula aneja situada en la cavidad abdominal a nivel de L2-L3, y se divide en cabeza,<br />
cuerpo y cola. La cabeza se adhiere al duodeno y el resto del páncreas, al bazo.<br />
Es una glándula mixta, con un <strong>com</strong>ponente exocrino y uno endocrino. Su función exocrina está relacionada con<br />
enzimas digestivas que forman el jugo pancreático, mientras que la función endocrina tiene que ver con la<br />
síntesis de hormonas relativas al control de la glucemia.<br />
Embriología<br />
Evolutivamente, el páncreas solo aparece desde aves y mamíferos. En los vertebrados inferiores no existe un<br />
órgano unificado que cumpla todas las funciones del páncreas, si bien estas funciones sí que se realizan de<br />
manera separada.<br />
El páncreas procede de dos divertículos que se forman en el epitelio anterior, en la zona del duodeno. Cada<br />
divertículo es independiente en irrigación e inervación y posteriormente se fusionan. En la patología conocida<br />
<strong>com</strong>o páncreas dividido, estos dos divertículos no se fusionan, de modo que el drenaje se produce<br />
mayoritariamente por el conducto de Santorini en vez de por el de Wirsung, <strong>com</strong>o es habitual.<br />
Parte de las células epiteliales que forman el páncreas se agregan formando acinos, que en el centro dejan una<br />
pequeña luz y más se forman las vías conductoras.<br />
En la porción endocrina, las células epiteliales no forman acinos, sino acúmulos <strong>com</strong>pactos, que quedan<br />
dispersos entre el parénquima exocrino.<br />
Estroma<br />
El estroma del páncreas es muy reducido y no presenta cápsula, sino solo una envuelta fina y laxa asociada al<br />
peritoneo.<br />
En la parte interna, encontramos un escaso estroma que permite una lobulación in<strong>com</strong>pleta del páncreas.<br />
Encontramos aquí abundantes corpúsculos de Pacini.<br />
Parénquima Exocrino: Adenómeros Exocrinos<br />
Son ácinos serosos, con células altas piramidales en torno a una luz estrecha y una ancha superficie basal.<br />
Nunca encontramos células mioepiteliales.<br />
Sus células secretoras son típicas serosas: altas, con uniones estrechas y núcleo plano. Presentan gránulos de<br />
zimógeno muy eosinófilos, si bien el resto de la célula es basófila, sobre todo la base, debido a un alto<br />
desarrollo del RER.<br />
La secreción que forman es rica en proteínas y constituye el jugo pancreático. Este jugo se segrega en cantidad<br />
de 1L diario, igual que el hígado con la bilis. Se <strong>com</strong>pone de enzimas que permiten la digestión última en el<br />
duodeno: azucares, lípidos, etc. La concentración de cada enzima depende de la persona y la dieta.<br />
Las enzimas pancreáticas más importantes son tripsinógeno, quimotripsinógeno, procarboxipeptidasa,<br />
proaminopeptidasa, elastasa, α-amilasa, lipasas, colesterolasa, fosfolipasa, desoxirribonucleasa y ribonucleasa.<br />
Buena parte de las enzimas son formas inactivas. Estas formas inactivas se activan en el duodeno, para lo que<br />
son fundamentales las enteroquinasas situadas en la chapa estriada de los enterocitos.<br />
El jugo pancreático es drenado por un sistema de vías que <strong>com</strong>ienza con las células centroacinares. Estas<br />
células se encuentran dentro del ácino, tapizando la luz por encima de las células secretoras. Son aplanadas, no<br />
piramidales, muy pálidas, carecen de gránulos de zimógeno y tienen desarrollo de orgánulos escaso. Están<br />
unidas entere sí y a las células secretoras. Las células centroacinares son la primera vía excretora y además<br />
segregan una proteína inhibidora de la tripsina. Esta inhibición es importante porque una activación prematura<br />
de las enzimas pancreáticas destruiría la glándula.
Página70<br />
El resto de vías que permiten la secreción son: el conducto intercalar, el intralobulillar, y dos principales. No<br />
encontramos conducto estriado.<br />
Las células intercalares, además de la proteína inhibidora de tripsina, contribuyen segregando sodio y<br />
bicarbonato. Esto es importante para neutralizar la acidez del quimo, lo que es imprescindible para la<br />
activación de las enzimas pancreáticas.<br />
El conducto intercalar está tapizado por un epitelio cúbico simple.<br />
El conducto interlobulillar está tapizado por en epitelio prismático simple, y en él encontramos células<br />
endocrinas y caliciformes<br />
Los conductos principales están tapizados por epitelio prismático simple. Encontramos asociado a este epitelio<br />
un conjuntivo (lámina propia).<br />
El conducto principal mayor o de Wirsung recoge el jugo pancreático que se segrega a la cola y va avanzando<br />
hasta llegar a la cabeza recogiendo todo el jugo, por lo que tiene forma de raspa de pescado. Se une al<br />
colédoco y alcanza el duodeno por la ampolla de Vater, donde encontramos el esfínter de Oddi.<br />
El conducto accesorio o de Santorini drena tan solo el jugo pancreático que se produce en la cabeza.<br />
Desemboca directamente en duodeno por la carúncula (abertura) menor.
Página71<br />
SISTEMA LIFÁTICO<br />
El sistema linfático es el sistema encargado de la inmunidad del organismo, llevando a cabo respuestas<br />
inmunológicas de defensa. Está formado por linfocitos y sus células madre, macrófagos, células dendríticas,<br />
granulocitos; así <strong>com</strong>o por <strong>com</strong>ponentes solubles: factores del <strong>com</strong>plemento, linfocinas, quimiocinas, y moléculas<br />
antibacterianas (defensinas, lisozimas, lactoferrina, catelicina).<br />
En el organismo hay una primera línea de defensa contra macromoléculas extrañas, que es el epitelio. Pero es el<br />
tejido linfoide el que <strong>com</strong>pone la segunda línea de defensa protegiendo al organismo de las moléculas extrañas que<br />
hayan atravesado la barrera epitelial o a aquellas propias que se hayan transformado. El tejido linfoide actúa<br />
mediante células inmuno<strong>com</strong>petentes, es decir, células capaces de distinguir entre moléculas propias y las no<br />
propias; ante estas las células linfáticas actúan atacándolas mediante el proceso de la inmunidad.<br />
Las células del sistema inmunitario se encuentran situadas al lado de otras de modo que forman una malla o retículo,<br />
dejando huecos donde se ubican las células inmunitarias. Esta malla se denomina estroma, y suele estar formado por<br />
células reticulares y fibras reticulares. En el timo el estroma no es reticular, sino de naturaleza epitelial.<br />
El tejido linfoide es una variación del tejido conjuntivo reticular, cuya célula principal es el linfocito. Puede ser difuso<br />
o nodular si los linfocitos están dispersos de forma laxa en el tejido conjuntivo o folicular si los linfocitos están<br />
asociados formando folículos (cúmulos densos esféricos).<br />
Además de tejido linfoide, el sistema inmunitario consta también de órganos linfoides. Estos órganos pueden ser<br />
primarios, si están implicados en maduración y formación de linfocitos inmuno<strong>com</strong>petentes (médula ósea y timo); o<br />
secundarios, si en ellos ocurren reacciones inmunológicas (ganglios linfáticos, bazo, y MALT).<br />
Los linfocitos circulan por la sangre y la linfa o bien están situados intraepitelialmente.<br />
CÉLULAS LINFOIDES<br />
Los linfocitos son leucocitos (entre el 20-50% del total) de entre los que suelen describir 3 grupos de linfocitos de<br />
acuerdo al tamaño, es decir, pequeños, medianos y grandes con diámetros aproximados de 6, 12 y 18μm<br />
respectivamente, aunque después de la transformación linfática estos linfocitos pueden llegar hasta los 30μm de<br />
diámetro. La mayoría de los linfocitos circulantes en sangre y linfa son células inmuno<strong>com</strong>petentes recirculantes, es<br />
decir, son capaces de reconocer y responder ante antígenos extraños y cambiar de posición anatómica.<br />
Las células precursoras de las células linfoides son las Unidades Formadoras de Colonias (UFC) y dan origen a los<br />
linfocitos madre o stem que se diferencian a linfocitos B o T en las primeras etapas de la vida fetal. A lo largo del<br />
desarrollo uterino y en la vida postnatal la posición de maduración de las células linfoides va cambiando hasta que<br />
finalmente se sitúan en los órganos linfoides primarios y secundarios.<br />
Ante un antígeno, los linfocitos pueden responder con una respuesta mediada por células o a una respuesta humoral<br />
en la que intervienen anticuerpos (Ac), también llamados inmunoglobulinas o Ig.<br />
Linfocitos B<br />
Son el 10-20% de los linfocitos y son células programadas genéticamente para responder únicamente ante un tipo de<br />
antígeno o sitio antigénico, de modo que al contactar con este se diferencian en inmunoblastos o plasmoblastos que<br />
proliferan y finalmente se diferencian a:<br />
<br />
<br />
Plasmocitos: sintetizan y secretan un Ac específico.<br />
Células con memoria: permiten que el organismo responda con mayor rapidez si vuelve a penetrar el mismo<br />
antígeno. Así, intervienen en la respuesta secundaria.<br />
Cuando el plasmocitos secreta el Ac, este se fija al antígeno y forma un <strong>com</strong>plejo antígeno-Ac que se eliminan<br />
principalmente por fagocitosis de macrófagos y eosinófilos. Estos <strong>com</strong>plejos también activan el sistema del
Página72<br />
<strong>com</strong>plemento, un conjunto de P plasmáticas, entre los que se encuentra el <strong>com</strong>ponente C3 que se fija a una bacteria<br />
antigénica y actúa <strong>com</strong>o ligando para que esta pueda ser eliminada por macrófagos. En algunas ocasiones este<br />
<strong>com</strong>plejo produce la lisis de la célula que contiene el antígeno.<br />
En definitiva, los linfocitos B <strong>com</strong>ponen la inmunidad humoral mediante la secreción de Ig.<br />
INMUNOGLOBULINAS o ANTICUERPOS (Ig/Ac)<br />
Las inmunoglobulinas son estructuras en forma de Y que fija antígenos. Están formadas por dos cadenas pesadas y<br />
dos cadenas ligeras que se asocian mediante enlaces del tipo puente disulfuro. Cada una de las cadenas tiene una<br />
secuencia aminoacídica constante y otra variable (C y V, respectivamente). Además, en cada inmunoglobulina se<br />
puede distinguir una región Fab, que incluye a la zona donde se une al antígeno, y una zona Fc formada por las<br />
regiones C de las cadenas pesadas que permite la unión con el sistema del <strong>com</strong>plemento.<br />
Existen varios tipos de Ig con funciones diferentes; destacan las Ig M, A, D, G y E.<br />
Linfocitos T<br />
Los linfocitos T representan el 35% de los linfocitos y tras interaccionar con su antígeno se transforman en<br />
linfoblastos, que proliferan y se transforman a distintos tipos de linfocitos T efectores entre los que se encuentran:<br />
Linfocitos Citotóxicos, Células T killer o LCT: reconocen en células vecinas péptidos que evidencian infección<br />
viral o de tumoración y las eliminan mediante lisis.<br />
Linfocitos T Helper o T H : asisten a los linfocitos B y otras células en su respuesta ante un antígeno. Estas<br />
células tienen en su superficie receptores CD3 por los que se unen a un <strong>com</strong>plejo peptídico de superficie de<br />
macrófagos o células B que hayan procesado un antígeno. Esto estimula a los linfocitos T helper, por lo que<br />
se dividen y producen unas hormonas polipeptídicas denominadas interleucinas o Ile que estimulan a los<br />
linfocitos B a dividirse y producir Ac.<br />
Linfocitos T supresores o células T S : suprimen la actividad de los linfocitos B.<br />
Según el tipo de linfocito aparecen en su superficie una serie de Cúmulos de Proteínas o CD que permiten identificar<br />
el tipo de linfocito con el que nos encontramos. Así, existen:<br />
CD2 Células NK CD20 Lf B<br />
CD3 Linfocitos T CD28 Lf T H<br />
CD4 Lf T H CD40 Lf B<br />
CD8 LCT y Lf T S CD56 Células NK<br />
CD16<br />
Células NK
Página73<br />
Los CD2 también se conocen <strong>com</strong>o receptores para eritrocitos de carnero, porque forman rosetas sobre los<br />
eritrocitos de los carneros diferenciándolos de los linfocitos B.<br />
Los CD5 y CD7 aparecen en los linfocitos T y reconocen Ac, sobre todo IgM.<br />
Los CD3 reconocen señales antigénicas dadas por células presentadoras de antígenos.<br />
Además, los linfocitos T sensibilizados por el contacto de un antígeno específico, liberan las linfocinas, unas<br />
sustancias solubles que estimulan la actividad de monocitos y macrófagos.<br />
Linfocitos Grandes<br />
Fotomicrografía de un Linfocito T y de un Linfocito B<br />
Los linfocitos grandes o células nulas representan el 5-10% de los linfocitos de la sangre periférica y contienen<br />
grandes gránulos azurófilos; no son adherentes ni son fagocíticos. Existen dos subclases de linfocitos grandes:<br />
Los Linfocitos Natural Killer o NK tienen una actividad que no depende de la activación antigénica. Se activan con la<br />
Ile-2 para eliminar tipos celulares malignos en procesos no mediados por Ac.<br />
Los Linfocitos Grandes que median la Citotoxicidad Celular Dependiente de Anticuerpos o CCDA intervienen en la<br />
unión de una célula efectora que tenga la porción Fc del Ac con el Ac correspondiente, ya sea libre o presente en una<br />
célula que esté previamente opsonizada (tenga sistema de <strong>com</strong>plemento en su superficie). Así, esta célula permite<br />
que un antígeno o una célula que presente este antígeno y haya sido reconocida, sea eliminada por lisis mediante<br />
una célula efectora.<br />
Células Presentadoras de Antígenos<br />
Dentro de este grupo se encuentran células <strong>com</strong>o los linfocitos B, macrófagos tisulares, células de Kupffer, células de<br />
Langerhans epidérmicas o células reticulares dendríticas de los nódulos linfáticos. Estas células fagocitan un<br />
elemento extraño, lo degradan a péptidos y fijan los restos a moléculas de mayor histo<strong>com</strong>patibilidad o CMH de su<br />
membrana plasmática.<br />
Los CMH son productos genéticos de un gran gen localizado en el cromosoma 6 que permiten la identificación de las<br />
células propias y también para activar células del sistema inmune para atacar la célula si el CMH no es reconocido.
Página74<br />
VASOS LINFÁTICOS<br />
Los vasos linfáticos se inician <strong>com</strong>o redes de capilares ciegos en el tejido conectivo laxo que abundan bajo el epitelio<br />
y las membranas mucosas. Permiten la extracción de sustancias y líquidos del LEC produciendo la linfa; este paso de<br />
moléculas esta facilitado por la mayor permeabilidad de los vasos linfáticos, en <strong>com</strong>paración con los sanguíneos,<br />
permitiendo el paso de moléculas grandes.<br />
La linfa originada en los vasos sanguíneos atraviesa los ganglios linfáticos, donde las sustancias extrañas se<br />
concentran por el sistema reticular formado por las células dendríticas y se da la respuesta inmune. Por último, los<br />
vasos linfáticos drenan la linfa al sistema venoso.<br />
TEJIDO LINFOIDE DIFUSO, NÓDULOS LINFÁTICOS Y AMÍGDALAS<br />
Tejido Linfoide Difuso<br />
El tejido linfoide difuso está formado por acumulaciones de tejido linfoide no encapsulado presente en las vías<br />
aéreas, digestivas y genitourinarias. Se localizan en la lámina propia de estas vías formando el Tejido Linfoide<br />
Asociado a Mucosas o MALT, que según su localización recibe un nombre u otro. Estos tejidos contienen linfocitos<br />
que interactúan con el medio y al contactar con antígenos viajan a ganglios linfáticos cercanos para sufrir procesos<br />
de diferenciación y proliferización.<br />
El epitelio en estas zonas de MALT es diferente al del resto del órgano y se denomina epitelio asociado a folículos<br />
(FAE). En el MALT encontramos un elevado número de células plasmáticas que secretan mayoritariamente IgA,<br />
aunque también pueden secretar IgG, IgM e IgE.<br />
En el MALT no encontramos vías linfáticas aferentes. El epitelio tiene uniones oclusivas que impiden que los<br />
antígenos lo atraviesen.<br />
GALT<br />
Se denomina GALT al tejido linfoide asociado al tubo digestivo, <strong>com</strong>puesto por varias unidades, llamada cada una<br />
GALU.<br />
Unas estructuras específicas más desarrolladas de este tejido linfoide en el tubo digestivo son las placas de Peyer,<br />
que son tan extensas y tan gruesas que son visibles a simple vista. Pueden ocupar no solo la zona de la mucosa sino<br />
que también invaden la submucosa. Se observan sobre todo en el íleon terminal.
Página75<br />
La placa de Peyer se forma cuando en una zona concreta del tubo digestivo se desarrolla un número muy elevado de<br />
folículos de tejido parafolicular. Aunque pueden aparecer en cualquier parte, se suelen formar en la pared opuesta a<br />
donde se inserta el mesenterio. El número máximo de placas de Peyer es de más de 300 y se alcanza en la infancia.<br />
La estructura histológica general del tubo digestivo está alterada en las placas de Peyer: apenas se encuentran<br />
criptas de Lieberkuhn y se disgrega la muscularis mucosae, porque los folículos son tan grandes que no caben en la<br />
mucosa e invaden la submucosa. Por su parte, el epitelio intestinal que cubre las placas apenas contiene células<br />
caliciformes. Los enterocitos dejan de ser células absortivas y adquieren una alta capacidad para degradar antígenos.<br />
La región cupular de la placa de Peyer se <strong>com</strong>pone de la cúpula y el epitelio cupular. La cúpula es una banda de<br />
lámina propia con abundancia de células inmunitarias, sobre todo linfocitos B, aunque también hay macrófagos,<br />
células dendríticas. La cúpula rodea o abraza el centro germinal.<br />
El centro germinal se <strong>com</strong>pone de linfocitos B, T CD4+ y células presentadoras de antígenos. Los linfocitos B acuden<br />
atraídos por las células dendríticas y se transforman en células plasmáticas. Éstas, en su inmensa mayoría, migran<br />
hacia otros folículos próximos de la placa o a zonas mas alejadas de la mucosa. La mayoría de las células plasmáticas<br />
secretan IgA, aunque también se encuentra IgM e IgE, que se libera para atraer mastocitos que liberan histamina. El<br />
enterocito capta la IgA después de que se dimeriza por acción de la cadena J. Este <strong>com</strong>plejo se une a un receptor en<br />
la membrana basal del enterocito. Se libera entonces la IgA unida al <strong>com</strong>ponente secretor (IgA secretor). IgA se une<br />
al <strong>com</strong>ponente secretor para dificultar su degradación y la unión de los microorganismos a los enterocitos.<br />
Durante una enteritis las placas de Peyer crecen tanto en longitud <strong>com</strong>o en grosor.<br />
BALT<br />
BALT es el tejido linfoide asociado a las vías aéreas; la unidad que lo <strong>com</strong>pone es el BALU. El BALT se encuentra sobre<br />
todo en la tráquea. Hay menos tejido linfoide que en el tubo digestivo pero los linfocitos dispersos son más <strong>com</strong>unes<br />
que en aquél.<br />
SALT<br />
El SALT es el tejido linfoide asociado a la piel. La propia piel forma parte del sistema inmunitario porque es una<br />
barrera, es decir, que forma parte de la inmunidad innata. El SALT refuerza la inmunidad a través de la piel porque<br />
puede detectar antígenos.<br />
Se <strong>com</strong>pone de linfocitos en la epidermis, células de Langerhans, macrófagos de la dermis y queratinocitos.<br />
Nódulos Linfáticos<br />
Son concentraciones localizadas de linfocitos en las paredes del tubo digestivo, vías aéreas y genitourinarias que<br />
están nítidamente definidos pero carecen de una cápsula. Aparecen en zonas específicas <strong>com</strong>o el apéndice y el ciego,<br />
las amígdalas y las placas de Peyer.<br />
Un nódulo linfático <strong>com</strong>puesto en su mayor parte por linfocitos pequeños se denomina nódulo primario, pero si<br />
aparecen linfocitos grandes será un nódulo secundario, que presenta dos zonas:<br />
Centro Germinativo: región central de carácter reactivo que se desarrolla cuando un linfocito ha reconocido<br />
a su antígeno y sufre transformación blástica. Aparece menos teñido porque contiene linfocitos grandes del<br />
tipo linfoblastos y plasmoblastos que por la gran cantidad de eucromatina de su núcleo hacen esta zona más<br />
clara.<br />
Anillo externo de linfocitos pequeños; aparece más teñido porque los linfocitos pequeños presentan<br />
heterocromatina.<br />
La presencia de un nódulo secundario evidencia que en ese tejido se están llevando a cabo procesos de<br />
transformación blástica y en consecuencia de inmunidad.
Página76<br />
Amígdalas<br />
Las amígdalas son acumulaciones de nódulos linfáticos que se sitúan alrededor del istmo de las fauces, la orofaringe<br />
y la rinofaringe formando el anillo Waldeyer en la bifurcación de los aparatos respiratorio y digestivo. Son 4: lingual,<br />
palatina, tubárica y faríngea.<br />
Las amígdalas tienen una superficie irregular, con invaginaciones bastante profundas denominadas criptas. En las<br />
criptas, aunque hay epitelio, éste es difícil de ver debido a la gran cantidad de linfocitos. Suelen estar además<br />
taponadas por la presencia de células descamadas tanto epiteliales <strong>com</strong>o linfoides.<br />
Las amígdalas tienen una cápsula de tejido conjuntivo que la delimita y facilita su extirpación. Presenta folículos de<br />
linfocitos B rodeados de tejido parafolicular de linfocitos T.<br />
Las amígdalas presentan además vasos eferentes que drenan la linfa hacia ganglios linfáticos, así <strong>com</strong>o vénulas de<br />
endotelio alto (VEA).<br />
Amígdala Lingual<br />
La amígdala lingual se encuentra en la mucosa de la raíz de la lengua.<br />
Está tapizada por epitelio plano estratificado no queratinizado con abundantes linfocitos. Este epitelio se invagina<br />
formando criptas no especialmente profundas ni ramificadas en cuy fondo desembocan las glándulas salivales<br />
mucosas. Alrededor de la mucosa lingual encontramos el músculo lingual.<br />
Cada cripta se asocia con una serie de folículos y tejido linfoide difuso, ambos en conjunto denominados folículo<br />
lingual, siendo la amígdala el conjunto de todos los folículos linguales presentes en la mucosa lingual. Entre folículo y<br />
folículo queda suficiente espacio para observar bandas de tejido conjuntivo en conexión con la cápsula.<br />
Amígdalas Palatinas<br />
Las amígdalas palatinas son dos, situadas a ambos lados de entrada de la faringe. Están tapizadas por epitelio plano<br />
estratificado no queratinizado muy grueso (hasta 1 cm)<br />
Las criptas son mucho más profundas y ramificadas, muy próximas entre si. Tanto en las criptas <strong>com</strong>o en el epitelio<br />
encontramos una gran cantidad de linfocitos.<br />
La cápsula de la amígdala lingual es muy gruesa y está muy tabicada, por lo que las amígdalas llegan a dividirse en<br />
lóbulos y lobulillos. Al igual que en la amígdala lingual, en la amígdala palatina encontramos glándulas salivales que<br />
en este caso no desembocan en las criptas sino en la superficie. Alrededor de la amígdala palatina encontramos<br />
músculo faríngeo.<br />
Amígdala Faríngea<br />
La amígdala faríngea es un órgano impar y se sitúa en la mucosa de la rinofaringe, adherida al periostio. Está tapizada<br />
por un epitelio respiratorio (epitelio pseudoestartificado ciliado con células caliciformes). Presenta parches de<br />
epitelio malpigiano. Esta amígdala no tiene criptas, pero sí pliegues. La cápsula tiene un <strong>com</strong>ponente mayoritario de<br />
fibras elásticas.<br />
La amígdala faríngea tiene glándulas seromucosas que drenan en la superficie. En ocasiones esta amígdala se inflama<br />
obstruyendo paso del aire y dando lugar a la patología <strong>com</strong>únmente conocida <strong>com</strong>o vegetaciones.
Página77<br />
GANGLIOS LINFÁTICOS<br />
Los ganglios linfáticos son estructuras intercaladas en la trayectoria de los vasos linfáticos.<br />
Existe un número muy elevado de ganglios, entre 600 y 700, que suelen aparecer en cadenas o grupos y son más<br />
abundantes en regiones <strong>com</strong>o axilas, cuello, ingles, región paraaórtica, mesenterios y mediastino.<br />
Los ganglios son órganos aplanados con forma de haba con un tamaño de unos 0,2 cm, aunque su estructura varía<br />
mucho en función de la situación funcional. Constan de una cápsula, corteza y médula.<br />
Los ganglios linfáticos filtran la linfa que fluye a su través y posibilitan que tengan lugar reacciones ante cualquier<br />
antígeno que esté circulando por las inmediaciones del ganglio.<br />
Vascularización<br />
La vascularización sanguínea de los ganglios corre a cargo de arterias que entran por el hilio ganglionar y dan<br />
arteriolas que se ubican en las trabéculas de la cápsula, desde donde acceden a la médula del ganglio formando<br />
capilares y a la corteza formando redes capilares que retornan por vénulas que aparecen en el fondo de la corteza.<br />
En los ganglios linfáticos encontramos además vénulas de endotelio alto (VEA), que son los principales puntos de<br />
acceso para los linfocitos T y B circulantes. Las VEA presentan un endotelio con un glicocálix con abundantes<br />
moléculas de adhesión <strong>com</strong>o selectina P, ICAM-1, ICAM-2, VCAM o ELAM-1. Estas moléculas sirven para<br />
interaccionar con receptores de la membrana de los linfocitos <strong>com</strong>o selectina L, LFA-1 y receptores acoplados a<br />
proteína G.<br />
Gracias a estas moléculas de adhesión y sus receptores disminuye la velocidad de circulación de los linfocitos, que<br />
<strong>com</strong>ienzan a adherirse al endotelio hasta que queda pegado a él. Estos vasos permiten el paso de los linfocitos que<br />
se adhieren a su pared pero impiden el paso de líquido al parénquima.<br />
Cada ganglio recibe numerosas vías linfáticas aferentes que perforan la cápsula y conforman el seno linfático<br />
subcapsular. La linfa es drenada, después de pasar por el ganglio, por uno o varios vasos eferentes que salen por el<br />
hilio: los vasos eferentes. Dentro del ganglio, la linfa circula por los senos linfáticos. La linfa sigue un flujo<br />
unidireccional, con valvas que impiden el reflujo, es decir, el retroceso de la linfa al ganglio anterior. El flujo linfático<br />
tiene una regulación intrínseca debido a los miocitos lisos de los vasos linfáticos y una regulación extrínseca debido al<br />
movimiento de los tejidos circundantes.
Página78<br />
Senos<br />
La linfa se aloja en el seno subcapsular o marginal y desde aquí se distribuye por los senos paratrabeculares o<br />
intermedios, en la corteza, o bien por los senos medulares, en la médula, desde donde sale hacía las vías eferentes<br />
del ganglio.<br />
Los senos linfáticos son espacios irregulares limitados por endotelio típico en la pared que limita con la cápsula, pero<br />
en la zona opuesta el endotelio es mucho más laxo y la lámina basal del mismo es in<strong>com</strong>pleta, lo que permite que<br />
por esa zona entren macrófagos al seno, así <strong>com</strong>o prolongaciones de células reticulares y fibras reticulares.<br />
Estroma<br />
Cápsula<br />
La cápsula del ganglio está formada por estratos de tejido conjuntivo denso, con fibras colágenas de tipo I y elásticas.<br />
La cápsula es más gruesa a nivel del hilio y desde la cápsula surgen trabéculas que se ramifican hacia el interior del<br />
ganglio.<br />
Estroma<br />
En el interior del ganglio encontramos un estroma de células reticulares y fibras colágenas de tipo III. El estroma se<br />
tiñe con técnicas de impregnación argéntica. Los huecos que deja la malla de estroma sirve para que se alojen las<br />
células libres, principalmente linfocitos, si bien también se encuentran macrófagos y células dendríticas<br />
interdigitantes y foliculares.<br />
Los macrófagos aparecen por todo el ganglio, son células presentadoras de antígeno y expresan MHC-II. Fagocitan<br />
además linfocitos apoptóticos.<br />
CÉLULAS DENDRÍTICAS<br />
Las células dendríticas interdigitantes son células estrelladas con un núcleo oval heterocromático que proceden de<br />
otras localizaciones, llegando al ganglio cuando en éste se detecta algún antígeno. Pueden reubicarse en el ganglio<br />
de modo que interaccionan con los linfocitos de memoria.<br />
Las células dendríticas foliculares están en los folículos del ganglio y no expresan MHC-II, por lo que no son células<br />
presentadoras de antígenos una función que suele estar típicamente asociada a las células dendríticas. No obstante,<br />
las células dendríticas foliculares sí que tienen otros marcadores, <strong>com</strong>o receptores del <strong>com</strong>plemento. Estas células<br />
con capaces de mostrar antígenos intactos en su membrana durante años. Atraen hacia los folículos, donde residen,<br />
a los linfocitos que se hayan activado en paracorteza. Evitan asimismo que los linfocitos B entren en apoptosis.<br />
Corteza<br />
La corteza es más basófila que la médula. Se puede dividir en una corteza externa, que tiene folículos y tejido linfoide<br />
interfolicular, y una corteza más profunda o paracorteza que nunca muestra folículos. La corteza y la médula no<br />
tienen un límite definido en el ganglio.<br />
La corteza externa se caracteriza por la aparición de folículos: agrupaciones esféricas de linfocitos T muy apretados y<br />
células dendríticas foliculares. Los folículos primarios son aquellos homogéneos, con linfocitos B aún no<br />
<strong>com</strong>prometidos. Los folículos secundarios aparecen después de que se produzca una reacción inmunológica. En<br />
estos folículos secundarios de distinguen zonas más teñidas y otras menos teñidas. La zona menos teñida<br />
corresponde al centro germinal, mientras que la zona más teñida corresponde a la corona o manto.<br />
El centro germinal es el lugar donde se dan procesos de diferenciación, proliferación y destrucción de linfocitos B. El<br />
linfocito B llega al folículo secundario, prolifera y se diferencia.<br />
Encontramos, junto con los linfocitos B, células dendríticas interdigitantes, linfocitos T H y macrófagos. En el centro<br />
germinal hay una zona oscura con centroblastos y otra clara con centrocitos.<br />
La corona o manto es una zona periférica que rodea al centro germinal. Es más gruesa en la zona próxima a la<br />
cápsula. En la corona encontramos linfocitos pequeños con IgM e IgD. La zona del manto cercana al folículo es la<br />
zona marginal, donde se ubican los linfocitos B de memoria.
Página79<br />
La paracorteza, corteza profunda o corteza yuxtamedular no presenta folículos. Tiene linfocitos T y células<br />
dendríticas interdigitantes. Es en esta zona donde encontramos las vénulas de endotelio alto por donde entran los<br />
linfocitos. Después de extravasarse, los linfocitos T quedan en paracorteza y los linfocitos B migran hacia la corteza<br />
externa. Si los linfocitos no experimentan una reacción inmunológica a las pocas horas, salen del ganglio.<br />
Médula<br />
La médula tiene amplios senos medulares. Los linfocitos se organizan aquí en cordones laxos ramificados.<br />
Encontramos, además de linfocitos, células plasmáticas y macrófagos. En los ganglios mesentéricos encontramos<br />
también mastocitos. Durante una reacción inmunológica, se produce una síntesis masiva de anticuerpos en<br />
reacciones inmunitarias.<br />
Funciones<br />
Los ganglios linfáticos filtran la linfa eliminando patógenos. Ralentizan la velocidad de la linfa, lo que permite la<br />
detección de anticuerpos. Esta función es tan importante que de hecho la linfa pasa al menos por dos ganglios antes<br />
de verter a la sangre. Los ganglios también filtran la linfa para facilitar la respuesta inmunológica: el antígeno llega al<br />
ganglio libremente o unido a células presentadoras de antígenos y activa los linfocitos del ganglio que sean<br />
específicos contra ese antígeno, lo que desencadena respuestas inmunológicas celulares y humorales, por lo que hay<br />
células y moléculas que salen del ganglio hacia la zona por la que entró el antígeno. Las células de memoria también<br />
salen del ganglio hacia la médula ósea, el bazo y el MALT.<br />
Reacciones Inmunológicas<br />
El antígeno llega, libre o asociado a una célula presentadora de antígeno, al ganglio linfático.<br />
Se activan entonces los linfocitos Th específicos para ese antígeno, aun no <strong>com</strong>prometidos.<br />
Este proceso de activación de linfocitos Th ocurre en paracorteza. Cuando se activan, proliferan y aumentan su<br />
tamaño, y al cabo de 5 o 6 días son linfocitos Th efectores o bien de memoria. Se activa además la población de<br />
linfocitos Tc, que también proliferan y crecen, diferenciando a Tc efectores o de memoria. Los linfocitos ya activados<br />
salen del ganglio hacia la zona por donde entró el antígeno.<br />
Además, los linfocitos Th activados desencadenan la activación de linfocitos B en paracorteza. Los linfocitos B<br />
acceden a paracorteza por las vénulas de endotelio alto o desde los folículos de la corteza externa. Los linfocitos B se<br />
activan en paracorteza al contactar con los linfocitos Th activados. En consecuencia, a los 5 días de la llegada del<br />
antígeno en paracorteza se generan pequeños focos donde se secretan las primeras inmunoglobulinas, de tipo IgM e<br />
IgG.<br />
Estas inmunoglobulinas secretadas salen por los senos linfáticos. En días posteriores, los linfocitos B activados migran<br />
a la médula ósea, donde la secreción de inmunoglobulinas se realiza directamente a la sangre.<br />
A la semana, migran linfocitos B y Th a los folículos primarios de la corteza externa, ya que las células dendríticas<br />
foliculares de dichos folículos los atraen. Los linfocitos B y Th quedan en la zona central del folículo proliferando, por<br />
lo que pasan a denominarse centroblastos. En ese momento se reduce la IgD expresada. El folículo pasa a llamarse<br />
entonces folículo secundario. Los linfocitos que ya estaban previamente en el folículo son empujados hacia el borde<br />
formando la corona o manto. Estos linfocitos de la corona son linfocitos pequeños con IgM e IgD. El manto es más<br />
grueso en la zona más próxima a la cápsula.<br />
El centro germinal es el lugar del folículo donde están los linfocitos activados o centroblastos, que se dividen,<br />
diferencian y se destruyen. Además encontramos células dendríticas foliculares, estroma de células reticulares,<br />
macrófagos y linfocitos T CD4+. Los centroblastos no ocupan todo el centro germinal por igual, sino que tienden a<br />
situarse en la mitad, más oscura, zona donde sufren las hipermutaciones somáticas, proceso por el que la célula<br />
modifica el orden de sus genes de modo que cambia la afinidad del receptor del linfocito por el antígeno. De este<br />
proceso surgen los centrocitos, que se sitúan en la zona más externas del centro germinal. Los centrocitos con<br />
receptores de baja afinidad por su antígeno entran en apoptosis y son eliminados por los macrófagos. Los centrocitos<br />
con receptores de alta afinidad por su antígeno se activan solo si contactan con células dendríticas foliculares. Los<br />
centrocitos que sobreviven a este proceso se dividen y originan linfocitos B de memoria y plasmoblastos que migran<br />
a la médula ósea, donde se habla ya de células plasmáticas, que vierten sus inmunoglobulinas, de tipo IgM e IgG, a la<br />
sangre. El tipo de inmunoglobulina depende del tipo de tejido infectado. Las células de memoria salen del centro<br />
germinal y se quedan bien en el manto, bien recirculando por el organismo.
Página80<br />
En una respuesta inmunológica suben los niveles de linfocitos T e inmunoglobulinas en linfa y sangre. La recirculación<br />
de los linfocitos T provoca que cada vez haya más linfocitos T pasando por el ganglio y aumentando así la respuesta<br />
inmunológica.<br />
Los ganglios linfáticos aumentan de tamaño durante una infección (linfadenitis). En estas situaciones también se<br />
detectan neutrófilos. Los ganglios retienen a menudo células cancerosas, por lo que es frecuente su extirpación para<br />
prevenir ciertos tipos de tumor. En el ganglio tienen lugar además linfomas malignos, cuyo foco son principalmente<br />
sobre los linfocitos<br />
TIMO<br />
El timo es un órgano bilobulado situado en el mediastino superior, bajo el esternón. Pesa un máximo de 50 g. en la<br />
infancia. Es un órgano linfoide primario, responsable de la diferenciación y maduración de los linfocitos T, de modo<br />
que cuando salen del timo ya son inmuno<strong>com</strong>petentes aunque no estén aun <strong>com</strong>prometidos.<br />
La manipulación de ratones recién nacidos mediante extirpación de timo provoca un descenso brusco de linfocitos T,<br />
es decir, los ratones no pueden mostrar reacciones inmunitarias celulares. Si un adulto no tiene timo, no se produce<br />
un grave descenso de los niveles de linfocitos T, porque en la infancia se crea un pool de linfocitos T en el timo, pool<br />
que ya se mantiene constante a pesar de que el timo funciona durante el resto de la vida.<br />
El timo es un órgano cuyo peso máximo se alcanza en la infancia. El timo sigue siendo funcional en la edad adulta,<br />
pero la actividad más esencial sucede durante la infancia y con la pubertad hay un descenso brusco del peso del<br />
timo. Este descenso puede tener que ver con el desarrollo de hormonas sexuales. Al involucionar, el timo adquiere<br />
peso en tejido adiposo.<br />
Embriología del Timo<br />
El timo deriva de la tercera bolsa faríngea externa e interna. En la semana 6 se observan ya primordios que empiezan<br />
a crecer y forman unas expansiones o evaginaciones tubulares. Estas masas pierden su conexión con la faringe y se<br />
ven entonces <strong>com</strong>o cordones macizos. Estos cordones se ven asociados a unas células mesenquimáticas que<br />
proceden de la cresta neural, que forman una cápsula y tabican el cordón. Las células epiteliales se separan unas de<br />
las otras y presentan un aspecto reticular. Siguen estando unidas por desmosomas porque siguen siendo aún células<br />
epiteliales, por lo que siguen teniendo filamentos de queratina 5 y 8. Hacia la semana 14 estas masas <strong>com</strong>ienzan a<br />
atraer células madre hematopoyéticas desde el saco vitelino, Esta células madre diferencian formando células<br />
dendríticas y macrófagos y las siguientes células que llegan ya diferencian hacia linfocitos T. Este es por tanto un<br />
proceso de formación de linfocitos T independiente de antígeno.<br />
Este proceso se continúa durante la vida fetal, derivando ya los linfocitos T de las células del hígado y no del saco<br />
vitelino. Una alteración en la relación epitelio-mesénquima produce un timo anormal, <strong>com</strong>o ocurre en el síndrome de<br />
DiGeorge.<br />
Estructura Histológica<br />
El timo tiene una cápsula conjuntiva delgada con tabiques que dividen el timo en lóbulos y lobulillos. No son tabiques<br />
<strong>com</strong>pletos, solo se extienden hasta el límite corti<strong>com</strong>edular. Además, NO hay fibras reticulares en el timo.<br />
El timo se divide en dos regiones: la corteza y la médula. La corteza es basófila y la médula es pálida y continua, por<br />
lo que los tabiques llegan solo hasta el límite corti<strong>com</strong>edular. No siempre se puede observar la continuidad en la<br />
médula del timo fetal.<br />
En la etapa adulta el timo tiene un estroma epitelial derivado de un precursor K5-K8 que aparece en las células<br />
reticuloepiteliales o epiteliorreticulares (CRE). En el timo ya maduro estas queratinas se expresan con la siguiente<br />
distribución: K8 en corteza y K5 en médula. Las células epiteliales expresan también dos factores de transcripción,<br />
Foxn1 y AIRE. Este estroma epitelial deja huecos donde se sitúan los linfocitos, macrófagos y células dendríticas.<br />
Foxn1 es fundamental para que las células epiteliales se puedan diferenciar en epiteliales de corteza y de médula.<br />
Regula también la expresión de las uniones de las células epiteliales, por lo que Foxn1 se debe estar expresando<br />
antes de que lleguen los linfocitos. Se expresa también en queratinocitos, <strong>com</strong>o se ha visto en ratones que tienen<br />
mutado el gen para Foxn1 y son desnudos y atímicos.
Página81<br />
Corteza<br />
La corteza tímica es una estructura basófila debido a su alta densidad celular. Está <strong>com</strong>puesta mayoritariamente por<br />
linfocitos, pero también se encuentran allí el resto de células inmunitarias.<br />
Las células reticulares de la corteza son en realidad células epiteliales estrelladas. Por debajo de la cápsula estas<br />
células se organizan formando una capa plana y continua que se continúa por debajo siguiendo los tabiques<br />
conjuntivos. Las CRE muestran lámina basal y forman una red similar a la que forman las células reticulares del bazo<br />
o de un ganglio linfático. Las células reticuloepiteliales quedan ocultas por la abundancia de linfocitos T.<br />
El núcleo de las CRE es grande y pálido, con 1 o 2 nucléolos, y su citoplasma es pálido. Están unidas por desmosomas<br />
y su queratina específica es K8. Además de formar una red, sintetizan una serie de factores <strong>com</strong>o la timopoyetiona y<br />
la timosina, esenciales para que los timocitos (linfocitos T) diferencien.<br />
Las células nodriza son CRE en la corteza del timo que presentan plegamientos de membrana plasmática, de modo<br />
que cada una de estas células nodriza alberga numerosos linfocitos en dichos pliegues.<br />
En el estroma de la corteza del timo no hay fibras reticulares, pero sí fibras colágenas.<br />
También se ha observado que puede haber algunos miocitos estriados que no parecen cumplir ninguna función, sino<br />
que son células que proceden de la cresta neural y han quedado ahí atrapadas.<br />
Los macrófagos y las células dendríticas interdigitantes expresan MHC-I y MHC-II, por lo que son esenciales para la<br />
diferenciación de linfocitos. Los macrófagos son PAS+ y se denominan células PAS positivas.<br />
A nivel cortical encontramos exclusivamente capilares que proceden de arteriolas del límite coriti<strong>com</strong>edular, desde<br />
donde salen capilares que ascienden por toda la corteza. Es ahí, en la corteza, donde encontramos la barrera<br />
hematotímica. Es una barrera crucial para conseguir aislar el ambiente del timo de la entrada de posibles antígenos<br />
que interfieran con la diferenciación de linfocitos T. La barrera hematotímica se <strong>com</strong>pone del endotelio de los<br />
capilares, una doble capa de células epiteliales, una doble lámina basal y un espacio mínimo conmacrófagos en él.<br />
Tipos de CRE en la corteza:<br />
CRE I<br />
Están entre la cápsula y las trabéculas rodeando los vasos corticales. Tienen núcleos polimorfos con nucleólos y se<br />
unen por uniones ocluyentes.<br />
CRE II<br />
Aparecen en la corteza media y tienen proyecciones largas y anchas; se unen por desmosomas y tienen un núcleo<br />
vesiculoso y citoplasma pálido. Presentan autoantígenos y CMH-I y CMH-II a los linfocitos T en desarrollo.<br />
CRE III<br />
Aparecen en la corteza profunda y en la unión corti<strong>com</strong>edular. Tienen proyecciones que les permiten unirse entre<br />
ellas y con las CRE adyacentes mediante uniones ocluyentes. Tienen núcleo y citoplasma denso.<br />
Médula<br />
La médula tímica se <strong>com</strong>pone de CRE, macrófagos, células dendríticas y linfocitos. Es más pálida que la corteza.<br />
Las células reticuloepiteliales son aquí mas abundantes que en corteza. Su queratina específica es K5. Sintetizan<br />
linfopoyetina estromal, que actúa sobre las células dendríticas medulares.<br />
En la médula del timo se observan los corpúsculos de Hassal: CRE que se cornifican y empiezan a agruparse<br />
formando capas hasta alcanzar los 30-150μm. Los corpúsculos de Hassal son muy basófilos y su número aumenta con<br />
la edad; tienen el centro muy queratinizado. No tienen una función clara asignada.<br />
Los macrófagos y las células dendríticas interdigitantes son muy abundantes en la médula tímica, especialmente en<br />
el límite corti<strong>com</strong>edular.
Página82<br />
Tipos de CRE en la médula:<br />
CRE IV<br />
Están en la médula superficial y unión corti<strong>com</strong>edular. Se unen entre ellas y con las CRE III mediante uniones<br />
ocluyentes. Tienen núcleo y citoplasma denso.<br />
CRE V<br />
Tienen núcleos polimorfos y cromatina perinuclear definida.<br />
CRE VI<br />
Forman los corpúsculos de Hassall. Tienen citoplasma pálido y están unidos por desmosomas con el centro<br />
cornificado-calcificado.<br />
Funciones<br />
Las células acceden al timo por los vasos situados en el límite corti<strong>com</strong>edular. En la corteza tiene lugar el proceso de<br />
diferenciación de linfocitos T, por lo que el 85% de éstos se encuentran allí. Además en la corteza tiene lugar una<br />
apoptosis masiva de linfocitos. Después de salir de la corteza, los linfocitos deben sufrir un proceso de maduración<br />
en la médula del timo para poder salir a la circulación. La maduración de linfocitos ocurre durante toda la vida del<br />
organismo, si bien sucede con más intensidad en la infancia.<br />
Maduración en corteza externa: selección positiva<br />
Los linfoblastos que llegan al límite corti<strong>com</strong>edular son precursores de linfocitos T originados en la médula ósea y no<br />
tienen marcadores CD4 ni CD8, es decir, son linfocitos T doble negativos. Se desplazan entonces hacia la zona<br />
subcapsular y empiezan a proliferar. Los linfoblastos son células del doble de tamaño que un linfocito normal.<br />
Los linfoblastos sufren entonces un proceso de reordenamiento génico, y empiezan a mostrar marcadores <strong>com</strong>o CD4<br />
o CD8 y un precursor de su receptor, el preTCR. La selección positiva de linfocitos es posible debido a su interacción<br />
con las células reticuloepiteliales. El <strong>com</strong>plejo MHC-I de esas células es clave para que el linfocito T exprese CD4 y el<br />
<strong>com</strong>plejo MHC-II para que exprese CD8. Si un linfocito T no reconoce alguno o ninguno de estos dos <strong>com</strong>plejos, el<br />
linfocito muere por apoptosis. Los linfocitos deben reconocer ambos <strong>com</strong>plejos para poder sobrevivir, es decir, de<br />
este proceso de selección positiva salen linfocitos CD4+/CD8+.<br />
Este proceso ocurre no en la zona subcapsular, sino a medida que los linfocitos van descendiendo por la corteza.<br />
Maduración en paracorteza: selección negativa<br />
A continuación los linfocitos sufren un proceso selección negativa o deleción clonal. En este proceso se consigue que<br />
los linfocitos doble positivo sean simple positivo, bien CD4+, bien CD8+. Las células reticuloepiteliales expresan el<br />
factor de transcripción AIRE, lo que les permite expresar todo tipo de antígenos del propio corazón, riñón, cerebro,…,<br />
es decir, expresan autoantígenos. Cuando el gen que codifica para AIRE muta, se produce la enfermedad APECED2 o<br />
APS-1. Existen otros problemas puntuales en los que los linfocitos reconocen solo un autoantígeno determinado,<br />
causando problemas en esa zona. Los linfocitos que interaccionan con los autoantígenos expuestos son eliminados<br />
por las células reticuloepiteliales, los macrófagos y las células dendríticas. Este proceso ocurre en la zona más<br />
profunda de la corteza.<br />
En el límite corti<strong>com</strong>edular los linfocitos, después de pasar por los dos procesos de selección, ya son diferenciados<br />
aunque no son aún inmuno<strong>com</strong>petentes. Para serlo, los linfocitos deben ir entonces a la médula ósea y sufrir una<br />
serie de cambios a lo largo de una semana, de modo que sean capaces de sintetizar sustancias propias de los<br />
linfocitos, <strong>com</strong>o las linfocinas y los factores.
Página83<br />
Regulación Hormonal de la Actividad Tímica<br />
La maduración de los linfocitos es posible gracias a una serie de hormonas paracrinas (sintetizadas por las CRE):<br />
timosina, timopoyetina, timulina y Factor Estimulante Tímico.<br />
Además, hay una serie de hormonas extratímicas <strong>com</strong>o:<br />
Somatotropina: estimula el desarrollo de los linfocitos corticales.<br />
Tiroxina: estimula a las CRE para la síntesis de timulina.<br />
Corticoides adrenales: inactivan la proliferización de linfocitos T.<br />
ACTH: estimula la síntesis de corticoides adrenales e indirectamente inhibe la proliferización de lf T.<br />
Vascularización<br />
Las trabéculas permiten la entrada de vasos sanguíneos y vasos linfáticos aferentes, pero no de eferentes.<br />
Barrera Hematotímica<br />
Los vasos sanguíneos pasan de las trabéculas al parénquima del timo a través de las partes más profundas de la<br />
trabécula, de modo que se llevan consigo una vaina de tejido conectivo que forma la vaina perivascular, cuyo grosor<br />
es directamente proporcional al diámetro del vaso. Donde es más gruesa hay fibras reticulares, fibroblastos,<br />
macrófagos y plasmocitos, pero en las zonas más finas sólo hay fibras reticulares y algunos fibroblastos (aquí SÍ hay<br />
fibras reticulares, porque es estroma).<br />
La vaina perivascular se encuentra rodeada de CRE de modo que se forma la barrera hematotímica entre la sangre y<br />
el parénquima tímico. Desde la luz vascular a las CRE aparecen:<br />
Endotelio vascular<br />
Lámina Basal Endotelial<br />
Lámina Perivascular<br />
Lámina Basal de las CRE<br />
CRE
Página84<br />
BAZO<br />
El bazo es un órgano intraperitoneal, situado en el hipocondrio izquierdo. Pesa aproximadamente 200 g (a partir de<br />
350 g se considera esplacnomegalia, frecuentemente asociada a trastornos inflamatorios, circulatorios o<br />
metabólicos). El bazo está interpuesto en la circulación sanguínea y no resulta indispensable para la vida. Por esta<br />
localización, el bazo es un filtro para células dañadas o extrañas de la sangre y es capaz asimismo de detectar<br />
antígenos circulantes, desarrollando respuestas inmunológicas ante ellos<br />
El bazo, a diferencia de los órganos linfoides vistos hasta ahora, no está estructurado en corteza y médula. Consta de<br />
un estroma que forma una cápsula y trabéculas y un parénquima o pulpa, de tipo rojo o blanco.<br />
Cápsula y Trabéculas<br />
Estas estructuras son de tejido conjuntivo denso con múltiples estratos donde es frecuente encontrar fibras elásticas.<br />
Estas fibras son importantes para la adaptación del bazo a los cambios de volumen sanguíneo. Hay miocitos lisos, por<br />
lo que la cápsula es contráctil, permitiendo la reserva de sangre en el bazo.<br />
En el hilio, la cápsula se engruesa y es muy friable. De la cápsula salen trabéculas que dan rigidez al bazo y lo dividen<br />
en secciones <strong>com</strong>unicadas.<br />
Pulpa<br />
La pulpa esplénica está formada por dos zonas funcionalmente diferentes, la pulpa roja y la pulpa blanca, cada una<br />
con su propio estroma reticular.<br />
La pulpa roja en fresco es de color rojo intenso y supone un 75% del parénquima. Está formada por muchos<br />
sinusoides entre los que quedan células formando bien placas, bien cordones de Billroth. La pulpa roja es la<br />
encargada del filtrado de la sangre.<br />
La pulpa blanca supone un 25% del parénquima esplénico que en fresco presenta tonalidad grisácea. Es una<br />
estructura muy basófila y se dispone <strong>com</strong>o áreas dispersas en la pulpa roja. Tiene una función eminentemente<br />
inmunológica.<br />
Pulpa roja<br />
La pulpa roja está formada por una red de sinusoides conectados entre sí. El endotelio de los sinusoides lo forman<br />
células alargadas denominadas costillas que apenas contactan entre sí y solo tienen uniones de sellado en sus<br />
extremos. Su eje principal se dispone paralelo al eje mayor del sinusoide. Bajo las costillas queda una lámina basal<br />
discontinua. Esta lámina basal se orienta distinta a las células: en perpendicular al eje mayor del sinusoide, en forma<br />
de aros. También se encuentran aquí fibras reticulares. Estos sinusoides carecen de pericitos o células contráctiles.<br />
Los cordones de Billroth son el conjunto de células dispuestas entre los sinusoides. Son fundamentalmente células<br />
plasmáticas, macrófagos y elementos formes de la sangre. Las prolongaciones de los macrófagos de los cordones<br />
penetran en los sinusoides. En esta zona externa a los sinusoides también encontramos estroma reticular.<br />
Pulpa blanca<br />
La pulpa blanca consta de áreas dispersas en la pulpa roja. Estas áreas son basófilas por la heterocromatina de la<br />
vaina de linfocitos asociada a cada arteria central que penetra. Esta vaina se denomina vaina linfática periarterial,<br />
VLP o PALS y se sitúa en la túnica adventicia de la arteria. En la VLP hay pequeños nódulos de hasta 1 cm llamados<br />
nódulos esplénicos o corpúsculos esplénicos de Malpighi; estos nódulos tienden a desplazar a la arteria central de<br />
modo que adquiere una posición más excéntrica de la que le corresponde. La VLP está <strong>com</strong>puestA sobre todo de<br />
linfocitos T, mientras que el nódulo lo hace sobre todo de linfocitos B.<br />
En el límite de la VLP con la pulpa roja encontramos la zona marginal con un estroma reticular laxo ricamente<br />
irrigada y con linfocitos B, T y macrófagos.
Página85<br />
Vascularización<br />
La arteria esplénica entra al bazo, se ramifica en arterias trabeculares y éstas pasan a la pulpa roja. Si se rodea de<br />
linfocitos, se forma la PALS y hablamos ya de pulpa blanca, es decir, arterias centrales, que son realmente arteriolas.<br />
La arteria central da arteriolas radiales que irrigan los corpúsculos de Malpighi y llegan a la zona marginal. La arteria<br />
central continua en su zona distal y acaba dando ramas terminales en la pulpa roja (arteriola penicila). Estas<br />
arteriolas constan tan solo de una capa de miocitos y un endotelio alto.<br />
Cada arteriola penicila da lugar a un capilar. Estos capilares son de endotelio alto, rodeado de una vaina de células<br />
reticulares y macrófagos. Esta vaina celular se denomina elipsoide y los capilares así conformados se llaman capilares<br />
envainados. Estos capilares acaban abriéndose de modo que los elementos formes de la sangre quedan en los<br />
cordones de Billroth (circulación abierta). La sangre que no se vierte a los cordones llega a la zona marginal y se<br />
vierte al seno marginal para después pasar a la pulpa roja. Una cierta cantidad de sangre va del seno marginal al seno<br />
cavernoso perimarginal, desde donde es drenada directamente por vénulas (circulación cerrada). La sangre de los<br />
cordones entra entre las costillas de los sinusoides y la sangre es drenada por vénulas, venas de la pulpa, venas<br />
trabeculares y venas esplénicas.<br />
El 90% del flujo esplénico forma parte de la circulación cerrada y tarda 2 min en cruzar el bazo. Sin embargo, en un<br />
momento dado, la mayoría de la sangre esplénica está en circulación esplénica o abierta y tarda entre 30 y 60<br />
minutos en salir del bazo, permitiendo así la filtración.<br />
Cada zona del bazo es irrigada por arterias terminales. Cuando se produce algún problema en alguna de estas<br />
arterias, se forma una cicatriz. A diferencia de lo que ocurre en otros órganos, en el bazo las arterias adyacentes no<br />
son capaces de cubrir las necesidades de esta zona infartada.<br />
Los vasos linfáticos se sitúan en la pulpa blanca y drenan la linfa dando paso a otros vasos linfáticos más grandes en<br />
las trabéculas y la cápsula. De ahí la linfa sale hacia los ganglios linfáticos situados próximos al hilio esplénico.<br />
Funciones<br />
El bazo filtra la sangre, forma células linfoides, elimina o inactiva antígenos de origen sanguíneo, destruye plaquetas<br />
y eritrocitos envejecidos y participa en la hematopoyesis.<br />
Durante la etapa fetal, la pulpa roja es capaz de albergar procesos de eritropoyesis, concretamente durante el<br />
segundo trimestre de la gestación. En la especie humana, esta función ya ha desaparecido en el niño y el adulto, si<br />
bien en otras especies se mantiene a lo largo de la vida del organismo.
Página86<br />
No obstante, una de las funciones principales del bazo es la de filtrar la sangre de células envejecidas o extrañas.<br />
Dentro de este papel destaca el proceso de hemólisis, esto es, la destrucción de eritrocitos y trombocitos anormales<br />
o envejecidos.<br />
La pulpa roja es además encargada de la eliminación de bacterias y virus por acción directa de macrófagos o por<br />
opsonización de los patógenos, evitando así procesos de sepsis y constituyendo una eficaz medida de prevención de<br />
infecciones renales, meníngeas y pulmonares.<br />
La pulpa roja actúa también <strong>com</strong>o reservorio de plaquetas, quedando un tercio del total en el bazo, constituyendo el<br />
denominado pool de reserva. En otras especies, el bazo actúa además <strong>com</strong>o reservorio de eritrocitos, si bien está<br />
función no se da en humanos, donde sólo un 3% de la población eritrocitaria se aloja en el bazo.<br />
En la función esplénica del filtrado de sangre tienen un importante papel los macrófagos de los cordones de Billroth<br />
en la pulpa roja, encargados de eliminar células dañadas, envejecidas o alteradas, así <strong>com</strong>o partículas o células<br />
extrañas.<br />
Por su parte, el proceso de hemólisis es posible debido a que los eritrocitos tienen la capacidad de deformarse, de<br />
modo que pueden salir de los sinusoides a los cordones esplénicos a través de las hendiduras que presentan las<br />
costillas y la lámina basal de dichos sinusoides. Los eritrocitos dañados, envejecidos o deteriorados pierden esta<br />
capacidad de deformarse, de modo que al intentar atravesar estas hendiduras de los sinusoides se rompen, de modo<br />
que sus restos son fagocitados por los macrófagos.<br />
El hierro de los eritrocitos es recuperado por el bazo y es transportado a la médula ósea unido a la transferrina, si<br />
bien también pueden observarse inclusiones de hierro en los macrófagos esplénicos en forma de ferritina o<br />
hemosiderina.<br />
El grupo hemo de la hemoglobina eritrocitaria es degradado a bilirrubina, la cual es transportada hasta el hígado<br />
unida a la albúmina y es después excretada con la bilis.<br />
En la pulpa blanca reside la función inmunológica, pues es aquí donde se desencadenan respuestas inmunitarias<br />
frente a antígenos. Los antígenos captados por células dendríticas interdigitantes son llevados hasta las PALS, donde<br />
activan linfocitos Tc y Th. Estos últimos activan linfocitos B, los cuales se transforman en células plasmáticas y migran<br />
hacia el nódulo primario, desplazando a los linfocitos T que allí había y proliferando, formando así un nódulo<br />
secundario donde se desarrolla una respuesta inmunitaria de tipo humoral.<br />
MÉDULA ÓSEA<br />
La Médula Ósea (MO) supone el 3,5-6% del peso corporal y se <strong>com</strong>pone de vasos sanguíneos sinusoides y una malla<br />
o red de células hematopoyéticas.<br />
La célula adventicia o reticular emite prolongaciones laminares hacia el interior de la sustancia de los cordones<br />
hematopoyéticos proporcionando sostén a las células sanguíneas en desarrollo. Además las células reticulares<br />
producen las fibras de colágeno III o reticulares y estimulan la diferenciación de las células pluripotenciales<br />
sanguíneas mediante la secreción de Factores Estimulantes de Colonias (FEC).<br />
Según va creciendo la célula sanguínea esta va desplazando a las células reticulares y se abre paso en el endotelio del<br />
sinusoide creando una abertura transitoria para el pasaje transcelular.<br />
Los cordones contienen además macrófagos, mastocitos y células adiposas; estas células aumentan en la médula<br />
ósea amarilla, un tipo de MO inactiva que retiene su potencial hematopoyético y puede volver a ser MO roja si es<br />
necesario. La MO gris es exclusiva en ancianos y patología.<br />
En la matriz extracelular de la MO aparecen fibras reticulares, colágeno I, laminina, fibronectina y hemonectina.
Página87<br />
SISTEMA TEGUMENTARIO<br />
La piel es el órgano mayor del cuerpo, con una superficie de 2m 2 , formando la cubierta corporal. Se introduce<br />
también en zonas mucocutáneas y supone un 16% de nuestro peso corporal. El grosor medio de la piel es de 0,5 mm,<br />
bien puede alcanzar hasta los 3 o 4 mm. Es de fácil exploración y biopsia y tiene un alto valor diagnóstico, no solo en<br />
trastornos dermatológicos.<br />
La piel consta de una capa externa y una interna.<br />
La capa externa recibe el nombre de epidermis y es de origen epitelial. La capa interna es de naturaleza conjuntiva y<br />
origen mesodérmico, y recibe el nombre de dermis o corion. Es aquí donde encontramos los anejos cutáneos o<br />
faneras, que son de origen epitelial.<br />
La piel descansa sobre la capa subcutánea conocida <strong>com</strong>o hipodermis, donde pueden llegar a introducirse algunos<br />
anejos.<br />
Funciones<br />
<br />
<br />
<br />
<br />
Protección: puede ser mecánica (daños físicos), química (deshidratación), física (rayos UV), biológica (barrera<br />
física, respuesta inmunitaria)…<br />
Metabolismo: síntesis de vitamina D y excreción de catabolitos, principalmente agua y sales.<br />
Termorrecepción: la piel es el órgano sensitivo más extenso y permite la termorregulación (homeotermos).<br />
Presenta receptores de diversos estímulos<br />
Absorbe sustancias liposolubles y alcohólicas, lo que permite la aplicación cutánea de tratamientos tópicos<br />
(cremas, pomadas, parches, liposomas…)<br />
Tipos de Piel<br />
La piel gruesa aparece en palma de manos y pies, donde hay mayor exposición a la abrasión, carecen de pelos y<br />
tienen una capa epidérmica más gruesa. La epidermis de esta zona es más gruesa porque el estrato córneo es de<br />
gran grosor.<br />
La piel fina recubre el resto de la superficie corporal y posee una epidermis mucho más delgada, con folículos pilosos<br />
en prácticamente toda su extensión.<br />
EPIDERMIS<br />
La epidermis es un epitelio plano estratificado queratinizado que deriva del ectodermo. La epidermis mide 0,4-1,5<br />
mm de espesor y en función de su grosor hablamos en histología de piel fina o gruesa.<br />
Se renueva constantemente. En ella se distinguen cuatro tipos celulares: los queratinocitos (80%), los melanocitos,<br />
las células de Langerhans y las células de Merkel.<br />
Los queratinocitos se organizan en cuatro estratos de tal forma que la puesta en marcha de su programa genético de<br />
diferenciación les lleva a desplazarse desde niveles basales a suprabasales a la vez que ocurren toda una serie de<br />
cambios a nivel estructural, ultraestructural y metabólico.<br />
Los otros tipos celulares aparecen intercalados entre los queratinocitos, pero no de forma aleatoria: los melanocitos<br />
y las células de Merkel permanecen siempre en la capa basal, mientras que las células de Langerhans se sitúan en<br />
estratos superiores.<br />
Los estratos de la epidermis son, a grandes rasgos:<br />
Estrato Basal o Germinativo: única capa de células madre o stem cells que por mitosis permiten la<br />
regeneración de los queratinocitos; tiene una gran basofilia por la aglomeración de núcleos.<br />
Estrato Espinoso: formado por varias capas de queratinocitos unidos por unos desmosomas localizados en<br />
unas prolongaciones en forma de espina que hacen característico este estrato.
Página88<br />
<br />
<br />
<br />
Estrato Granuloso: es la capa más superficial de la porción no queratinizada de la epidermis y está formada<br />
por una cantidad variable de queratinocitos que contienen gránulos de queratohialina.<br />
Estrato Lúcido: exclusivo de la piel gruesa, tiene aspecto refringente.<br />
Estrato Córneo: es la capa que mas varía de grosor (de piel fina a piel gruesa).<br />
Queratinocitos<br />
Los queratinocitos suponen el 80% del total celular epidérmico y, durante su ascensión por la epidermis hacia el<br />
estrato córneo, sufren un proceso de diferenciación que incluye cambios estructurales, ultraestructurales y<br />
metabólicos que los terminan conduciendo a la apoptosis. Es por ello por lo que los queratinocitos son las células<br />
que determinan las capas mencionadas anteriormente.<br />
Los queratinocitos presentan un importante desarrollo de tonofilamentos, cuya expresión génica está altamente<br />
controlada. Los tonofilamentos son filamentos intermedios de queratina (K) asociada en pares formados por una<br />
queratina ácida y una básica o neutra. El tipo de queratina del tonofilamento varía en función del tipo de epitelio y<br />
del grado de diferenciación.<br />
El proceso de diferenciación del queratinocito implica el cumplimiento de dos funcinoes:<br />
Producción de K, que llega a ocupar toda la célula.<br />
Crear una barrera contra el agua.<br />
La diferenciación de los queratinocitos implica un aumento del tamaño celular, tendencia a una morfología plana,<br />
regulación de la expresión génica de pares de K, aparición de nuevos orgánulos, reorganización de su ultraestructura,<br />
cambios metabólicos, cambios en la membrana plasmática, apoptosis y deshidratación.<br />
Al final del proceso de diferenciación, encontramos escamas repletas de filamentos de queratina con membrana<br />
plasmática que acaba rompiéndose, quedando solo una envuelta que se ha formado bajo la membrana. Entre las<br />
escamas se forma una matriz lipídica.<br />
Este proceso de diferenciación hace que distingamos cuatro estratos distintos en la epidermis (basal, espinoso,<br />
granuloso y córneo). Los queratinocitos vivos, correspondientes a los tres primeros estratos, conforman el estrato<br />
mucoso de Malpighi.<br />
ESTRATO BASAL<br />
Es una capa única de células cúbicas, basófilas, con abundantes orgánulos, <strong>com</strong>o ribosomas libres, un pequeños<br />
Aparato de Golgi y RER.<br />
Expresan el par K5/K14 y presentan bajos niveles de Ca 2+ por la presencia de la proteína ligadora de calcio.<br />
Las células de este estrato presentan uniones de adhesión con la lámina basal; algunas de estas uniones de adhesión<br />
son marcadores de estas células (α 3 β 1 , α 6 β 4 ). Encontramos también hemidesmosomas y desmosomas.<br />
El 10% del estrato se <strong>com</strong>pone de células madre encargadas de renovar la epidermis. El 50% son células<br />
amplificadoras, que mantienen su capacidad de división durante mucho tiempo para amplificar la renovación<br />
iniciada por las células madre. El 40% restante son células postmitóticas, ya preparadas para ascender a estratos<br />
superiores.<br />
El estrato basal renueva la epidermis y forma los anejos cutáneos. Al conjunto formado por una célula basal y todas<br />
sus derivadas se le denomina unidad proliferativa epidérmica.<br />
ESTRATO ESPINOSO<br />
La célula postmitótica del estrato basal rompe las uniones con las células adyacentes y asciende llegando al estrato<br />
espinoso, estrato formado por entre 5 y 10 capas de células. Las células del estrato espinoso presentan una serie de<br />
espinas, prolongaciones donde se anclan los tonofilamentos y se convierten en puntos de unión entre dos células<br />
vecinas, además de ser uniones de <strong>com</strong>unicación. Estas espinas presentan un engrosamiento central conocido <strong>com</strong>o<br />
nódulos de Bizzozzero.<br />
Las células del estrato espinoso dejan entre si espacios bastante amplios, dando lugar a los canales faciales. En los<br />
puntos donde las células sí contactan, encontramos desmosomas entre las células, formados por desmogleína tipo 3.
Página89<br />
En el estrato espinoso se desarrollan tonofilamentos del par K1/K10, muy eficaces en llevar a cabo funciones de<br />
unión, permitiendo cohesionar el estrato e impidiendo su destrucción. Por otro lado, los genes del par K6/K16 se<br />
expresan en toda capa basal de cualquier epitelio, aunque la traducción del gen solo se produce cuando es necesaria<br />
la regeneración del epitelio (heridas, psoriasis, queratosis…); es decir, sólo en situaciones excepcionales es posible<br />
encontrar K6 y K16 en este estrato. En este estrato también aparece la proteína involucrina.<br />
Las células de estrato espinoso van creciendo y aparece en ellas un nuevo orgánulo, los cuerpos de Odland,<br />
lamelares o queratinosomas. Son gránulos con un contenido dispuesto en lamelas, formado por glucoproteínas,<br />
glucolípidos, fosfolípidos, esteroles e hidrolasas.<br />
Las células de este estrato presentan una gran cantidad de polirribosomas y se pueden evidenciar los primeros<br />
gránulos de queratohialina en la parte más superficial.<br />
ESTRATO GRANULOSO<br />
El estrato granuloso no es tan grueso <strong>com</strong>o el espinoso. Las células de este estrato producen inicialmente<br />
tonofilamentos K1/K10, que según son sintetizados son fosforilados, de modo que se detectan tonofilamentos<br />
K2/K12. La célula sintetiza una sustancia que embebe y aglutina la gran cantidad de tonofilamentos sintetizados,<br />
formando una matriz de filagrina que, junto con los tonofilamentos, conforma los gránulos de queratohialina,<br />
basófilos, irregulares y sin membrana.<br />
Se produce en este estrato la síntesis de otros materiales que se organizan bajo la membrana plasmática formando la<br />
banda marginal, que se desarrolla para formar después una envuelta cornificada. Está <strong>com</strong>puesta de lípidos unidos<br />
mediante enlaces covalentes, formando una estructura muy insoluble, con un grosor de entre 7 y 15 nm.<br />
En esta envuelta cornificada encontramos diversas proteínas proteínas: involucrina, queratohialina, loricrina, elafina,<br />
repetina, desmoplaquina, envoplaquina, periplaquina, cistatina α, pancornulinas, pequeñas proteínas ricas en<br />
prolina…<br />
La involucrina se sintetiza libre en el citosol y migra a la membrana plasmática, donde se une a ciertas regiones<br />
interdesmosómicas de algunas proteínas. La loricrina, sintetizada en gránulos L, es dominante al final de la vida del<br />
queratinocito, suponiendo un 70% del peso de la envuelta.<br />
Los cuerpos de Odland se siguen sintetizando en este estrato y suponen ya un 15% del volumen celular, de modo que<br />
empiezan a exocitarse. Las enzimas extracelulares y del cuerpo de Odland transforman el material lamelar en<br />
láminas que se disponen entre los queratinocitos. Ese material se expande y es alterado químicamente, formando<br />
finalmente una barrera de propiedades hidrofóbicas.<br />
El proceso de diferenciación requiere un interior celular rico en calcio, pero el queratinocito tiene niveles bajos del<br />
mismo. A partir del estrato granuloso, el queratinocito ya no sintetiza la proteína ligadora de calcio, de manera que<br />
el calcio que estaba asociado a esta proteína deja de estarlo y suben los niveles de calcio, lo que permite que tengan<br />
lugar las transformaciones.<br />
Al final del proceso de diferenciación, la célula entra en apoptosis. Sintetiza una serie de enzimas endonucleasas,<br />
pierde orgánulos, vierte enzimas que lisan <strong>com</strong>ponentes celulares... Algunas enzimas están sintetizadas desde<br />
estratos anteriores, pero están bloqueadas hasta este momento. La célula que aparece en el estrato más superficial<br />
de esta capa ha perdido el núcleo, gran cantidad de sus organelas y presenta una gran concentración de eleidina en<br />
su interior.<br />
La célula va perdiendo peso durante el proceso de transición del estrato granuloso al estrato córneo. El ascenso<br />
hasta el estrato córneo por el estrato de Malpighi dura 14 días, y el ascenso por estrato córneo dura otros 14 días,<br />
haciendo un total de 28.<br />
El estrato lúcido es una zona justo por encima del estrato granuloso, de afinidad tintorial diferente a éste. No es un<br />
estrato real, sino un concepto de microscopía óptica.<br />
ESTRATO CÓRNEO<br />
Este estrato está formado por escamas cornificadas o corneocitos. El corneocito es una especie de saco repleto de<br />
queratinas, rodeado por una membrana plasmática reforzada por una envuelta cornificada. Estos sacos están<br />
<strong>com</strong>pactados gracias a una matriz extracelular que es altamente hidrofóbica.
Página90<br />
Los corneocitos son grandes y aplanados y no presentan núcleo, salvo en alteraciones <strong>com</strong>o psoriasis o<br />
paraqueratosis. Son marcadamente eosinófilos.<br />
El estrato córneo no es un estrato inerte, ya que en él se mantienen actividades metabólicas: anclaje de corneocitos,<br />
maduración de la envuelta cornificada y cambios en la matriz extracelular El estrato córneo permite que la<br />
descamación tenga lugar en el momento preciso.<br />
Es además una barrera mecánica que protege a las capas inferiores, y una barrera hídrica que evita la deshidratación<br />
de la epidermis.<br />
El estrato córneo se divide en una zona más profunda, el estrato conyunto o <strong>com</strong>pacto; y una más superficial, el<br />
estrato disyunto o descamado.<br />
El estrato conyunto es una parte del estrato córneo donde no se produce descamación, ya que los corneocitos están<br />
unidos mediante desmosomas formados por desmogleína 1. La envuelta cornificada bajo la membrana es todavía<br />
frágil e inmadura.<br />
Alteraciones en este estrato provocan una disolución prematura de desmosomas, lo que genera una descamación<br />
temprana y alteración de la barrera hídrica.<br />
En el estrato disyunto o descamativo se produce la disolución de los desmosomas, generando huecos en la matriz<br />
extracelular que forman una red de canales por donde pueden circular elementos hidrófobos. La envuelta<br />
cornificada es ya más rígida, con un grosor máximo de 15 o 16 nm.<br />
Se empieza a disolver la membrana plasmática y la matriz extracelular sufre una serie de cambios moleculares: no se<br />
encuentran azúcares ni líquidos polares, que son degradados, pero sí se hallan abundantes lípidos neutros, esteroles<br />
y ceramidas, haciendo que la matriz sea más hidrofóbica.<br />
Así pues, en esta parte del estrato córneo se observa una alternancia de corneocitos y matriz extracelular. Los<br />
corneocitos pierden la membrana plasmática y quedan envueltos exclusivamente por la envuelta cornificada rígida.<br />
La matriz extracelular es muy hidrofóbica, con canales por donde circula material hidrófilo.<br />
La descamación se produce finalmente por la disolución de los desmosomas y la acción de enzimas que degradan la<br />
membrana plasmática, la envuelta y la matriz extracelular.<br />
Finalmente, aparece una última capa que se denomina manto ácido: se sitúa sobre el estrato córneo y está formado<br />
por las escamas, la matriz extracelular, el sudor ecrino y el sebo Lubrifica y acidifica la superficie, de modo que<br />
permite que se alojen determinados gérmenes saprófitos en la piel, que aseguran que la piel no sea colonizada por<br />
otros gérmenes dañinos. Los jabones alcalinos y los detergentes dañan el manto ácido.
Página91<br />
Melanocitos<br />
Los melanocitos son células que derivan de la cresta neural. Alcanzan la dermis hacia las semanas 10 u 11 del<br />
desarrollo y en la semana 12 o 14 ya están en la epidermis. Sintetizan melanina a lo largo de toda la vida del<br />
individuo.<br />
La melanina es un pigmento argirófilo, de modo que mediante sales argénticas se impregnan los melanosomas, es<br />
decir, se detectan melanocitos y queratinocitos. Para detectar solo melanocitos por tinción histoquímica, se localizan<br />
enzimas que participan en la síntesis de melanina, <strong>com</strong>o la tirosinasa DOPA. También se usa la tinción de Masson-<br />
Fontana.<br />
Los melanocitos se asientan sobre la membrana basal, si bien, ante un estímulo sostenido, pueden aparecer<br />
melanocitos en estratos superiores. Los melanocitos aunque no muestran ni hemidsmosomas ni desmosomas que<br />
los unan con queratinocitos. Sí encontramos, no obstante, bandas basales electrodensas.<br />
Sus prolongaciones se llaman dendritas, son ascendentes, y se meten entre los queratinocitos del estrato espinoso.<br />
El núcleo celular es ovalado y se sitúa en paralelo a la lámina basal. No presentan tonofilamentos, pero sí filamentos<br />
intermedios y micro túbulos. Sintetizan y depositan la melanina en los melanosomas.<br />
Existen dos tipos de pigmentos sintetizados por el melanocito: la eumelanina es un pigmento marrón y la<br />
feomelanina es anaranjada o amarillenta.<br />
A lo largo de su diferenciación para ser maduros y funcionales, los melanocitos presentan cuatro fases:<br />
FASE I: los melanocitos sintetizan los premelanosomas, que se identifican histoquímicamente porque contienen<br />
tirosinasa.<br />
FASE II: se observa un interior celular no homogéneo, apareciendo membranas internas o lamelas.<br />
FASE III: se sintetiza melanina que se deposita sobre las membranas internas que aparecen en la fase II. La melanina<br />
se forma a partir de la oxidación de la tirosina a 3,4-dihidroxifenilalanina o DOPA, que por acción de la tirosinasa se<br />
transforma en melanina.<br />
FASE IV: ya aparecen melanosomas que no tienen estriaciones y son ya electrodensos. Se dan cambios de la matriz<br />
extracelular y en las enzimas.<br />
Los melanosomas tienen pH ácido y, aquellos con eumelanina son más alargados y con la matriz estriada, mientras<br />
que los melanocitos con feomelanina no presentan lamelas, sino una matriz de tipo granuloso, con melanosomas<br />
redondos.<br />
Los melanosomas se sintetizan en el soma del melanocito y se transportan a lo largo de las dendritas vía kinesina, es<br />
decir, por transporte asociado a microtúbulos;también vuelven hacia el soma por el mismo mecanismo.<br />
Los melanosomas deben ser retenidos en los extremos más distales de las dendritas (retención periférica). En este<br />
proceso intervienen los microtúbulos y una serie de proteínas <strong>com</strong>o Rab27a o miosina Va.<br />
Los melanosomas son trasferidos entonces a los queratinocitos adyacentes. De hecho, la mayoría de la melanina se<br />
detecta en queratinocitos, que contienen gránulos de melanina. Estos gránulos son en realidad la imagen que se<br />
observa al óptico de la melanina en estas células.<br />
En los queratinocitos hay <strong>com</strong>plejos delimitados por membranas en los cuales encontramos melanosomas. Estos<br />
<strong>com</strong>plejos se fusionan con lisosomas formando lisosomas secundarios. En individuos de raza blanca, cada <strong>com</strong>plejo<br />
tiene múltiples melanosomas, mientras que en la raza negra hay un único melanosoma de gran tamaño.<br />
En el queratinocito es donde la melanina cumple su función: protección ante radiaciones UV, por lo que la melanina<br />
se sitúa por encima de los núcleos de los queratinocitos.<br />
Estos melanosomas en el queratinocito empiezan a ser degradados a lo largo de la diferenciación de queratinocitos.<br />
En la raza blanca no encontramos melanosomas más allá de estrato granuloso, mientras que en la raza negra sí se<br />
observan más allá.<br />
La unidad epidermomelánica hace referencia a la consideración del melanocito <strong>com</strong>o célula secretora, de modo que<br />
cada unidad se <strong>com</strong>pone de un melanocito y los, aproximadamente, 36 queratinocitos a los que transfiere los<br />
melanosomas. En <strong>com</strong>paración, existe un melanocito para cada 4 a 10 queratinocitos.<br />
La cantidad de melanocitos en todas las razas es igual.<br />
El número de melanocitos es distinto en distintas zonas de la piel. Estas diferencias zonales están marcadas<br />
genéticamente. Con la edad se pierden melanocitos con la edad.
Página92<br />
Células de Langerhans<br />
Estas células fueron descritas por Paul Langerhans en 1868. Se tiñen con impregnaciones áuricas (cloruro de oro), y<br />
también se detectan por la tinción inmunohistoquímica de la ATPasa o por inmunocitoquímicas CD1 y CD100<br />
(también por CD4, CD45). Suponen un 2-4% de las células epidérmicas.<br />
Las células de Langerhans derivan de precursores que encontramos en la médula ósea, desde donde migran,<br />
alcanzando la epidermis en la semana 6 o 7 del desarrollo.<br />
Se localizan en el estrato espinoso, es decir, en mitad de queratinocitos que ascienden, por lo que ellas deben<br />
intentar permanecer en el lugar, presentando un movimiento contracorriente.<br />
Tienen un soma estrellado, de manera que sus prolongaciones se sitúan entre los queratinocitos. Tienen un<br />
citoplasma pálido con muchos orgánulos, núcleo lobulado muy basófilo, y no presentan tonofilamentos ni<br />
desmosomas que los unan con queratinocitos. Presentan abundancia de cadherinas E.<br />
El orgánulo típico de la célula de Langerhans es el gránulo de Birbeck, con una característica forma de raqueta de<br />
tenis. En su interior encontramos la langerina, una lectina que permite reconocer carbohidratos.<br />
También encontramos células de Langerhans en órganos linfoides y en epitelios estratificados planos. Su número<br />
varía en función de la zona y disminuye con la edad.<br />
Además, posee receptores de superficie Fc, Ia y C3.<br />
La célula de Langerhans es una célula presentadora de antígenos. Cuando la célula de Langerhans reconoce un<br />
antígeno, sufre una serie de cambios: deja de ser una célula presentadora de antígenos, incrementa de tamaño,<br />
pierde los gránulos de Birbeck, las cadherinas E y diversos marcadores.<br />
Las células de Langerhans migran entonces hacia los vasos linfáticos, atravesando los canales faciales, la lámina basal<br />
y alcanzando la dermis. Llegan al ganglio linfático, activando linfocitos y dando lugar a una respuesta inmunológica,<br />
de tal manera que los linfocitos activados salen de los ganglios y acuden a aquella zona de la epidermis donde se<br />
captó el antígeno inicialmente. Parte de los linfocitos quedan en la epidermis (suponen un 0.5% del total<br />
epidérmico), sobre todo en alergias de contacto.<br />
Así, entre las funciones de las células de Langerhans encontramos:<br />
· Inducen respuestas específicas contra antígeno<br />
· Activan linfocitos Tc<br />
· Vigilan contra ataques de gérmenes infecciosos<br />
· Están implicadas en el rechazo a injertos<br />
Las células forman parte del SALT, donde están implicados los linfocitos T residentes y los queratinocitos, ya que las<br />
células de Langerhans provocan en el queratinocito la capacidad de síntesis de citocinas que modifican a las células<br />
de Langerhans.<br />
Células de Merkel<br />
La célula de Merkel se puede observar por impregnación aúrica y mediante la IHQ de sinaptofisina. Son células<br />
epidérmicas modificadas que se sitúan en el estrato basal de la epidermis. Su número es escaso, más abundante en<br />
la piel gruesa o los labios. Son de origen ectodérmico y se sitúan en su localización antes que las fibras nerviosas del<br />
disco, por lo que las células de Merkel actúan <strong>com</strong>o foco de atracción de la fibra nerviosa. Su citoplasma es claro, con<br />
núcleo lobulado, desarrollo alto de tonofilamentos (K8-K18 ó K8-K19), algunos melanosomas en su citoplasma<br />
ligeramente más denso que el de las células de Langerhans y hemidesmosomas y desmosomas con los queratinocitos<br />
vecinos.<br />
Puesto que junto a ella se localiza una fibra nerviosa, la célula de Merkel tiene gránulos neurosecretores de núcleo<br />
denso de 80nm con determinados neurotransmisores (VIP, met-encefalina y bombesina).<br />
La célula de Merkel esta asociada al bulo terminal expandido de una fibra nerviosa mielínica aferente formando el<br />
disco de Merkel, un mecanorreceptor de adaptación lenta. Se trata de una terminación nerviosa de extremo dilatado<br />
que atraviesa la lámina basal y llega al extremo basal de la epidermis, disponiéndose enfrentada a una célula de<br />
Merkel.
Página93<br />
DERMIS<br />
La dermis es la capa más interna de la piel. Es un tejido conjuntivo de origen mesodérmico con función<br />
eminentemente de sostén y de albergar los anejos cutáneos. Presenta dos sustratos, uno papilar y otro reticular.<br />
Unión Dermoepidérmica<br />
La unión dermoepidérmica es la frontera entre la dermis y la epidermis. Se <strong>com</strong>pone de la membrana basal de los<br />
queratinocitos del estrato basal de la epidermis y de la lámina basal subyacente a éstos.<br />
La unión dermoepidérmica permite cohesionar la dermis y la epidermis. Además, es la responsable de la polaridad de<br />
los queratinocitos basales y de la organización de citoesqueleto; y proporciona señales tanto durante el desarrollo<br />
<strong>com</strong>o durante la reparación de la piel tras una lesión. Es una barrera semipermeable.<br />
Si bien en la generación de la lámina basal colaboran los fibroblastos de la dermis, la lámina basal es formada<br />
principalmente por los propios queratinocitos del estrato basal epidérmico. En la lámina basal se distinguen tres<br />
zonas: la lámina lúcida, la lámina densa y la lámina reticular.<br />
La lámina lúcida se <strong>com</strong>pone de glucoproteínas que interaccionan con receptores de membrana tipo integrina y son<br />
responsables de la función de cohesión de la lámina.<br />
En la lámina densa encontramos fibras de colágeno IV y VII. El colágeno IV forma una red que da soporte y<br />
flexibilidad a la lámina. Encontramos asimismo proteglucanos, formando una red que almacena agua y actúa <strong>com</strong>o<br />
filtro o barrera en función de tamaño y carga de las partículas (impide el paso de moléculas mayores de 40 kDa). El<br />
colágeno VII forma haces que se dirigen hacia la dermis y se insertan en placas de anclaje.<br />
La lámina reticular presenta colágeno tipo I, III, V y VI, así <strong>com</strong>o fibras elásticas.<br />
Las llamadas fibras de oxitalán están constituidas por los <strong>com</strong>ponentes periféricos microfibrilares cubiertos por<br />
elastina soluble. Son elementos flexibles que permiten la a<strong>com</strong>odación ante deformaciones del tejido cutáneo sin<br />
<strong>com</strong>prometer su integridad estructural.<br />
A mayor aumento se pueden diferenciar unas estructuras entre la dermis y la epidermis: papilas dérmicas y redes de<br />
crestas epidérmicas.<br />
Las papilas dérmicas se proyectan desde la dermis hacia la epidermis y se <strong>com</strong>plementan con proyecciones<br />
epidérmicas hacia la dermis que forman las crestas epidérmicas. Estas estructuras son más frecuentes en sitios de<br />
mayor estrés mecánico y además son más largas y mucho más densamente agrupadas.<br />
Dermis Papilar<br />
La dermis papilar se extiende bajo la epidermis y alrededor de las faneras. Se trata de un tejido conjuntivo laxo rico<br />
en sustancia fundamental, con ácido hialurónico y condroitín sulfato. Presenta además fibrillas colágenas de tipo I y<br />
III.<br />
Encontramos en la dermis papilar un <strong>com</strong>plejo fibrilar elástico, con fibras de oxitalán y elaunina, además de fibras<br />
elásticas que se disponen en un reticulado irregular. La función de este <strong>com</strong>plejo fibrilar elástico es facilitar el anclaje<br />
dermoepidérmico porque conforman las papilas dérmicas, donde hay gran concentración de vasos y terminaciones<br />
nerviosas.<br />
Dermis Reticular<br />
La dermis reticular es una capa de tejido conjuntivo denso, con fibras de colágeno I formando haces gruesos. Estos<br />
haces son responsables de la fortaleza mecánica de la dermis y de que la piel pueda someterse a un proceso de<br />
curtido; este estrés mecánico se evidencia en las líneas de Langer. Presenta también fibras elásticas que dotan a la<br />
dermis de elasticidad y flexibilidad. Encontramos, además de las fibras, células conjuntivas y del sistema<br />
inmunológico.<br />
El límite inferior de la dermis reticular es difuso. Según se desciende, aumenta la cantidad de adipocitos hasta llegar<br />
a la hipodermis, donde se observa una lobulación del tejido epitelial. La proporción dermis/hipodermis varía en<br />
función del sexo.
Página94<br />
La dermis reticular se atrofia con la edad: pierde sustancia fundamental, lo que eventualmente conduce a su<br />
deshidratación, presenta una mayor rigidez de las fibras colágenas, y una pérdida de fibras elásticas y de densidad<br />
celular.<br />
En la dermis reticular hay miocitos lisos dispersos entre los haces colágenos, a veces formando haces no muy<br />
gruesos, <strong>com</strong>o en el caso del músculo horripilador. Estos miocitos lisos abundan en tejidos eréctiles, <strong>com</strong>o los<br />
genitales externos y aréola mamaria.<br />
Debajo de esta capa se sitúa el panículo adiposo, de espesor variable; esta capa junto con el tejido conectivo laxo<br />
conforma la hipodermis o tejido subcutáneo.<br />
Por debajo del panículo adiposo en muchos animales hay una capa de tejido muscular estriado que conforma el<br />
panículo carnoso. En el cuello, la cara y los párpados encontramos miocitos esqueléticos, formando los músculos<br />
faciales (mímicos o de la expresión).<br />
Vascularización<br />
La dermis está vascularizada por dos plexos, uno profundo, que discurre por la dermis reticular, y otro superficial,<br />
que discurre por la dermis papilar. Del plexo profundo salen otros de menor calibre asociados a los anejos. Del plexo<br />
superficial salen asas capilares que se introducen en las papilas dérmicas.<br />
Existen en la dermis una serie de anastomosis arteriovenosas con función termorreguladora a nivel local: el plexo<br />
superficial se cierra con el frio, minimizando la pérdida de calor, y viceversa<br />
En la dermis reticular de lechos ungeales, las almohadillas plantares y en el cutis facial encontramos las estructuras<br />
vasculares conocidas <strong>com</strong>o glomus. Son anastomosis arteriovenosas ricamente inervadas, con un canal arterial (canal<br />
de Sucquer-Hoyer), donde encontramos células glómicas: miocitos lisos dispuestos a modo de esfínter.<br />
Los vasos linfáticos de la dermis son detectables en estimulación de los mismos (por ejemplo, durante una urticaria<br />
en un proceso alérgico). Recogen el líquido intersticial y permiten la migración de las células de Langerhans.<br />
Dermatoglifos<br />
La piel, en su superficie, presenta surcos, pliegues, depresiones, líneas de flexión… que conforman las arrugas<br />
epidérmicas. Estas arrugas son características de cada individuo y tienen un fundamento genético, de modo que<br />
pueden presentar patrones alterados en algunos trastornos genéticos. Además, algunas de estas arrugas, <strong>com</strong>o las<br />
huellas dactilares, tienen valor diagnóstico en medicina legal.<br />
La irregularidad de la superficie cutánea es un reflejo de la unión dermoepidérmica, porque las crestas epidérmicas<br />
tienden a disponerse en paralelo con una papila dérmica entre ellas forman estas irregularidades superficiales que se<br />
conocen <strong>com</strong>o dermatoglifos.<br />
Los dermatoglifos son mayores en zonas <strong>com</strong>o las palmas de manos y pies, por lo que la superficie de estas zonas es<br />
mayor que en aquellas zonas donde hay menor irregularidad. Esta mayor irregularidad implica que haya mayor<br />
cantidad de crestas epidérmicas y papilas dérmicas, por lo que la el estrato basal tiene una gran dimensión en estas<br />
zonas, que coinciden con zonas de piel gruesa. Se cree, que debido a esta gran extensión de superficie derivada de<br />
las uniones dermoepidérmicas, hay una gran actividad mitótica que significa una gran cantidad de estrato córneo<br />
que justifica la estructura de la piel gruesa.
Página95<br />
PELO<br />
El pelo es una excreción epidérmica anclada en la dermis o hipodermis que tiene un tallo libre rodeado por capas de<br />
vainas cilíndricas de células. En el cuero cabelludo hay unos 100000 pelos, pero en el total del cuerpo existen unos 5<br />
millones de pelos.<br />
Folículo Piloso<br />
Esquema de un pelo<br />
Los folículos pilosos permiten el desarrollo y crecimiento del pelo y <strong>com</strong>ienzan a formarse a partir del tercer mes de<br />
desarrollo.<br />
Con la pubertad, parte de las células donde había vello <strong>com</strong>ienzan a formar pelo terminal, <strong>com</strong>o en las axilas, región<br />
anogenital, bigote, barba y extremidades (en varones).<br />
Aparecen por todo el cuerpo excepto palmas, plantas, lados de los dedos, superficie lateral de los pies (bajo el<br />
tobillo), labios, glande, prepucio, clítoris, labios menores, superficie interna de los labios mayores.<br />
El folículo tiene un diámetro prácticamente uniforme excepto en la raíz (parte más inferior del folículo piloso), donde<br />
se expande formando el bulbo. La base del bulbo está invaginada por la entrada de un manguito de tejido conectivo<br />
laxo vascularizado llamado papila dérmica. En el bulbo se ubica la matriz, una capa basal de células epiteliales en<br />
mitosis constante estimulada por sustancias liberadas por los vasos de la papila dérmica.<br />
Entre las células madre encontramos melanocitos con gran cantidad de dendritas.<br />
Las células de la matriz originan la vaina radicular interna y el tallo, pero no lo hacen de manera homogénea: las<br />
células de la zona central originan el tallo y las más periféricas originan la vaina radicular interna.<br />
Cuando las células de la matriz <strong>com</strong>ienzan a ascender, pasan a una zona de transición, la zona queratógena, donde se<br />
produce la transferencia de los melanosomas a las futuras células del tallo.<br />
La raíz del folículo piloso se divide en infundíbulo, istmo, y segmento inferior (con el bulbo en la base):<br />
· Infundíbulo: desde la superficie hasta la desembocadura del conducto de la glándula sebácea.<br />
· Istmo: desde la desembocadura del conducto de la glándula sebácea hasta la inserción del músculo erector.<br />
· Segmento inferior: desde la inserción del músculo erector hasta el final.<br />
El infundíbulo y el istmo son siempre permanentes, mientras que el segmento inferior sufre cambios cíclicos. La<br />
secreción de la glándula sebácea sale por el infundíbulo, que es por tanto vía conductora de la secreción sebácea.
Página96<br />
Histológicamente, la raíz presenta cuatro capas, desde la parte más externa hasta el tallo: la vaina conjuntiva, la<br />
membrana vítrea, la vaina radicular externa y la vaina radicular interna.<br />
Vaina Conjuntiva<br />
Esta vaina es tejido conjuntivo que rodea periféricamente a la raíz. Se divide en una capa más externa, con fibras<br />
elásticas y colágenas orientadas longitudinalmente; y una capa interna, con fibras colágenas en disposición<br />
perpendicular. Entre ambas capas se localiza el plexo nervioso peritriquial: fibras aferentes que recogen el cambio de<br />
orientación del tallo.<br />
La membrana vítrea no es otra cosa que la lámina basal que separa esta vaina conjuntiva de las siguientes capas de<br />
carácter epitelial.<br />
Vaina Radicular Externa<br />
Es un epitelio que se halla en continuidad con la epidermis. Las células están queratinzadas con queratina blanda (no<br />
dura, <strong>com</strong>o la del tallo). En la zona del istmo se estrecha y pierde los estratos córneo y granuloso.<br />
Profundiza, se adelgaza y se continúa con una región que se denomina matriz. En la zona del infundíbulo, esta vaina<br />
no está en contacto con el tallo, sino que media una separación. En el istmo y el segmento inferior, encontramos la<br />
vaina radicular interna a continuación de la externa.<br />
En la zona final del istmo, forma un engrosamiento que recibe el nombre de rodete.<br />
En él se localizan células madre que permiten renovar la epidermis: van a la capa basal, descienden al bulbo<br />
renovando la matriz y renuevan las glándulas sebáceas.<br />
Vaina Radicular Interna<br />
Es un epitelio que no está en continuidad con epidermis, sino que deriva de la matriz y se encuentra rodeando el<br />
tallo en el istmo y el segmento inferior. Por encima de éstos, se desintegra y se pierde. Sus células presentan<br />
queratina blanda.<br />
Histológicamente, se distinguen en la vaina tres capas, de más externa a más interna:<br />
Capa de Henle: epitelio simple cúbico bajo o plano.<br />
Capa de Huxley: epitelio estratificado plano. Presenta gránulos eosinófilos de tricohialina.<br />
Cutícula de la vaina radicular interna: epitelio estratificado plano queratinizado, en contacto con la cutícula<br />
del tallo, fijando el tallo a la raíz.<br />
Las capas se fusionan en el infundíbulo y acaban descamándose <strong>com</strong>o una masa eosinófila.<br />
Tallo<br />
El pelo o tallo es un filamento de longitud muy variable (desde 2 mm. hasta más de 1 m) y grosor variable. Sale de la<br />
superficie de la piel con orientación oblicua. Está queratinizado, con una queratina más dura que en epidermis, con<br />
residuos de azufre, de manera que las células no se pueden descamar.<br />
El tallo se <strong>com</strong>pone de tres capas: cutícula, corteza y médula.<br />
La cutícula es epitelio plano estratificado queratinizado, con células dispuestas <strong>com</strong>o tejas (imbricadas), de modo<br />
que es muy impermeable.<br />
La corteza es un epitelio cúbico queratinizado con melanosomas en número variable, que proceden de los<br />
melanocitos alojados en el bulbo.<br />
La médula es un epitelio queratinizado con espacios de aire entre medias. Estos espacios son la causa de que el pelo<br />
se vuelva cano, ya que con la edad aumentan los espacios.<br />
El vello es un pelo más fino, blando y corto. No tiene médula ni pigmento, y se introduce hasta la dermis. Cubre el<br />
tronco y los miembros.<br />
El pelo terminal es más grueso y firme, crece mucho, tiene médula y está pigmentado. Se aloja en la hipodermis y se<br />
encuentra en el cuero cabelludo, pestañas, cejas, etc.
Página97<br />
El color del pelo terminal es muy variable y depende de dos factores: la cantidad de melanina y la distribución de los<br />
melanosomas: el pelo cano tiene melanocitos escasos y melanina, el pelo rubio es escaso en melanosomas pero<br />
normal melanocitos y el pelo rojo presenta gran cantidad de feomelanina.<br />
Crecimiento del Pelo<br />
El crecimiento del pelo es dependiente de la matriz, de modo que el tallo crece en longitud, paro lo cual la vaina<br />
radicular desaparece a la altura del infundíbulo. Es un crecimiento cíclico, con tres fases:<br />
1. Fase anagénica (hasta 8 años): el folículo está creciendo y el tallo también (de manera más activa en<br />
mujeres). El segmento inferior es muy profundo y alcanza la hipodermis. LA papila dérmica muy bien<br />
formada y la matriz es muy activa. Supone el 85-90% del proceso.<br />
2. Fase catagénica (2 ó 3 semanas): se produce un cese en el crecimiento y el folículo involuciona. Se observa<br />
una retracción del segmento inferior, atrofia del bulbo y reducción drástica de la actividad mitótica en la<br />
matriz y de la síntesis de melanina en melanocitos. Supone un 10-15% del proceso.<br />
3. Fase telogénica (3 ó 4 meses): el segmento inferior es muy corto (
Página98<br />
GLÁNDULAS SEBÁCEAS<br />
Las glándulas sebáceas son glándulas holocrinas alveolares ramificadas que se originan <strong>com</strong>o brotes de la vaina<br />
radicular externa del folículo piloso. La secreción es de un sebo que se libera cuando la célula secretora muere al<br />
llenarse <strong>com</strong>pletamente por el producto, de modo que la secreción y los restos celulares pasan al canal pilosebáceo,<br />
un canal corto y ancho de epitelio estratificado plano.<br />
Se asocian casi siempre a un folículo piloso. Pueden estar a ambos lados del folículo (generalmente hay más de una<br />
por folículo), pero suelen encontrarse en el ángulo obtuso del tallo. Son particularmente abundantes en la piel<br />
seborreica.<br />
En algunas zonas no se asocian a folículos, <strong>com</strong>o:<br />
Glándulas de Fordyce: en la <strong>com</strong>isura de los labios.<br />
Glándulas de Montgomery: en la areola mamaria para suavizar la zona de succión.<br />
Glándulas en el área genital (glándulas de labios menores en la mujer y glándulas de Tyson en glande).<br />
Glándulas de Meibonio o tarsales, en los párpados.<br />
Presentan células basales aplanadas, ricas en orgánulos, de activa división. Están unidas por desmosomas y su lámina<br />
basal es continua con la epidermis y el folículo piloso. Las células basales contienen REL y RER, ribosomas libres,<br />
mitocondrias, glucógeno y escasas inclusiones lipídicas, pero según ascienden se desarrollan REL y crestas tubulares,<br />
que permiten que la célula se vaya cargando de lípidos. Estas inclusiones lipídicas son pequeñas gotitas separadas<br />
por delgados tabiques de citoplasma. Finalmente entran en apoptosis, es decir, siguen un mecanismo de secreción<br />
holocrina. La célula tarda 8 días en realizar este ciclo y morir.<br />
La secreción de estas glándulas es el sebo y presentan una secreción continua. El sebo es conducido por un conducto<br />
excretor de epitelio estratificado plano, en continuación con la vaina radicular externa. El sebo es lubricante,<br />
emoliente, protege frente a las temperaturas extremas y tiene propiedades bacteriostáticas.<br />
Las glándulas son activas desde el nacimiento, mucho más en la pubertad. Las hormonas sexuales masculinas<br />
estimulan su actividad, mientras que las femeninas la inhiben.<br />
GLÁNDULAS SUDORÍPARAS<br />
Glándulas Sudoríparas Ecrinas<br />
Las glándulas sudoríparas ecrinas aparecen a lo largo de toda la superficie de la piel, excepto en los labios y genitales<br />
externos.<br />
Son activas desde el nacimiento, no se asocian al folículo piloso, son más pequeñas que las apocrinas, su estructura<br />
es tubular enrollada simple y su conducto excretor se denomina segmento canalicular.<br />
Se encuentran en la dermis profunda o en la hipodermis. Son epitelios pseudoestratificados, con células secretoras<br />
claras y oscuras y células miopiteliales.<br />
El sudor ecrino es pobre en proteínas, hipotónico, con sales <strong>com</strong>o cloruro sódico, potasio y amonio (se pierden<br />
electrolitos con él), así <strong>com</strong>o lactato y urea. Se pueden secretar hasta 10 de sudor ecrino al día.<br />
CÉLULAS CLARAS, OSCURAS Y MIOEPITELIALES<br />
Las células claras son más pequeñas, se apoyan basalmente y no alcanzan la luz demasiado. Presentan pliegues,<br />
almacenan glucógeno (son PAS+) y muchas mitocondrias. Secretan el <strong>com</strong>ponente acuoso del sudor ecrino, que sale<br />
por la membrana lateral, de manera que alcanza un espacio llamado canalículo o capilar secretor.<br />
Las células oscuras ocupan gran cantidad de la superficie luminal. Sintetizan glicoproteínas, que se almacenan en el<br />
citoplasma apical, por lo que presentan un gran desarrollo del RER y del aparato de Golgi.
Página99<br />
Las células mioepiteliales facilitan la salida del sudor; presentan gran cantidad de filamentos de actina que al ser tan<br />
eosinófilos permiten ser fácilmente identificados.<br />
Segmento Canalicular<br />
Es el nombre que recibe el conducto secretor de la glándula sudorípara ecrina.<br />
Presenta un enrollamiento no muy tortuoso y una luz estrecha, tapizada por epitelio estratificado cúbico formado<br />
por una capa basal de células y una capa de células luminales, es decir, es predominantemente biestratificado.<br />
Permite drenar el sudor ecrino hasta alcanzar la epidermis, donde desaparece el conducto y aparece un túnel a lo<br />
ancho de la epidermis para secretar el sudor.<br />
En este segmento las células son basales (núcleo redondo, nucleolo, mitocondrias) o luminales (tonofilamentos,<br />
citoplasma vítreo). En el segmento canalicular no encontramos células mioepiteliales.<br />
El segmento canalicular permite la salida del sudor y modifica su <strong>com</strong>posición: se reabsorbe agua y sodio, función<br />
que es controlada por la aldosterona. El sudor es isotónico cuándo es sintetizado y acaba siendo hipotónico al<br />
secretarse.<br />
Las glándulas ecrinas responden a estímulos térmicos. Un incremento de la temperatura hace que las células sean<br />
más activas, de modo que la evaporación del sudor refrigera el cuerpo. Las glándulas ecrinas se activan primero en la<br />
frente y el cuero cabelludo, luego en la cara y el resto del cuerpo y finalmente en la piel glabra.<br />
Presentan un control de tipo colinérgico: se incrementa la perfusión en la zona a refrigerar, evitando el<br />
sobrecalentamiento.<br />
Responden asimismo a estímulos emocionales. El control de estos estímulos es de tipo adrenérgico, de modo que<br />
responden primero la piel glabra y las axilas.<br />
Glándula Sudorípara Apocrina<br />
La glándula sudorípara apocrina proviene del folículo piloso, permanediendo unidas a él, y aparecen en las axilas,<br />
areolas, pezones de las glándulas mamarias, región perineal y genitales externos. Las glándulas de Moll y<br />
ceruminosas tienen su misma estructura.<br />
Son glándulas de gran tamaño (5 mm), de tipo tubular enrollada, a veces ramificada que drenan a un conducto<br />
excretor que se abre al infundíbulo, por encima del conducto sebáceo. Se activan en la pubertad.<br />
Los adenómeros presentan una luz muy dilatada, delimitada por epitelio simple cúbico o cúbico bajo formado por un<br />
único tipo celular de carácter eosinófilo y generalmente con una protrusión en forma de ampolla en su superficie<br />
apical que contiene los gránulos de excreción.<br />
El adenómero sintetiza y segrega el sudor, además de almacenarlo temporalmente. Es un sudor lechoso, viscoso, de<br />
color blanquecino, inodoro, rico en proteínas, azúcares, lípidos, hierro, y lipofucsina.<br />
Al microscopio óptico parece que el citoplasma se pierde en la secreción, pero al electrónico se ve que no es así. En<br />
realidad es una secreción de tipo merocrina.<br />
Hay además un grupo de células mioepiteliales facilitan la secreción del sudor; se localizan en la porción secretora de<br />
la glándula entre las células secretoras y junto a la lámina basal de estas.<br />
El conducto está tapizado por un epitelio cúbico estratificado, de dos a tres células de espesor. Tiene una luz angosta<br />
y desemboca en el conducto folicular sin alterar la secreción de las células secretoras, por lo que su función es<br />
exclusivamente conductora.<br />
El sudor tiene sustancias bacteriostáticas y bactericidas. Es transformado por las bacterias, pudiendo adquirir<br />
sustancias odoríferas, actuando <strong>com</strong>o feromonas (que participan en la percepción de sexos).<br />
Responden a estímulos emocionales y sensitivos, pero no térmicos. Es un control de tipo adrenérgico.
Página100<br />
Glándula Sudorípara Apoecrina<br />
Mezcla de las glándulas apocrina y ecrina porque la secreción es ecrina mediante una secreción tipo apocrina.<br />
Aparece en los pliegues cutáneos (se asocia a hiperhidrosis) y aparecen en la adolescencia.<br />
UÑA<br />
Las uñas o placas ungueales son placas córneas, curvas, situadas en la cara dorsal de las falanges terminales de los<br />
dedos. Se asientan sobre una epidermis que se denomina lecho ungueal, en la cual solo hay estratos basal y<br />
espinoso. Se caracteriza porque las papilas dérmicas son grandes y muy vascularizadas, menos en la zona de la<br />
luneta.<br />
Las uñas se separan del resto de la epidermis por el surco ungueal, y presentan una elevación llamada rodete<br />
ungueal.<br />
La placa ungueal es transparente, con escamas córneas densamente apretadas. Son placas de queratina dura, de<br />
modo que no se descama; en la formación de esta K dura no intervienen gránulos de queratohialina. La queratina<br />
dura se <strong>com</strong>pone de filamentos intermedios de K densamente agrupada con una matriz de K amorfa muy rica en<br />
azufre. Presenta una raíz, proximalmente, y una lúnula, distalmente.<br />
En la raíz encontramos una matriz de células madre en continua división, que salen de la matriz para diferenciarse y<br />
adquieren queratina dura. La placa ungueal se desliza, sin contribución del lecho.<br />
La lúnula tiene células solo parcialmente queratinizadas y tiene forma de semiluna blanquecina.<br />
En la raíz, la uña está cubierta por el eponiquio o cutícula. Si se pierde el eponiquio, la matriz puede inflamarse e<br />
infectarse.<br />
Bajo la uña hay un reborde epidérmico llamado hiponiquio, que asegura la impermeabilidad del lecho. Si se rompe,<br />
se dan infecciones bacterianas y fúngicas.<br />
Esquema de una uña
Página101<br />
SISTEMA ENDOCRINO<br />
El sistema endocrino juega, junto con el sistema nervioso, un importante papel en la <strong>com</strong>unicación intercelular. Su<br />
acción es más lenta y prolongada que la acción del sistema nervioso, y está regulada por la liberación de hormonas.<br />
Las hormonas tienen 4 mecanismos principales de acción:<br />
1. Hormonas liposolubles: entran en la célula diana y producen cambios en ella.<br />
2. Proteínas: se unen a proteínas específicas de la membrana celular de la célula diana.<br />
3. Variación del voltaje de membrana ( ); en células musculares y nerviosas.<br />
4. Retroalimentación negativa: el producto inhibe la posterior secreción.<br />
El sistema endocrino está formado por células secretoras llamadas células endocrinas que se organizan<br />
anatómicamente <strong>com</strong>o órganos endocrinos (hipófisis, tiroides, paratiroides, suprarrenales y epífisis), se agrupan<br />
dentro del órgano (islotes de Langerhans, células intersticiales de Leydig y células endocrinas ováricas) o dispersas y<br />
aisladas entre otras células en tejidos epiteliales (<strong>com</strong>o el sistema neuroendocrino difuso o APUD). Estas células,<br />
aunque se encuentren dispersas en otros tejidos, no deben subestimarse, pues suponen el mayor órgano endocrino<br />
del organismo, con su función endocrina y paracrina sobre las células epiteliales vecinas.<br />
Las células endocrinas se disponen en los órganos <strong>com</strong>o nidos o cordones muy vascularizados para verter<br />
eficazmente su secreción al torrente sanguíneo.<br />
El origen de las glándulas es epitelial, excepto el <strong>com</strong>ponente neuronal de la hipófisis.<br />
Tipos de hormonas<br />
<br />
<br />
<br />
<br />
Esteroides: son derivados del colesterol, <strong>com</strong>o la progesterona, el cortisol, el estradiol y la testosterona.<br />
Pequeños péptidos: encefalina, vasopresina y hormona liberadora de la hormona tiroidea.<br />
Proteínas y Glucoproteínas: Factor de Crecimiento Nervioso, F.C. Epidérmico, paratohormona y Hormona<br />
Estimuladora de la tiroides.<br />
Derivados de aa: tiroxina y catecolaminas <strong>com</strong>o adrenalina y noradrenalina.<br />
Otros autores (Ross, 4ª ed.) incluyen en un solo grupo a los péptidos pequeños, las P y las glucoP.<br />
HIPÓFISIS<br />
La hipófisis es un órgano macizo que junto con el<br />
hipotálamo conforma lo que se conoce <strong>com</strong>o el<br />
diencéfalo.<br />
Macroscópicamente se define <strong>com</strong>o una glándula<br />
endocrina <strong>com</strong>puesta del tamaño de un guisante<br />
(10x3x6mm y un peso de 0,5g en varones normales y<br />
1,5g en mujeres multíparas) que se localiza en la silla<br />
turca del hueso esfenoides.<br />
No es independiente del encéfalo debido a que se une a<br />
este mediante un pequeño tallo llamado infundíbulo que<br />
lo fija al hipotálamo.
Página102<br />
Se pueden distinguir dos grandes zonas funcionales:<br />
a) Adenohipófisis o hipófisis A, tejido epitelial glandular.<br />
b) Neurohipófisis o hipófisis P, tejido secretor neural.<br />
Embriología de la Hipófisis<br />
La adenohipófisis deriva de una evaginación del techo del estomodeo (ectodermo de orofaringe): la bolsa de Rathke.<br />
La neurohipófisis deriva de una evaginación del suelo del diencéfalo: el infundíbulo (neuroectodérmica).<br />
Cuando las dos evaginaciones contactan, la conexión con el estomodeo se pierde pero la del infundíbulo con el<br />
diencéfalo persiste y se convertirá en el tallo neural. La porción más inferior prolifera y se convierte en pars nervosa<br />
Se desarrollan fibras nerviosas hacia el<br />
tallo neural y algunas hacia la pars<br />
nervosa. La porción de la bolsa de Rathke<br />
en contacto con la pars nervosa se<br />
convertirá en la pars intermedia. El resto<br />
crece mucho y se convierte en la pars<br />
distalis, de la que salen dos lengüetas que<br />
van creciendo alrededor del tallo neural y<br />
finalmente se unen formando una<br />
envuelta, la pars tuberalis.<br />
Pueden quedar restos de la luz de la bolsa<br />
de Rathke en forma de pequeñas vesículas<br />
con coloide junto a la neurohipófisis.<br />
También puede haber restos de la <strong>com</strong>unicación de la bolsa con el estomodeo, lo que se denomina hipófisis faríngea.<br />
Irrigación<br />
La irrigación sanguínea proviene de dos grupos de vasos:<br />
I. Arterias Hipofisiarias Superiores: irrigan a la pars tubelaris, la eminencia media y el tallo infundibular. La sangre<br />
viene de las arterias carótidas internas y de la <strong>com</strong>unicante P del polígono de Willis.<br />
II. Arterias Hipofisiarias Inferiores: irrigan fundamentalmente la pars nervosa y su sangre proviene<br />
exclusivamente de la arteria carótida interna.<br />
Sistema Portal Hipofisiario<br />
La mayor parte de la adenohipófisis no tiene irrigación arterial directa.<br />
Las arterias que irrigan la pars tubelaris, la eminencia media y el tallo infundibular dan origen a capilares fenestrados<br />
(el plexo capilar primario) que drenan en las venas porta hipofisiarias que transcurren a lo largo de la pars tubelaris<br />
hasta dar origen a un plexo capilar secundario de sinusoides fenestrados.<br />
Este Sistema Porta Hipofisiario transporta las secreciones neuroendocrinas de los nervios hipotálamicos desde sus<br />
sitios de liberación en la eminencia media y el tallo infundibular hasta las células de la pars distalis.<br />
Hay otro plexo capilar terciario en la pars nervosa que procede de la arteria hipofisaria inferior. Recoge las<br />
secreciones de la neurohipófisis.<br />
Inervación<br />
La hipófisis está penetrada por los nn. que ingresan en el tallo infundibular y en la pars nervosa desde los núcleos<br />
hipotalámicos, que en conjunto forman parte de la neurohipófisis.<br />
Además, hay fibras posganglionares del sistema nervioso autónomo con función vasomotora que se introducen en la<br />
adenohipófisis.
Página103<br />
Adenohipófisis<br />
La adenohipófisis costa de una cápsula y tabiques poco desarrollados que sostienen a un parénquima formado por<br />
células endocrinas que se organizan en grupos y cordones separados por capilares fenestrados de diámetro muy<br />
grande.<br />
Las células de la adenohipófisis sintetizan y secretan hormonas tróficas <strong>com</strong>o respuesta a estímulos de la<br />
neurohipófisis para regular la actividad de las células de otras glándulas y tejidos endocrinos.<br />
Está formada por 3 derivados de la bolsa de Rathke:<br />
Pars distalis: representa la mayor parte de la adenohipófisis y resulta del engrosamiento de la pared A de la<br />
bolsa de Rathke.<br />
Pars intermedia: delgado remanente de la pared P y que termina en el infundíbulo.<br />
Pars tuberalis: collar o vaina alrededor de la pars intermedia que se origina por engrosamiento de las<br />
paredes laterales de la bolsa de Rathke.<br />
PARS DISTALIS<br />
Parénquima<br />
Corresponde al 75% de la glándula e inicialmente sus células fueron descritas según sus características tintoriales de<br />
los gránulos de secreción. Así resultaron 3 tipos celulares:<br />
1. Basófilas (10%)<br />
2. Acidófilas (40%) CROMÓFILAS<br />
3. Cromófobas (50%): ha liberado sus gránulos y contienen lisosomas secundarios y cuerpos enigmáticos<br />
(vacuolas perinucleares).<br />
De todos modos, hoy se usan otros métodos para conocer los distintos tipos celulares contenidos en esta zona:<br />
HISTOQUÍMICOS<br />
El uso de tinciones específicas <strong>com</strong>o la tinción del ácido peryódico de Schiff o PAS, que revela dos tipos celulares:<br />
PAS+: producen hormonas glucoproteicas; son más escasas y basófilas (mucosas).<br />
PAS-: producen hormonas proteináceas; son más abundantes y acidófilas o cromófobas (serosas).<br />
También se usan las tinciones tricrómicas (Mallory, Cleveland-Wolfe,…) y otras específicas que permiten dividir a las<br />
células según sus funciones.<br />
ESTUDIOS HISTOFISIOLÓGICOS<br />
Identificación de las funciones celulares, y en consecuencia de sus tipos, mediante la <strong>com</strong>binación de tinciones y<br />
manipulación de cantidad o tamaño en órganos blanco de las hormonas tróficas. Así, algunas células que se creían<br />
cromófobas estaban en período de desgranulamiento tras una secreción.<br />
MICROSCOPÍA ELECTRÓNICA E INMUNOHISTOQUÍMICA<br />
Aporta más datos para diferenciación celular.<br />
Junto a las células glandulares aparecen las células foliculoestrelladas, de naturaleza epitelial con largas<br />
prolongaciones que permiten dar sostén a la glándula. Estas células pueden delimitar pequeñas cavidades junto a la<br />
pars nervosa, por lo que se habla también de folículos. No se ha definido ninguna función concreta.
Página104<br />
CÉLULAS ACIDÓFILAS<br />
Las células acidófilas son las segundas más numerosas (40%). Tienden a ubicarse en las regiones más laterales de la<br />
pars distalis. No son muy grandes, son redondeadas y son característicos los gránulos eosinófilos. Encontramos dos<br />
tipos:<br />
<br />
<br />
Células somatotróficas: son el 50% de todas las hipofisarias. Se sitúan en las zonas laterales y tienen<br />
abundante RER y aparato de Golgi. Tienen gránulos de secreción bastante densos, de unos 300-350nm.<br />
Son estimuladas por la GHRH, pero también pueden ser inhibidas por la somatostatina o cuando los niveles<br />
de glucemia son elevados. Secretan GH.<br />
Células lactotróficas o mamotróficas: forman el 15-20% del total celular hipofisario. No tienden a ubicarse en<br />
ningunas situación concreta, sino que son uniformes por toda la pars distalis. Su porcentaje aumenta en las<br />
mujeres gestantes y en la lactancia. En esos periodos aumenta también su tamaño y su dotación de<br />
orgánulos (RER, Golgi y gránulos). Los gránulos son muy densos y tienen un tamaño mayor que los de las<br />
somatotróficas, de 600-900nm. Los orgánulos que aumentan, tras la lactancia se eliminan por autofagia.<br />
Su principal regulador es la dopamina, que es inhibidora; la prolactina estimula la dopamina (retroinhibición).<br />
También son estimuladas por PRH y TRH. Secretan prolactina.<br />
CÉLULAS BASÓFILAS<br />
Las células basófilas son muy minoritarias (10%). Su tamaño es redondeado y mayor, y sus gránulos son ligeramente<br />
basófilos tiñéndose claramente con PAS. Hay tres tipos:<br />
<br />
<br />
<br />
Células tirotróficas: producen TSH (hormona estimulante del tiroides), tirotrofina o tirotropina. Del total de<br />
células de la pars distalis, las tirotróficas son el 5%. Tienden a situarse en la porción más anterior. Tienen<br />
morfología muy variable y orgánulos abundantes. Los gránulos son muy densos pero bastante pequeños, de<br />
unos 100-150nm.<br />
Responden a la TRH. Son inhibidas cuando la T 3 y la T 4 se segregan de forma activa (retroinhibición).<br />
Células gonadotróficas: producen FSH (hormona estimulante de folículos) y LH (hormona luteinizante). Son el<br />
10% del total. Se distribuyen de forma uniforme. Sintetizan dos hormonas, FSH y LH, que se empaquetan<br />
juntas en las mismas vesículas. Los gránulos son pequeños, de 200-250nm.<br />
La liberación de la hormona corre a cargo de la GnRH que tiene secreción pulsátil cada 90 minutos. La FSH<br />
está estimulada por la activina, sintetizada tanto por las células de Sertoli <strong>com</strong>o por las foliculares. La<br />
liberación tanto de FSH <strong>com</strong>o de GnRH es inhibida por la inhibina. Las propias células de la pars distalis<br />
secretan inhibina, así <strong>com</strong>o las foliculares y las de Sertoli. También son inhibidas por el estradiol.<br />
Células corticotróficas: producen ACTH (hormona corticotrofa) o corticotrofina. Son el 20% del total y se<br />
sitúan en la línea media. Pueden presentar prolongaciones. Poseen un citoplasma bastante claro, un núcleo<br />
excéntrico y muchas mitocondrias. El RER forma cisternas cortas. Los gránulos de secreción son pocos y<br />
pequeños (200-250nm) y se sitúan en la periferia.<br />
Estas células se regulan por la CRH, sintetizada en los núcleos paraventriculares del hipotálamo. Responden a<br />
situaciones de estrés y son estimuladas por la ADH e inhibidas por el cortisol.<br />
Sintetizan POMC <strong>com</strong>o precursor, que no es exclusivo de estas células, pues también se encuentra en el<br />
hipotálamo, hígado, corazón, riñones y tejidos digestivos (puede que también en los reproductores). Se han<br />
formulado varias hipótesis.<br />
Estas células sintetizan ACTH y la hormona β-lipotrófica (β-LPH).
Página105<br />
Hormonas<br />
Todas las hormonas adenohipofisiarias se definen <strong>com</strong>o proteínas pequeñas o glucoproteínas que suelen ser<br />
producidas por un tipo celular, excepto en las células adrenocorticolipotrofas que producen 2 (ACTH y LPH) y las<br />
gonadotrofas (LH y FSH).<br />
La GH (hormona del crecimiento) o somatotrofina es una hormona que se libera de forma pulsátil (pico al inicio del<br />
sueño). Estimula los hepatocitos, que segregan IGF1 o somatomedina C. El IGF1 actúa sobre la zona de reserva de<br />
cartílago metafisario, por lo que actúa directamente en el crecimiento en longitud de los huesos largos. También<br />
estimula la diferenciación de las células satélites, aumentando el desarrollo de la musculatura esquelética.<br />
Como patologías encontramos:<br />
Adenomas: tumor benigno que produce un exceso de GH.<br />
Gigantismo: tumor secretor de GH en la infancia o pubertad que hace que los cartílagos epifisarios crezcan en<br />
exceso.<br />
Enanismo hipofisario: déficit de GH en niño. Se trata administrando GH humana sintetizada mediante<br />
tecnología genética.<br />
La prolactina es una hormona de secreción pulsátil. Hay un pico cada vez que el bebé succiona. Es importante en<br />
todo el periodo de lactancia (mamogénesis, lactogénesis y galactopoyesis).<br />
La TSH o tirotropina se encarga de controlar la función del tiroides y el crecimiento. Hay una conexión con las<br />
lactotróficas ya que estimula la síntesis y secreción de prolactina<br />
Un déficit en la secreción de TSH produce hipotiroidismo, con reducción del metabolismo celular, temperatura y<br />
metabolismo basal. También produce lentitud mental.<br />
La FSH (hormona foliculoestimulante) estimula el desarrollo de los folículos ováricos. Es responsable de que haya<br />
más activina, inhibina y estradiol. También estimula la producción de estrógenos y del ABP en las células de Sertoli.<br />
La LH (hormona luteinizante) estimula la maduración folicular, la ovulación y la formación del cuerpo lúteo. También<br />
estimula la estereidogénesis (progesterona) en el folículo ovárico y el cuerpo lúteo y, en los hombres, la síntesis de<br />
testosterona en las células de Leydig (se conoce <strong>com</strong>o ICSH).<br />
Un déficit en FSH y LH lleva a la reducción de la fertilidad. En las mujeres provoca trastornos menstruales y en los<br />
hombres hipogonadismo hipogonadotrópico (testículos pequeños e infertilidad). Cuando se produce la extirpación de<br />
testículos (orquiectomía o castración) u ovarios (ooferectomía) aumentan los niveles de FSH y LH. Las células<br />
gonadotróficas se hipertrofian y muestran un aspecto vacuolado pasando a conocerse <strong>com</strong>o células de la castración.<br />
La β-LPH (hormona lipotrófica) rinde γ-LPH y β-endorfina. Tanto la β-LPH <strong>com</strong>o la γ-LPH tienen carácter lipolítico. La<br />
γ-LPH contiene en su secuencia la β-MSH (hormona estimuladora de melanocitos), pero esta última no se secreta en<br />
el hombre.<br />
La ACTH (hormona adrenocorticotrófica) es una molécula que se segrega de forma circadiana con picos al inicio del<br />
día. Estimula células de la corteza suprarrenal, fundamentalmente de las zonas fascicular y reticular, donde se<br />
sintetizan glucocorticoides y esteroides sexuales. Estimula la lipolisis y aumenta la pigmentación de la piel.<br />
En vertebrados inferiores se puede degradar rindiendo dos moléculas, la α-MSH (hormona estimuladora de<br />
melanocitos) y CLIP (péptido intermedio parecido a corticotropina). Tanto la α-MSH <strong>com</strong>o la β-MSH regulan la<br />
dispersión de gránulos de melanina en melanóforos y son responsables de los cambios de coloración en la piel de<br />
peces, anfibios y reptiles.<br />
Como patologías asociadas encontramos la enfermedad de Cushing, que se debe a un adenoma en la adenohipófisis<br />
que aumenta la secreción de ACTH. Hay aumento de cortisol, tendencia a la obesidad, osteoporosis y atrofia<br />
muscular. También podemos encontrar déficit de ACTH, lo que provoca reducción en la secreción de andrógenos de la<br />
corteza suprarrenal. En las mujeres esto ocasiona la pérdida del vello axilar y púbico.
Página106<br />
TIPO CÉLULA<br />
TINCIÓN<br />
GENERAL<br />
TINCIÓN<br />
ESPECÍFICA<br />
PRODUCTO<br />
(hormona)<br />
TAMAÑO/<br />
FORMA<br />
OTRAS<br />
CARACTERÍSTICAS<br />
Somatotrofas<br />
(50%)<br />
Acidófilas Orange G STH<br />
Pequeñas<br />
redondas<br />
y<br />
X<br />
Mamotrofas o<br />
lactotrofas<br />
(15-20%)<br />
Acidófilas<br />
Orange G<br />
Eritrosina<br />
Carmín<br />
LTH<br />
Variable<br />
Gran cantidad lisosomas tras<br />
lactancia<br />
Adrenocorticolipotrofas<br />
(15-20%) Basófilas PAS ACTH y LPH<br />
Redondas y<br />
ovales<br />
Inclusiones lipídicas y<br />
filamentos de 6-8nm<br />
Gonadotrofas<br />
(10%)<br />
Tirotrofas (5%)<br />
Basófilas<br />
Basófilas<br />
PAS<br />
Aldehídofuscina<br />
PAS (débil)<br />
FSH y LH/ISCH<br />
TSH<br />
FSH: grandes y<br />
redondas<br />
ISCH: pequeñas y<br />
redondas<br />
Grandes y<br />
poligonales<br />
FSH: vesículas RER<br />
distendidas<br />
ISCH: RER mínimo<br />
X<br />
Tabla resumen de células Pars distalis de Adenohipófisis con sus características más importantes
Página107<br />
PARS INTERMEDIA<br />
La pars intermedia sólo aparece en la etapa fetal y en un corto periodo postnatal, pero en otras especies está bien<br />
desarrollada. Se puede encontrar rodeando una serie de cavidades quísticas que representan la luz residual de la<br />
bolsa de Rathke.<br />
Las células pueden ser basófilas o cromófobas. Estas células pueden en ocasiones invadir la pars nervosa<br />
produciendo una infiltración basófila. Estas células pueden ocasionalmente formar folículos (con coloide). Es curioso<br />
observar que en estos folículos el epitelio no es igual por toda su superficie. En la parte posterior es cúbico simple, y<br />
en A es estratificado.<br />
Estas células basófilas forman la melanocortina, que en poiquilotermos se escinde y forma la α-MSH y β-MSH. En la<br />
etapa fetal se forma α-MSH, pero en el adulto pierde funcionalidad y las células basófilas se consideran<br />
corticotróficas.<br />
PARS TUBERALIS<br />
A partir de la pars distalis se forman dos lengüetas que crecen a lo largo del tallo hipofisario. Rodean el tallo y se<br />
fusionan formando una capa celular delgada, la pars tuberalis. Es la parte más vascularizada gracias al sistema porta<br />
hipotálamo-hipofisario.<br />
Desde el punto de vista histológico tienen células basófilas organizadas en cordones longitudinales y ocasionalmente<br />
hay folículos.<br />
Su función en el hombre no es fundamentalmente secretora, aunque existen algunas células gonadotrofas<br />
funcionales y además las células de esta zona presentan receptores de melatonina.<br />
------------------------------------FACTORES DE REGULACIÓN HORMONAL------------------------------------------------------<br />
La liberación de las hormonas tróficas está regulada por el hipotálamo a través de hormonas llamadas factores<br />
reguladores hipotalámicos que son vertidos al sistema porta hipofisiario; se usan las siglas RH (GnRH = Regulador<br />
Hipotalámico de la gonadotropina) o Fr/I según activen o inhiban la liberación de una hormona).<br />
Su función es activadora (factores liberadores) o inhibidora (factores inhibidores) de la secreción y se liberan según<br />
los niveles de hormonas tróficas en plasma.<br />
Por ejemplo, si disminuye el nivel de hormonas tiroideas se secreta el factor liberador de tirotrofina (TRF) para<br />
aumentar la cantidad de hormona tiroide en sangre a través de la tirotrofina.<br />
Existen:<br />
Factor liberador de la hormona del crecimiento (GRF)<br />
Factor inhibidor de la hormona del crecimiento (GIF)<br />
Factor liberador de prolactina (PRF)<br />
Factor inhibidor de prolactina (PIF)<br />
Factor liberador de corticotrofina (CRF)<br />
Factor liberador de gonadotrofina (GnRF)<br />
Factor liberador de tirotrofina (TRF).
Página108<br />
Neurohipófisis<br />
Está formada por la eminencia media, el tallo neural (eje del tallo hipofisario), la pars nervosa del lóbulo P y los<br />
nervios hipotalámicos.<br />
La pars nervosa almacena y libera hormonas sintetizadas a nivel hipotalámico. Capilares fenestrados con mastocitos,<br />
fibroblastos y pituicitos asociados a ellos.<br />
Los pituicitos son células de tipo glial, que, por tanto, van a dar sostén. Tienen un núcleo redondo u oval con un<br />
citoplasma rico en gliofilamentos PGFA+ y gotas de lípidos. Tienen muchas prolongaciones a veces largas que se<br />
asocian a los axones y terminan en la lámina basal de los capilares. El contacto con la lámina basal endotelial no es<br />
permanente porque en el momento en el que se produce la descarga de la hormona los pituicitos se retraen para<br />
dejar espacio al proceso.<br />
Tractos nerviosos<br />
Son fibras que proceden de neuronas hipotalámicas neurosecretoras. Son neuronas que no acaban en sinapsis, sino<br />
que vierten su secreción a los capilares. Atraviesan el infundíbulo, formando el tracto hipotálamo-hipofisario, en el<br />
que hay unos 100000 axones amielínicos. Si seccionamos el tracto hipotálamo-hipofisario vemos dos tipos de fibras.<br />
Unas forman el tracto tubero-hipofisario o tubero-infundibular, que termina en la eminencia media. Otras fibras van<br />
a llegar hasta la pars nervosa y forman el tracto supraóptico-hipofisario o paraventricular-hipofisario.<br />
El tracto tubero-hipofisario está formado por axones amielínicos de neuronas situadas en los núcleos ventromedial,<br />
dorsomedial e infundibular del hipotálamo. Acaban en la eminencia media y liberan hormonas hipofisotropas que se<br />
vierten en el plexo capilar primario.<br />
Si nos situamos en la eminencia media, aparte del tracto tubero-hipofisario encontramos otras fibras que vierten al<br />
plexo primario oxitocina y vasopresina (ADH). Estas neuronas proceden de los núcleos supraóptico y paraventricular.<br />
La sangre del plexo primario está enriquecida en hormonas y va directamente a irrigar la pars distalis.<br />
El tracto supraóptico-hipofisario está formado por axones que transportan oxitocina y ADH, en este caso<br />
procedentes de las neuronas magnocelulares de los núcleos supraóptico y paraventricular del hipotálamo. Su<br />
recorrido es más largo y el transporte de las hormonas se realiza asociado a otra proteína. Cursan por el tallo neural<br />
y acaban vertiendo en la pars nervosa. La oxitocina la sintetizan las neuronas del paraventricular, mientras que las<br />
del supraóptico segregan ADH.<br />
GRÁNULOS SECRECIÓN<br />
Los axones de este tracto presentan dilataciones de trecho en trecho que se observan bien con sales de cromo y<br />
aluminio. Son de 3 tipos:<br />
1. Cuerpos de Herring: puntos de almacenamiento temporal de las hormonas oxitocina y ADH en multitud de<br />
gránulos de secreción de 10-30nm. Estas hormonas viajan a lo largo de los axones asociadas a la neurofisina<br />
(neurofisina I se asocia con oxitocina y la neurofisina II con ADH).<br />
2. Vesículas de 30nm que contienen Ach.<br />
3. Gránulos heterogéneos con tamaños de 50-80nm (semejantes gránulos de núcleo denso de la médula<br />
suprarrenal).<br />
HORMONAS<br />
La ADH y la oxitocina son hormonas secretadas por fibras nn diferentes pero que están formadas por ocho péptidos<br />
cada una y sólo difieren en 2 aa, porque tienen un precursor <strong>com</strong>ún.<br />
Cada hormona se asocia en su gránulo de secreción con la neurofisina correspondiente y ATP y son liberados ante<br />
estímulos determinados.<br />
La oxitocina se libera por estímulos nerviosos que alcanzan el hipotálamo. Pueden deberse a la succión del lactante<br />
(mama) o por distensión de la vagina y cuello uterino. La oxitocina estimula la contracción de los miocitos uterinos
Página109<br />
(orgasmos y menstruación), la contracción de los miocitos del útero grávido en el periodo perinatal (parto) y la<br />
contracción de las células mioepiteliales en la mama durante la lactancia (reflejo de eyección). En el varón estimula la<br />
contracción de la musculatura del conducto deferente.<br />
Su secreción se debe a reflejos sensoriomotores básicos <strong>com</strong>o la succión del lactante o la distensión del útero.<br />
En cuanto a la ADH, vasopresina u hormona antidiurética, aumenta la permeabilidad de la porción distal de la<br />
nefrona (túbulo contorneado distal y túbulos colectores). Se libera cuando hay una disminución en el volumen<br />
sanguíneo o un aumento en la presión osmótica del plasma. También responde a traumatismos, dolor, tensión<br />
emocional y determinadas drogas <strong>com</strong>o la nicotina. Se inhibe por la retención de agua.<br />
También es vasoconstrictora cuando aumenta en condiciones no fisiológicas.<br />
La diabetes insípida neurogénica se debe a un daño en las neuronas hipotalámicas que secretan ADH (ya sea por<br />
traumatismo craneal, destrucción autoinmune o un tumor). Los niveles de ADH bajan y se produce poliuria (hasta<br />
20L/día). En la diabetes insípida nefrogénica no hay respuesta a vasopresina, pero en la neurogénica sí la hay.<br />
GLÁNDULA PINEAL O EPÍFISIS<br />
La glándula pineal o epífisis se describe <strong>com</strong>o una glándula endocrina o neuroendocrina, aunque su función no está<br />
bien conocida en humanos.<br />
Mide 5-8mm de largo y 3-5mm de ancho. Pesa entre 100 y 200mg; encuentra en la línea media del encéfalo.<br />
Embriología<br />
En la 2º semana de gestación se origina la epífisis a partir del neuroectodermo de la porción P del techo del<br />
diencéfalo (línea media del tercer ventrículo) y permanece unida al encéfalo por un orto pedículo. Crece y empieza a<br />
plegarse hasta que finalmente se <strong>com</strong>pacta. Esta masa sólida de células va a estar rodeada por una fina cápsula de<br />
conjuntivo que deriva de las meninges (piamadre).<br />
Estructura Histológica<br />
Posee una CÁPSULA formada por piamadre que emite unos tabiques, que dividen en lobulillos el parénquima,<br />
formado por pinealocitos y células intersticiales. Los tabiques conducen nervios amielínicos y vasos al interior del<br />
parénquima.<br />
NO hay barrera hematoencefálica.<br />
Los pinealocitos (95%) son células grandes y pálidas que se disponen en nidos o cordones formando lobulillos. Bajo<br />
su membrana hay cisternas subsuperficiales que almacenan calcio. Tienen un cilio que involuciona tras la etapa<br />
embrionaria (9+0). Su núcleo es redondo y el nucléolo es prominente. Tienen desarrollo<br />
normal de orgánulos, con RER, escaso REL y aparato de Golgi, muchas mitocondrias,<br />
pequeñas inclusiones de lípidos y gránulos de secreción muy densos (200nm). Son<br />
características las sinapsis en cinta o barras sinápticas (<strong>com</strong>o en la retina y el oído<br />
interno). Estas neuronas presentan una o dos prolongaciones, a veces bastante largas,<br />
que terminan expandiéndose junto a un capilar. Hay grandes haces de microtúbulos.<br />
Las células próximas se unen por uniones en hendidura. Se apoyan en la lámina basal y<br />
son secretoras de melatonina.<br />
Se llaman células intersticiales son células gliales cuyos núcleos son más pequeños y<br />
oscuros. Son GFAP+, equivalente a los astrocitos. Se disponen entre los pinealocitos o<br />
formando la cápsula, y dan soporte a las células. Suponen el 5% de células de la<br />
glándula.<br />
PGFA = GFAP = Proteína Glial Fibrilar Ácida = son P fibrosas que forman los filamentos<br />
intermedios del citoesqueleto de células gliales <strong>com</strong>o astrocitos o células de Schwann
Página110<br />
En la epífisis hay acérvulos cerebrales, cuerpos arenáceos o arenilla cerebral, que son depósitos de fosfato cálcico<br />
depositados en los pinealocitos, hasta que los expulsan. Pueden llegar a cristalizar en la matriz extracelular sin alterar<br />
la función de la pineal. No es un proceso degenerativo porque los primeros cristales se observan en la primera<br />
década de vida y van aumentando con la edad.<br />
El hecho de que haya estos cristales sirve en la clínica <strong>com</strong>o punto de referencia: si los cristales están fuera de la línea<br />
media, hay algo que ha desviado la glándula de su punto normal, <strong>com</strong>o una hemorragia cerebral o un tumor.<br />
Inervación<br />
Tiene inervación simpática por fibras postganglionares que proceden del ganglio cervical superior y que llegan a la<br />
glándula por el nervio coronario. El sistema nervioso simpático estimula la síntesis de melatonina. Intervienen <strong>com</strong>o<br />
neurotransmisores la noradrenalina y el neuropéptido Y (NPY). La destrucción de forma bilateral el ganglio cervical<br />
superior lleva al cese de la secreción de melatonina.<br />
También hay fibras parasimpáticas que sinaptan con células ganglionares de la propia pineal. También hay otras<br />
fibras que vienen de niveles superiores del SNC, de la cisura posterior a través de la cintilla óptica.<br />
Funciones<br />
En vertebrados inferiores, esta estructura es el tercer ojo u ojo parietal ya que es un receptor de la información<br />
lumínica. Para ello, la epífisis está fuera del cráneo justo por debajo de la piel. La información lumínica se envía por el<br />
nervio pineal al cerebro.<br />
En los mamíferos, la epífisis está protegida por el cráneo y no es un fotorreceptor, pero sigue vinculada a la luz de<br />
forma indirecta. Sintetiza, a partir de serotonina, la hormona melatonina, que no se puede almacenar sino que según<br />
se estimula la glándula se sintetiza y expulsa. Esta síntesis <strong>com</strong>ienza cuando se apaga la luz, y los niveles máximos se<br />
dan por la noche. La exposición a la luz interrumpe la síntesis de melatonina.<br />
Hormona<br />
Los efectos de la melatonina son la inducción de la somnolencia y la regulación de ritmos, del que el más estudiado<br />
es el circadiano noche-día. Todas las funciones se desarrollan de día y de noche pero no de forma uniforme.<br />
La energía lumínica la recoge la retina y llega hasta las células ganglionares, que producen melanopsina. Esta<br />
melanopsina alcanza el núcleo supraquiasmático del hipotálamo, relacionado con el ganglio cervical superior, que<br />
inerva los pinealocitos. El núcleo supraquiasmático es el principal marcapasos endógeno, es decir, el principal<br />
responsable de la puesta en marcha de los ritmos que regulan las acciones fisiológicas. Este ritmo se transmite a<br />
todas las neuronas neurosecretoras del SNC.<br />
La melatonina juega un papel importante en la adaptación a los cambios súbitos en la curación del ciclo día-noche<br />
(jet lag). También es importante en las personas que tienen que hacer cambios crónicos en su ciclo día-noche (turnos<br />
laborales). Se ha relacionado con las respuestas emocionales que varían a lo largo de las estaciones (trastorno<br />
afectivo estacional). Estos patrones no aparecen en el recién nacido sino que están ya desde la etapa fetal gracias a<br />
la melatonina materna, que atraviesa la placenta. La melatonina actúa sobre el eje hipotálamo-hipofisario.<br />
En las especies donde la actividad sexual varía con las estaciones, es responsable de los periodos fértiles y de reposo<br />
sexual, siendo capaz de inhibir las gónadas, las gonadotropinas y los órganos anejos. En el hombre no tiene efectos<br />
sobre las funciones reproductoras. Más recientemente se ha puesto de moda <strong>com</strong>o posible fármaco<br />
antienvejecimiento porque es antioxidante (protege frente a radicales libres).<br />
Puede haber pinealomas, que son tumores agresivos que destruyen la pineal. Tienden a aparecer más en niños se<br />
asocian a una pubertad muy precoz (primera regla delante de los ocho años y espermatogénesis y secreción<br />
androgénica antes de los nueve años). Tiene efectos indirectos sobre las gónadas a través del hipotálamo.
Página111<br />
GLÁNDULA TIROIDES<br />
Es una glándula endocrina situada en la base del cuello, debajo de la laringe y delante de la tráquea. Está formada<br />
por dos lóbulos unidos por el istmo, aunque en ocasiones puede aparecer un lóbulo accesorio, el lóbulo piramidal,<br />
por encima del istmo. Tiene un color pardo rojizo.<br />
El tamaño de cada lóbulo es de 5cm en altura y un peso de 20-30g.<br />
Es una glándula endocrina que se encarga de la síntesis de calcitonina, T 3 (triyodotironina) y T 4 (tiroxina). Su síntesis<br />
responde a la TSH, sintetizada por células de la pars distalis hipofisaria, desde la semana 22 del desarrollo.<br />
Embriología<br />
Se forma a partir de una evaginación del endodermo en la base<br />
de la faringe. Comienza a detectarse en la cuarta semana del<br />
desarrollo y empieza a crecer en dirección caudal. Desciende a lo<br />
largo del conducto tirogloso (día 21) y cuando se sitúa frente la<br />
tráquea se divide en dos lóbulos (día 30). El conducto tirogloso<br />
supone una conexión transitoria con el agujero ciego. Una<br />
semana después el conducto tirogloso se rompe y la tiroides<br />
queda <strong>com</strong>o una masa independiente. En un porcentaje<br />
importante de personas (40%) no se reabsorbe correctamente el<br />
conducto tiroglosa (día 40) y pueden quedar restos del conducto<br />
y originar quistes o un seno tirogloso. En la novena semana del<br />
desarrollo, la masa se sitúa en su posición definitiva. Las células<br />
endodérmicas se organizan inicialmente en láminas y luego se<br />
pliegan formando folículos. En la semana 14 se detecta ya<br />
coloide, por lo que las células son funcionales.<br />
Independientemente de esto, se produce la formación de las<br />
células C, que derivan de la quinta bolsa faríngea. La quinta bolsa<br />
faríngea es una evaginación pequeña que aparece debajo de la cuarta bolsa faríngea, poblada por células que<br />
proceden de la cresta neural y que se denominan cuerpos ultimobranquiales. Proliferan, pierden la conexión con la<br />
bolsa faríngea y migran. Alcanzan la tiroides y allí se dispersan e incorporan en los lóbulos.<br />
Estructura general<br />
La tiroides está formada por una cápsula bastante gruesa de conjuntivo rica en colágeno. Se distingue una zona<br />
externa de conjuntivo laxo y una zona interna mucho más gruesa de tejido conectivo denso de la que parten<br />
tabiques que lobulan la glándula. Estos tabiques facilitan aislar partes de la glándula en cirugías.<br />
La zona interna tiene un parénquima y un estroma: el estroma es un tejido conjuntivo interfolicular en el que hay<br />
fibras reticulares y capilares fenestrados y el parénquima se organiza en folículos con células foliculares y células C.<br />
Los folículos son unidades estructurales y funcionales huecas en las que se delimita por un epitelio cúbico simple un<br />
espacio ocupado por el coloide, que es un material gelatinoso PAS+. Cada folículo mide de 50μm a 1mm. El coloide<br />
es considerado una secreción exocrina de las células foliculares y está formado por tiroglobulina y otras<br />
glucoproteínas y enzimas. No es una hormona, sino una forma inactiva en la que se almacenan temporalmente. En<br />
las preparaciones suele artefactarse.<br />
Células foliculares<br />
Las células foliculares son generalmente cúbicas pero su altura cambia en función de su grado de actividad. En<br />
situaciones de hipotiroidismo, <strong>com</strong>o en las deficiencias de yodo, tienden a aplanarse, pero el tiroides puede verse<br />
hipertrófico en respuesta a la TSH. En cambio, en el hipertiroidismo <strong>com</strong>o la enfermedad de Graves crecen en altura y<br />
hay exceso de T 3 y T 4 . Se apoyan en una lámina basal y alcanzan el coloide.<br />
Tienen gran desarrollo del RER y el aparato de Golgi, y se distinguen en ellas gránulos subapicales no muy grandes<br />
(200nm), grandes gotas de coloide (hasta 4μm), y finalmente otras estructuras delimitadas por membrana, unas
Página112<br />
vesículas que son muy densas. Los gránulos subapicales contienen tiroglobulina. Es una globulina sintetizada en el<br />
RER y el aparato de Golgi; estos gránulos se vierten al coloide por exocitosis. Las gotas de coloide son zonas de<br />
coloide que han sido endocitadas. Las vesículas muy densas son lisosomas unidos a las gotas de coloide que van a<br />
procesar la tiroglobulina.<br />
Las células foliculares presentan en su membrana plasmática muchas microvellosidades cortas y en determinados<br />
momentos pueden emitir pseudópodos que rodean las gotas de coloide (para su endocitosis). Presentan abundantes<br />
<strong>com</strong>plejos de unión, <strong>com</strong>o cualquier célula epitelial.<br />
Fases de la secreción<br />
Presentan dos fases:<br />
La primera es la fase exocrina, en la que su membrana plasmática basal capta el yoduro que circula por el<br />
plasma. Para ello presentan unos canales dependientes de ATP (ATPasa). Esta ATPasa se inhibe por el<br />
preclorato. Este transporte permite concentrar el ion yoduro en su interior (x200), que difunde en el interior<br />
de la célula y será vertido al coloide por la membrana apical.<br />
En esta misma fase la célula sintetiza la tiroglobulina en el RER y el aparato de Golgi. Sintetiza también un<br />
enzima, la peroxidasa tiroidea. Ambas se sintetizan en la misma vesícula y son exocitadas al coloide. La<br />
peroxidasa tiroidea, una vez en el coloide pero al lado de la membrana apical, se activa y transforma el<br />
yoduro en yodo, que se une con la tiroglobulina. De esta forma se produce la yodación de la tiroglobulina.<br />
<br />
La segunda fase es la fase endocrina. La presencia de TSH estimula los tirocitos para que endociten gotas de<br />
yodotiroglobulina gracias a los pseudópodos que pueden emitir hacia el coloide. Las gotas de<br />
yodotiroglobulina se dirigen por el citoesqueleto hasta los lisosomas, se fusionan con ellos y <strong>com</strong>ienza la<br />
degradación de la yodotiroglobulina. La yodotiroglobulina da lugar a varios productos (MIT, DIT, T 3 y T 4 ). Al<br />
final sólo tendremos T 3 y T 4 . En estas células es mucho más abundante la T 4 , que se encuentra en proporción<br />
20:1. Ambas difunden por la membrana basal y llegan a la sangre, donde se unen a proteínas séricas para<br />
circular. Estas células son las únicas capaces de sintetizar T 4 . La mayor parte de T 3 se forma en los tejidos<br />
periféricos a partir de la T 4 . El propiltiouracilo también bloquea la conversión de T 4 en T 3 en los tejidos<br />
periféricos.<br />
Hormonas<br />
La tiroxina o T 4 es exclusivamente formada en el hombre por las células foliculares de la tiroides. La triyodotironina o<br />
T 3 se forma sobre todo en tejidos periféricos (hígado, riñones y corazón) a partir de la T 4 . Estos dos tipos de<br />
hormonas son capaces de atravesar la membrana plasmática y unirse a unas regiones del ADN, las TRE.<br />
Función<br />
El principal efecto de las hormonas tiroideas es el aumento generalizado del metabolismo basal. Estimulan el<br />
consumo de oxígeno y aumentan la producción de calor, la actividad cardiaca y las secreciones endocrinas.<br />
Estimulan el crecimiento en longitud de los huesos independientemente del aumento en el consumo de oxígeno. Por<br />
tanto, un déficit de estas hormonas en la infancia hace que el niño crezca por debajo de lo que debería. Estimulan en<br />
particular la actividad de los osteoblastos.<br />
Las hormonas tiroideas maternas atraviesan la membrana placentaria y son esenciales porque influyen en el<br />
desarrollo inicial del SNC del feto. Las hormonas tiroideas fetales las encontramos desde la semana 14.<br />
Un déficit de T 3 y T 4 en el desarrollo fetal ocasiona un bajo número de neuronas, defectos en la mielinización y<br />
conlleva retraso mental. Son lesiones irreversibles del SNC. Esto ocurre tanto si el déficit viene de la madre antes del<br />
desarrollo del tiroides fetal <strong>com</strong>o si es del propio feto. Sin embargo, si el déficit es materno el retraso mental será<br />
grave.<br />
El cretinismo o hipotiroidismo congénito es un trastorno en el que hay ausencia congénita de tiroides. Faltan la T 3 y la<br />
T 4 , lo que conlleva falta de estimulación de las células somatotropas. Por ello, además de lesiones neurológicas<br />
irreversibles se produce enanismo.<br />
Por último, el bocio puede corresponder a distintas situaciones patológicas. Se puede dar con hipotiroidismo y con<br />
hipertiroidismo. El más corriente es el bocio endémico, que se debía a una falta de yodo en la dieta. La tiroiditis de<br />
Hashimoto es autoinmune y la enfermedad de Graves o bocio tóxico produce hipertiroidismo.
Página113<br />
El hipotiroidismo cursa con reducción del metabolismo basal, hipotermia e intolerancia al frío. Se reduce la<br />
sudoración y hay vasoconstricción cutánea. Si no se trata aparece mixedema, trastornos cutáneos, enlentecimiento<br />
de la actividad cerebral y somnolencia. En la enfermedad de Hashimoto hay hipotiroidismo que se debe a una<br />
reacción autoinmune. El organismo genera anticuerpos microsomales contra la peroxidasa tiroidea y la tiroglobulina.<br />
Se reduce el número de folículos, lo que conlleva una reducción funcional de la glándula.<br />
El hipertiroidismo se debe, por ejemplo, a la enfermedad de Graves. Es una enfermedad autoinmunitaria en la que el<br />
tiroides es hiperfuncional. Hay aumento en el número y tamaño de los folículos, pero hay escaso coloide. En estos<br />
pacientes el tiroides segrega de forma masiva T 3 y T 4 hasta que los niveles llegan a ser tóxicos. Los niveles de TSH son<br />
normales. Los linfocitos segregan IL1, TNF e interferón γ, que estimulan las células foliculares a producir más<br />
citosinas. Las plasmáticas sintetizan anticuerpos contra el receptor de TSH que se encuentra en la membrana<br />
plasmática basal de las células foliculares. Al imitar los efectos de la TSH, se producen grandes cantidades de<br />
hormonas tiroideas de forma incontrolada. Se encuentran infiltrados linfocitarios en los músculos extraoculares y el<br />
tejido orbitario. Acaban diferenciándose a adipocitos y forman edema y exoftalmos. Además, la T 3 estimula el<br />
fosfolambano, que es una proteína implicada en la liberación y captación de calcio en el retículo sarcoplásmico de los<br />
miocitos cardiacos. Esto provoca taquicardia y temblores.<br />
Células Parafoliculares o C<br />
Las células C son células procedentes de la cresta neural (forma parte del sistema neuroendocrino difuso) y les<br />
corresponde el 0.1% de la masa de la tiroides. Nunca se encuentran en contacto con el coloide. Forman parte del<br />
sistema neuroendocirno difuso, con la síntesis y secreción de calcitonina. Generalmente forman grupos pequeños o<br />
están aisladas en el epitelio glandular. Son difíciles de distinguir y se consigue gracias a la tinción con plata o actinio.<br />
Estas células C muestran desmosomas que las unen entre sí. Son células de mayor tamaño, más abombadas, con<br />
núcleos claros y grandes y citoplasma pálido. Tienen desarrollo de RER y aparato de Golgi, y destacan sus vesículas<br />
electrodensas pequeñas (60-150nm) que contienen calcitonina.<br />
Hormona<br />
El gen que codifica para la calcitonina (CGRP) se expresa en células del hipotálamo y la hipófisis. Es capaz de actuar<br />
<strong>com</strong>o vasodilatador y neurotransmisor. El déficit parcial o total (tras tiroidectomía) de esta hormona no conlleva<br />
ninguna enfermedad clínica.<br />
Funciones ↓CALCEMIA<br />
La calcitonina no responde al control hipofisario. Sus efectos son opuestos a los de la PTH. Estimula la calcificación<br />
del osteoide y disminuye la calcemia. También disminuye la actividad de los osteoclastos, con lo que disminuye la<br />
resorción ósea. Existe un bucle de retroalimentación positivo en el que la calcemia elevada estimula la secreción de<br />
calcitonina por las células C. Es independiente de la acción del hipotálamo e hipófisis.<br />
Hay carcinomas de tiroides que afectan a las células C y se llaman carcinomas medulares de tiroides. En estas<br />
personas hay producción excesiva de calcitonina y no hay alteración en los niveles de calcio ni repercusiones a nivel<br />
óseo. En la enfermedad ósea de Paget hay una actividad excesiva de los osteoclastos, por lo que hay resorción ósea<br />
exagerada. El hueso nuevo que se forma tiene una estructura mal organizada. La calcitonina se usa <strong>com</strong>o<br />
tratamiento del dolor en personas con metástasis esqueléticas. Tiene sus efectos a través del SNC.<br />
El oncocito, célula de Hürthle u oxífila que desarrolla un cáncer similar al carcinoma folicular, aunque tiende a ser<br />
más agresivos.
Página114<br />
GLÁNDULA PARATIROIDES<br />
Son cuatro pequeñas masas situadas en la parte posterior del tiroides. Hay dos superiores y dos inferiores. También<br />
se pueden incluir <strong>com</strong>o paratiroides unas glándulas accesorias que están en el cuello o el mediastino. Pesan 130g y<br />
tiene un color pardo amarillento.<br />
Las superiores derivan de la cuarta bolsa faríngea y las inferiores derivan de la tercera (<strong>com</strong>o el timo).<br />
Estructura histológica<br />
Cada una de estas masas está envuelta por una cápsula fina. Hay tabiques in<strong>com</strong>pletos pero no se habla de lóbulos.<br />
En el parénquima hay adipocitos infiltrados, sobre todo desde la pubertad (en el adulto son el 50% de la masa) y el<br />
estroma posee sinusoides y fibras reticulares.<br />
El parénquima se organiza en cordones anastomosados. Ocasionalmente se describen algunos folículos. Hay dos<br />
tipos de células: las principales y las oxífilas o acidófilas.<br />
Tipos celulares<br />
Las células principales son abundantes y pequeñas con núcleo pálido, grande y central, y citoplasma pálido. En<br />
reposo almacenan mucho glucógeno. En la fase activa hay poco glucógeno y desarrollo de RER y aparato de Golgi.<br />
Aparecen vesículas electrodensas con la hormona paratiroidea. Son las únicas que se ven hasta la pubertad.<br />
<br />
<br />
Claras: PAS+, con glucógeno y con vesículas de secreción; son INACTIVAS<br />
Oscuras: PAS- y con gránulos de secreción; son ACTIVAS.<br />
Las células oxífilas aparecen hacia los 6-7 años y tienden a estar en pequeños grupos dispersos. Son poco<br />
abundantes y su número va creciendo. Son mucho más grandes, su núcleo más pequeño y basófilo, y el citoplasma<br />
es muy eosinófilo. Tiene pocos orgánulos, pero muchas mitocondrias. No hay vesículas de secreción. Desarrollan<br />
desmosomas que las unen entre sí y con las células principales. No hay función conocida para estas células oxífilas.<br />
Se cree que son unas células de transición o las células principales en degeneración.
Página115<br />
Hormona<br />
Las células principales segregan PTH (parathormona). Esta proteína es sintetizada a partir de un precursor en el RER,<br />
y en el Golgi sufre proteólisis. Además, presentan en su membrana un receptor de calcio o CaSR (son sensibles a los<br />
niveles de calcio). Tienen función de control del mantenimiento de la calcemia en tejidos y líquidos.<br />
Cuando el calcio se une a CaSR se desencadena una cascada cuyo efecto final es la inhibición en la secreción de la<br />
PTH. Cuando el calcio es bajo, la célula aumenta la secreción de PTH.<br />
Funciones<br />
↑ CALCEMIA<br />
La PTH actúa regulando los niveles de calcio y de fosfato. Los niveles de calcio se controlan de una forma más lenta y<br />
sostenida que con la calcitonina, porquela calcitonina tiene efectos instantáneos muy breves y muy rápidos. La PTH<br />
promueve la reabsorción del hueso que ya está mineralizado. De esta forma aumenta los de calcio en sangre. Activa<br />
la osteólisis por osteoclastos.<br />
- La PTH actúa a través de los osteoblastos para activar los osteoclastos. Ante una hipocalcemia, la PTH<br />
promueve la síntesis de M-CSF (factor estimulador de colonias de macrófagos). Este factor difunde, se une a<br />
los monocitos que circulan en sangre y promueve su diferenciación a macrófagos. En estos macrófagos<br />
<strong>com</strong>ienza a expresarse el gen RANK. Cuando esto se produce, podemos decir que el macrófago está tomando<br />
la vía para convertirse en un osteoclasto.<br />
Un segundo efecto de la PTH sobre los osteoblastos es la inducción del gen de RANKL. RANKL y RANK se<br />
unen. Si esto ocurre, la consecuencia final será la diferenciación del precursor del osteoclasto a un<br />
osteoclasto inmaduro.<br />
También actúa sobre los osteoblastos induciendo la expresión de un gen que codifica para la<br />
osteoprotegerina. Compite con RANK para impedir que se una a RANKL. Esto impide que se formen<br />
osteoclastos maduros. De esta forma, la PTH controla la maduración de osteoclastos, pero no su acción<br />
directa.<br />
- También actúa a nivel renal y tiene varios efectos. Promueve la reabsorción de calcio en los túbulos<br />
uriníferos, lo que favorece la elevación de la calcemia. En los túbulos renales aumenta la reabsorción de<br />
fosfato (disminuye su excreción renal). Por último, promueve la producción renal de vitamina D activa a<br />
partir de la 25-OH vitamina D 3 .<br />
- En el tubo digestivo estimula la absorción de calcio (estimula la formación de 1,25-<br />
dihidroxicolecalciferol/1,25-OH) tanto de forma directa <strong>com</strong>o por la vitamina D activa. La hipercalcemia<br />
provoca una retroalimentación negativa sobre la PTH.<br />
El hipoparatiroidismo puede producirse de forma accidental en una cirugía de tiroides. Es mortal si no se corrige a<br />
tiempo ya que en 24-48h la bajada de calcio provoca excitabilidad nerviosa con parestesias y crisis de tetania y<br />
epilepsias. Causa asfixia y muerte si no se suministra PTH. También puede haber hipoparatiroidismo idiopático en el<br />
que la persona no puede responder bien a los niveles de calcio. Puede deberse a una mutación en CaSR, lo que hace<br />
que el organismo reaccione <strong>com</strong>o si estuviéramos en hipercalcemia constante, por lo que no segrega PTH.<br />
El hiperparatiroidismo puede deberse a la hipercalcemia benigna familiar. Es genética y se debe a una mutación en<br />
CaSR que impide responder a los niveles de calcio en sangre. En este caso se produce una secreción masiva de PTH.<br />
Esta secreción puede ser extremadamente grave si la mutación está en los dos alelos y habrá que extirpar el<br />
paratiroides. También puede haber hiperparatiroidismo por un tumor benigno funcional (adenoma). Un exceso de<br />
PTH lleva a fosfaturia hipercalciuria con formación de cálculos renales, hipercalcemia por desmineralización ósea y<br />
formación de quistes.
Página116<br />
GLÁNDULAS SUPRARRENALES<br />
Son una estructura par que se encuentra en el polo superior de cada riñón, en el tejido adiposo perirrenal. En el<br />
adulto pesan unos 10g. Su interior se divide en corteza y médula. Cada parte actúa de forma independiente desde el<br />
punto de vista embriológico y funcional. Están envueltas por una cápsula gruesa de la que salen tabiques.<br />
Embriología<br />
A la sexta semana se forma la corteza de la glándula. El epitelio del mesotelio invade el mesonefros y se forma una<br />
corteza fetal o provisional. Hay una segunda invasión un par de semanas después. Estas células no se mezclan con la<br />
corteza fetal, sino que forman la corteza definitiva o permanente. La corteza fetal es muy acidófila y constituye<br />
durante esta parte del desarrollo embrionario la mayor parte de la glándula (80%). La corteza definitiva tiene células<br />
más pequeñas y es más basófila. Tiene un aspecto que recuerda a la zona glomerular del adulto.<br />
Las células de la corteza fetal son bastante grandes y tienen gran desarrollo de los orgánulos del metabolismo<br />
lipídico. Se organizan en cordones celulares irregulares. Son controladas por la ACTH fetal y la placenta. La corteza<br />
definitiva está formada por células más pequeñas. La placenta y las suprarrenales colaboran para formar <strong>com</strong>puestos<br />
que no se pueden sintetizar de forma independiente, es decir, forman la unidad fetoplacentaria. Sintetizan<br />
glucocorticoides, aldosterona, andrógenos y estrógenos.<br />
En la etapa fetal no se distingue una médula <strong>com</strong>o tal. A partir de la séptima semana, algunas células de las crestas<br />
neurales alcanzan la glándula y quedan en la posición central. Esta invasión paulatina se produce a lo largo de todo el<br />
desarrollo embrionario, incluso en los momentos más terminales, hasta el nacimiento.<br />
En el feto, la glándula suprarrenal tiene un tamaño similar al riñón, siendo su pico máximo en peso al final del cuarto<br />
mes. Al final de la gestación, el feto es capaz de sintetizar unos niveles de esteroides dobles de los del adulto (100-<br />
200mg de esteroides al día).<br />
Tras el nacimiento, la corteza fetal involuciona. Esto supone una pérdida importante en peso (50%).<br />
Simultáneamente, la corteza definitiva <strong>com</strong>ienza a crecer y se constituyen las tres capas que conocemos en el adulto.<br />
Esto es un proceso largo que dura al menos cuatro años. Además de este proceso, las células cromafines<br />
procedentes de la cresta neural <strong>com</strong>ienzan a agregarse y se constituye por primera vez una médula de forma visible.<br />
Funciones en la etapa fetal<br />
Una función fundamental en el feto es la síntesis de la DHEA (DeHidroEpiAndrosterona). Es un andrógeno débil que<br />
pasa por la sangre hasta el sincitiotrofoblasto, donde se transforma en estrógenos. Los estrógenos se eliminan por la<br />
orina de la madre. Una caída brusca de los niveles de estrógenos en orina tiene valor diagnóstico <strong>com</strong>o posible<br />
muerte fetal.<br />
Además, en la corteza suprarrenal fetal se produce la síntesis de glucocorticoides, que pueden ser placentarios o<br />
maternos. Son esenciales en el desarrollo del feto. A partir del octavo mes son esenciales para sintetizar el<br />
surfactante por los neumocitos tipo II. También son esenciales en el desarrollo del eje hipotálamo-hipofisario. Por<br />
último, son responsables del proceso involutivo del timo a final del desarrollo y los primeros años de vida.<br />
Estructura histológica<br />
La corteza es la mayor parte de la glándula (80-90%). Tiene una tonalidad pardo-amarillenta y es esencial para la<br />
vida. Su origen es mesodérmico y sintetiza las hormonas esteroideas (derivadas del colesterol o corticosteroides). La<br />
cápsula emite tabiques hacia el interior de la corteza con vasos y nervios hacia el parénquima.<br />
La médula es el 10-15% de la suprarrenal. Su color es pardo-rojizo y no es esencial para la vida. Su origen es<br />
neuroectodérmico (crestas neurales) y sintetiza catecolaminas.
Página117<br />
Corteza<br />
Organización y tipos celulares<br />
Se organiza en tres zonas concéntricas,<br />
glomerular, fascicular y reticular. Las dos últimas<br />
actúan de forma conjunta formando una unidad<br />
funcional. La capa glomerular sintetiza<br />
mineralocorticoides y las otras dos,<br />
glucocorticoides y hormonas sexuales. Estas<br />
células no tienen gránulos de secreción.<br />
La zona glomerular es la más externa de las tres<br />
regiones corticales. Es un 10-15% del grosor total<br />
de la corteza. Las células endocrinas se organizan<br />
en cordones ovalados con forma de arco o grupo<br />
redondeado. Alrededor tenemos abundantes<br />
capilares fenestrados o sinusoides.<br />
La zona glomerular es muy basófila,<br />
hipercromática. Esto se debe a que las células<br />
están muy juntas y hay mucha densidad de<br />
núcleos redondos, pequeños y muy basófilos.<br />
Además, los citoplasmas son amplios y basófilos<br />
porque tienen muchas cisternas cortas de RER<br />
que forman grumos. También tienen mucho REL,<br />
Golgi y mitocondrias con crestas tubulares. Hay<br />
pequeñas gotas de lípidos de forma minoritaria.<br />
Estas células se encargan de sintetizar<br />
mineralocorticoides, sobre todo aldosterona.<br />
Carecen de 17α-hidroxilasa, por lo que no pueden<br />
sintetizar cortisol y ni esteroides sexuales. Tienen<br />
importante capacidad regenerativa. Cuando se<br />
produce un daño puntual <strong>com</strong>o una extirpación quirúrgica, a partir de la zona que quede se regenera la corteza<br />
<strong>com</strong>pleta (también las zonas fascicular y reticular).<br />
La mayor parte de la corteza (75%) la ocupa la zona fascicular. Las células se agrupan en cordones muy largos, rectos<br />
y paralelos entre sí. Junto a ellos hay capilares fenestrados rectos. Las células son más grandes, cúbicas y tienen un<br />
núcleo grande, central y bastante claro. En ocasiones pueden ser binucleadas. El citoplasma es acidófilo y muy<br />
espumoso, <strong>com</strong>o si estuviera lleno de burbujas. Esas burbujas son gotas de lípidos que pueden ser bastante grandes.<br />
Esto hizo que se llamaran espongiocitos. Tienen mucho REL y mitocondrias con crestas tubulares. También tienen<br />
RER y AG.<br />
Estas células no sintetizan aldosterona. Tienen 17α-hidroxilasa y 17,20-hidroxilasa. Sintetizan glucocorticoides y<br />
esteroides sexuales.<br />
Está regulada por el sistema hipotálamo-factor hipofisiario liberador de corticotrofina (CRF) – hormona<br />
adrenocorticotrófica (ACTH). Las células de esta zona se atrofian con la hipofisectomía, por lo que la ACTH se<br />
relaciona con el crecimiento y el mantenimiento celular, pero también estimula la síntesis de esteroides e<br />
incrementa el flujo sanguíneo a través de la glándula suprarrenal.<br />
La zona reticular es el porcentaje menor en grosor de la corteza (5-10%). Hay células endocrinas que forman<br />
cordones cortos y ramificados que se envuelven por capilares fenestrados o sinusoides. Las células son cilíndricas<br />
estrechas con citoplasma acidófilo. Podemos encontrar escasas gotas lipídicas y pueden acumular lipofuscina. Tienen<br />
muchos lisosomas y el desarrollo de REL y de mitocondrias tubulares no es tan exagerado.<br />
La mayor parte de su capacidad es la de sintetizar esteroides sexuales (DHEA y androstenodiona). Tienen una escasa<br />
secreción de glucocorticoides.
Página118<br />
Hormonas<br />
La síntesis de aldosterona en la zona glomerular de la corteza se regula mediante la angiotensina II. Las células<br />
glomerulares no responden al cortisol porque se convierte en cortisona en los hepatocitos. Una disminución de la<br />
tensión arterial o los niveles de sodio en plasma aumenta la secreción de renina lo que hace que se forme<br />
angiotensina I, que se transforma en angiotensina II en los pulmones. La angiotensina II estimula la síntesis de<br />
aldosterona.<br />
La regulación de los glucocorticoides de las zonas fascicular y reticular se controla fundamentalmente por ACTH. Hay<br />
dos situaciones patológicas ligadas a esta hormona, la enfermedad de Cushing y el síndrome de Cushing. En la<br />
enfermedad de Cushing hay aumento en la secreción de aldosterona, cortisol y andrógenos suprarrenales por un<br />
tumor en la adenohipófisis que provoca un exceso de ACTH. En el síndrome de Cushing tenemos un tumor en la propia<br />
corteza suprarrenal. Se produce hiperproducción de cortisol, aldosterona y andrógenos suprarrenales.<br />
Funciones<br />
Los mineralocorticoides, es decir, la aldosterona, actúa sobre todo sobre el túbulo contorneado distal y el túbulo<br />
colector del riñón aumentando la reabsorción de sodio y la excreción de potasio. Aumenta la retención de agua y la<br />
secreción renal de potasio e hidrogeniones.<br />
En el síndrome de Conn o aldosteronismo primario hay síntesis excesiva de aldosterona debido a un tumor en la zona<br />
glomerular. En el hiperaldosteronismo secundario hay aumento en la secreción de aldosterona por aumento en la<br />
secreción de renina (fallo en la regulación).<br />
Los glucocorticoides tienen distintos efectos, pero por su efecto en glucosa y glucógeno justifica el nombre dado.<br />
El cortisol:<br />
Efectos metabólicos: aumenta la glucemia (inverso a la insulina). A nivel hepático, los hepatocitos están<br />
estimulados y captan una mayor cantidad de aminoácidos y glucosa. Aumenta la síntesis de glucosa a partir<br />
de aminoácidos, la gluconeogénesis y la glucogenólisis. También estimula la degradación de proteínas. En el<br />
caso de las células adiposas, el cortisol promueve la movilización de los ácidos grasos.<br />
Efecto antiestrés: la administración de cortisol y glucocorticoides hace que el individuo esté más preparado<br />
para <strong>com</strong>batir situaciones de estrés y desgaste tanto físico <strong>com</strong>o psíquico. Por ejemplo, ayuda en<br />
infecciones, temperaturas extremas y cirugías.<br />
Efecto antiinflamatorio: los glucocorticoides inhiben la capacidad del individuo para responder a una lesión y<br />
deprimen la respuesta inmune tanto humoral <strong>com</strong>o celular. Estos efectos se relacionan con la inhibición de<br />
la colagenasa, la inhibición de la síntesis de leucotrienos, la migración de macrófagos y la inhibición de la<br />
proliferación de linfocitos. Se suministran en respuestas <strong>com</strong>o el asma, la artritis reumatoide y situaciones<br />
puntuales de alergia. También se aplican para evitar rechazos en situaciones de personas trasplantadas.<br />
Los grlucocorticoides circulantes pueden actuar directamente sobre la hipófisis, pero es más <strong>com</strong>ún que ejerzan su<br />
control por retroalimentación sobre las neuronas del núcleo arqueado del hipotálamo. Esto induce la liberación de<br />
CRF a la circulación portal hipotálamo-hipofisaria. También hay evidencias de que los glucocorticoides circulantes y<br />
los efectos fisiológicos que causan estimulan los centros encefálicos superiores, los cuales, a su vez, inducen la<br />
liberación de CRF por las células hipotalámicas.<br />
La DHEA y la androstenodiona son andrógenos (gonadocorticoides) débiles que pueden convertirse en testosterona y<br />
estrógenos. En la mujer, los andrógenos tienen <strong>com</strong>o fuente primaria la corteza suprarrenal. Son los responsables de<br />
la aparición de vello axilar y púbico en la pubertad.<br />
El síndrome adrenogenital conlleva la masculinización de la mujer, es decir, la hipertrofia del clítoris y el hirsutismo,<br />
por una producción excesiva de andrógenos.<br />
Las hiperplasias suprarrenales congénitas se deben a distintas mutaciones. La hiperplasia suprarrenal congénita<br />
lipoidea se relaciona con la mutación del gen que codifica para StAR (proteína reguladora estereidogénica aguda),<br />
que interviene en el transporte del colesterol a nivel de la membrana mitocondrial externa. Hay hipersecreción de<br />
ACTH e hiperplasia de esta glándula. Sin embargo, provoca defectos en la síntesis de los esteroides suprarrenales y<br />
gonadales. Otra hiperplasia suprarrenal congénita tiene que distintos enzimas de la síntesis de cortisol afectados. La<br />
corteza responde a ACTH y se produce hiperplasia cortical. La mayor parte de las personas con esta patología (90%)<br />
tienen mutado el gen de la enzima CYP21. La 17α-hidroxiprogesterona no puede convertirse en desoxicortisol. Los<br />
efectos son hipotensión y bajos niveles de sodio en plasma. Sin embargo, hay altos niveles de andrógenos. En<br />
lactantes niñas se observa virilización. El 10% de los pacientes tiene una mutación en el gen de la enzima CYP11. La
Página119<br />
desoxicorticosterona no puede convertirse en corticosterona. En este caso los pacientes retienen agua y sal, y son<br />
hipertensos.<br />
También se puede destruir la corteza suprarrenal. El síndrome de Waterhouse-Friderichsen se produce en lactantes y<br />
es una destrucción aguda de la corteza suprarrenal por septicemia por meningococos. Otra enfermedad más<br />
frecuente es una destrucción crónica y se conoce <strong>com</strong>o enfermedad de Addison. Es autoinmune o provocada por<br />
tuberculosis. El déficit en cortisol promueve una mayor secreción de ACTH y hay una mayor pigmentación de la piel.<br />
La falta de mineralocorticoides causa hipotensión con shock circulatorio. La falta de cortisol reduce las respuestas<br />
vasopresoras frente a catecolaminas y contribuye a la hipotensión. Determina la astenia.<br />
Médula<br />
Estructura histológica<br />
No hay un límite claro que separe la médula de la corteza. Está formada por cordones de células muy irregulares, las<br />
células cromafines (del sistema APUD) y también hay células ganglionares. Hay importante vascularización a base de<br />
capilares fenestrados y vénulas. Esta porción de la glándula tiene un origen embriológico diferente, no es necesaria<br />
para la vida y no tiene capacidad regenerativa.<br />
Las células ganglionares son neuronas simpáticas dispersas en medio de los cordones de células cromafines. Son<br />
difíciles de observar. Sus axones se dirigen a la corteza suprarrenal. Inervan la corteza y modulan la función de esas<br />
células endocrinas. Parte de los axones terminan a nivel de los vasos sanguíneos.<br />
Células cromafines<br />
Las células cromafines son las más abundantes y se agrupan en grupos ovoides y cordones cortos anastomosados.<br />
Son bastante grandes, pálidas y funcionalmente equivalen a neuronas simpáticas postganglionares sin<br />
prolongaciones. Las hormonas segregadas por la corteza suprarrenal ejercen un control sobre las células cromafines<br />
medulares impidiendo que emitan prolongaciones nerviosas.<br />
Tienen importante desarrollo de orgánulos, sobre todo RER y Golgi. En su citoplasma hay gran cantidad de vesículas<br />
en las que se da la síntesis y almacenamiento de catecolaminas. Las catecolaminas van unidas a cromograninas.<br />
Existen dos tipos de gránulos:<br />
o Gránulos grandes con núcleo denso y excéntrico que tienen noradrenalina.<br />
o Gránulos pequeños y homogéneos de núcleo menos denso que contienen adrenalina.<br />
Hay tinciones específicas <strong>com</strong>o la tinción cromafín y métodos argentófilos, así <strong>com</strong>o IHQ <strong>com</strong>o la cromogranina o la<br />
enolasa neurona específica.<br />
Hormonas<br />
La adrenalina y la noradrenalina representan menos del 20% del contenido de los gránulos de secreción medulares,<br />
porque además contienen unas P solubles llamadas cromograninas que permiten conferir la densidad al contenido<br />
granular fijando las catecolaminas mediante ATP y Ca 2+ .<br />
La noradrenalina se sintetiza dentro del propio gránulo con la colaboración de la dopamina-β-hidroxilasa. Se exporta<br />
al citosol para ser transformada a adrenalina mediante la PNMT o feniletanolamina N-metiltransferasa. Este enzima<br />
se sintetiza sólo cuando son inducidas por los glucocorticoides que llegan desde la corteza por sangre.<br />
La epinefrina vuelve a entrar en los gránulos de secreción. La reserpina es un fármaco que puede bloquear la<br />
excreción de estas catecolaminas al bloquear su transporte.<br />
La mayor parte de las células cromafines son ricas en adrenalina (80%) y sintetizan en porcentaje más bajo<br />
noradrenalina (20%). Ambas catecolaminas se segregan a la sangre. Hay datos de que estas mismas células pueden<br />
sintetizar y excretar dopamina y encefalinas, aunque no se sabe su función. Ambas catecolaminas, cuando se<br />
degradan, originan dos productos, el ácido vanilmandélico y la metanefrina, que se eliminan por la orina y se usan en<br />
clínica <strong>com</strong>o medición de la producción de catecolaminas.<br />
Los feocromocitomas son tumores en la médula suprarrenal. Suelen cursar con valores elevados de ácido<br />
vanilmandélico y metanefrina. Determinan hipertensión mantenida o episódica. En el síndrome de la neoplasia<br />
endocrina múltiple (MEN) hay tumores en distintas glándulas endocrinas, no solo en la suprarrenal.
Página120<br />
La médula suprarrenal está regulada fundamentalmente por fibras nerviosas simpáticas. Las fibras preganglionares<br />
son las que terminan inervando las células cromafines. Cuando se produce la estimulación simpática se produce la<br />
liberación de catecolaminas. Si se cortan las fibras, se dejan de producir catecolaminas.<br />
Funciones<br />
Las catecolaminas son moléculas que cuando se liberan dotan al organismo de mayor capacidad de respuesta ante<br />
situaciones súbitas catastróficas (emergencias). Dan lugar a respuestas rápidas que permiten salir de una<br />
emergencia. Permite afrontar un esfuerzo físico máximo.<br />
Los niveles basales de catecolaminas son bastante bajos. Ante una situación de estrés hay un pico muy alto que se<br />
puede correlacionar con diferentes parámetros. Para muchos autores, el verdadero producto de secreción es la<br />
adrenalina y la noradrenalina es sólo un precursor. Por ello, cuando se extirpan la suprarrenales disminuye la<br />
adrenalina, pero se siguen encontrando niveles de noradrenalina que proviene de los neurotransmisores.<br />
Las catecolaminas estimulan a nivel hepático la glucogenólisis (aumenta la glucemia) y la lipolisis (liberación de<br />
ácidos grasos libres a la sangre). Actúan a través de dos receptores:<br />
Receptores α-adrenérgicos: se expresan en miocitos lisos y provocan vasoconstricción.<br />
Receptores β-adrenérgicos: cuando las catecolaminas se unen a un receptor β-adrenérgico tendremos<br />
vasodilatación en los miocitos estriados y broncodilatación.<br />
Así, las catecolaminas producen aumento de la tensión arterial, frecuencia cardiaca, volumen minuto cardiaco y<br />
frecuencia respiratoria. Además, son responsables de la vasodilatación coronaria y esquelética y de la<br />
vasoconstricción en la piel y el tubo digestivo.<br />
Vascularización<br />
Encontramos diferentes plexos vasculares que proceden de tres arterias, superiores, medias e inferiores. Las<br />
inferiores vienen de la renal, las medias de la aorta y la superior de la frénica inferior. Hay un plexo a nivel capsular,<br />
otro a nivel cortical y otro a nivel medular. Por otro lado, hay unas arteriolas que irrigan la cápsula.<br />
Las arteriolas corticales darán lugar al plexo cortical. Origina capilares fenestrados rectos (sinusoides), que forman<br />
una red en la zona reticular y se vierten las distintas hormonas. Drenan por unas venas colectoras situadas a nivel<br />
corti<strong>com</strong>edular. No encontramos a nivel de la corteza grandes vasos venosos y no hay linfáticos.<br />
A nivel medular hay una doble irrigación. Hay sangre que alcanza la médula tras atravesar toda la corteza. No es muy<br />
rica en oxígeno pero sí lo es en hormonas. Estas hormonas permiten que las células cromafines sinteticen PMNT,<br />
necesario para transformar la norepinefrina en epinefrina. La otra vascularización procede de una arteriola que<br />
atraviesa la corteza pero no recoge hormonas. Llega más oxigenada y suministra la capacidad de respuesta rápida<br />
ante una situación de estrés. La sangre drena por una vena central y llegará a la vena suprarrenal. Las venas<br />
medulares poseen almohadillas y se en su pared hay desarrollo de los miocitos lisos en disposición longitudinal.<br />
Las células que secretan adrenalina son más abundantes en las zonas medulares irrigadas por la sangre que ha<br />
pasado antes por los sinusoides corticales por la activación de la PNMT por parte de los glucocorticoides. Así, la<br />
sangre llega directamente desde la médula a la corteza en un tipo especial de sistema porta. En cambio, aquellas<br />
zonas con irrigación de los capilares derivados de las arteriolas corticales son más ricas en noradrenalina.
Página121<br />
PÁNCREAS ENDOCRINO<br />
El páncreas endocrino está formado por islotes de Langerhans, que se disponen en el espesor del páncreas exocrino.<br />
Se encuentran formando entre el 1-2% del volumen pancreático (aproximadamente 1 millón de islotes), aunque son<br />
más numerosos en la cola.<br />
Estroma<br />
Está formado por una fina capa de fibras colágenas.<br />
Parénquima<br />
Los islotes tienen un diámetro de 100-500µm y están formados por varios<br />
tipos celulares que están profusamente irrigados por una red de capilares<br />
fenestrados.<br />
Aparecen 3 tipos celulares principales que mediante el fijado de Zenker-formol<br />
y la tinción de Mallory-Azán aparecen:<br />
• Células A o α (15-20%) son de color rojo. tienden a situarse en la zona<br />
periférica del islote. Su citoplasma tiene gran abundancia de vesículas<br />
o gránulos secretores pequeños (250nm) de centro muy denso y halo<br />
claro. Aunque la principal hormona que sintetizan es el glucagón, también se han encontrado GIP, CCK y<br />
ACTH-endorfina (precursor).<br />
• Células B o β aparecen de color pardo; se usa <strong>com</strong>o tinción específica el grimelius. Constituyen la mayor<br />
parte del islote (70%). Se sitúan en el centro. Sus gránulos son más grandes (300nm) y tienen un halo claro y<br />
un centro denso que suele aparecer cristalizado (insulina cristalizada). Se dividen activamente, por lo que<br />
continuamente se ajusta su número en función de los requerimientos de glucosa que tiene el organismo<br />
cada día.<br />
• Células D o δ aparecen de color azul; se usa <strong>com</strong>o tinción específica las sales de plata. Son un porcentaje bajo<br />
(5-10%) que se encuentra en la zona periférica del islote. Los gránulos tienen un tamaño superior a las<br />
células A y su matriz es homogénea en las vesículas. Son estimuladas por los mismos factores que las B. Las<br />
células D sintetizan y segregan somatostatina y gastrina.<br />
Mediante estas técnicas no se tiñen el 5% de las células, por lo que aparecen otros 3 tipos celulares que se conocen<br />
<strong>com</strong>o células accesorias pancreáticoendocrinas.<br />
También hay células G (productoras de gastrina) en los islotes.<br />
Vascularización<br />
Cada islote presenta vascularización propia que constituye el sistema porta ínsuloacinar. Dentro del islote y<br />
delimitándolo encontramos abundantes fibras reticulares y fibras del SN S y SN PS.<br />
Cada islote recibe una irrigación independiente de una arteriola aferente. y foma una red capilar fenestrada en él.<br />
Dentro del islote, la red capilar pasa primero por las zonas periféricas (células A y D) y posteriormente llega a la zona<br />
central, donde encontramos células B. Esto se conoce <strong>com</strong>o irrigación en cascada.<br />
La sangre con las hormonas es drenada por una vénula que drena a una vena porta que a continuación vuelve a<br />
formar una red capilar en el acino exocrino. Esto forma lo que se conoce <strong>com</strong>o el sistema porta ínsuloacinar.
Página122<br />
El hecho de que la sangre que pasa por el islote luego vaya a irrigar los ácinos exocrinos permite que las hormonas<br />
que irrigan el islote tengan efecto paracrino sobre las zonas exocrinas del páncreas. De esta forma, la síntesis de jugo<br />
pancreático será:<br />
<br />
<br />
Estimulada por la insulina, VIP y CCK<br />
Inhibida por el glucagón, el péptido P y la somatostatina.<br />
Hormonas<br />
INSULINA: es la secreción más abundante y actúa en H, mm. esquelético y tej. adiposo. Estimula la captación de<br />
G, almacenamiento de glucógeno y la glucolisis. Además estimula la síntesis de AG, inhibe la actividad lipasa de<br />
los adipocitos y favorece la captación de aa de la sangre e inhibe el catabolismo proteico.<br />
Es en general una hormona anabólica (excepto de hidratos de carbono).<br />
Deficiencia = DM I<br />
GLUCAGÓN: contrarios efectos a la insulina<br />
SOMATOSTONINA: inhibe la secreción de insulina y glucagón; actúa:<br />
Inhibe glucagón y libera insulina si: hiperglucemia, aa y AG libres altos en sangre, CCK o secretina.<br />
Inhibe insulina y libera glucagón: hipoglucemia, bajo nivel de aa y AG libres o altos niveles de insulina.<br />
Control Nervioso<br />
Los islotes de Langerhans están regulados por el SN S y SN PS, aunque sólo el 10% de las células están en<br />
contacto con neuronas:<br />
! SN PS> aumenta insulina y glucagón<br />
! SN S> en situaciones de estrés aumenta la insulina y disminuye el glucagón.
Página123<br />
TIPO<br />
CELULAR<br />
α<br />
β<br />
δ<br />
TIPO<br />
CELULAR<br />
TAMAÑO<br />
% HE<br />
POSICIÓN HORMONA<br />
ME:<br />
gránulos<br />
Grandes 15-20 Acidófilas Periféricas Glucagón Contenido denso y homogéneo de<br />
250nm<br />
Pequeñas 70 Basófilas Centrales Insulina Contenido de contorno cuadrangular<br />
(por insulina cristalizada) de 300nm<br />
5-10 Periféricas Somatostonina Gránulos más grandes con material<br />
menos electrodenso<br />
CÉLULAS<br />
ACCESORIAS PANCREÁTICOENDOCRINAS<br />
SECRECIÓN<br />
ACCIÓN<br />
PP/F<br />
(
Página125<br />
APARATO REPRODUCTOR FEMENINO<br />
Funciones<br />
Función reproductora: se encarga de formar los gametos femeninos y proporcionar un entorno adecuado para la<br />
fecundación. Si un gameto femenino une a un gameto masculino el aparato se modifica para propiciar la correcta<br />
implantación del embrión, su nutrición y crecimiento. Terminada la gestación, facilita la expulsión del feto maduro.<br />
Función endocrina: produce y secreta hormonas.<br />
Características<br />
Son característicos los cambios que sufre a lo largo de la vida, desde la infancia a la madurez reproductora y tras la<br />
menopausia. Se pueden constatar también cambios durante la época fértil de forma cíclica, que están regulados de<br />
forma endocrina.<br />
Generalidades embriológicas: la gónada indiferenciada inicial se desarrolla hacia ovario si las hormonas que<br />
desarrollan hacia la vía masculina están ausentes (factor determinante de testículo y factor inhibidor Mülleriano). Si<br />
no, se desarrollarán hacia testículo. Subsiguientemente se dará el desarrollo de las vías y genitales externos<br />
femeninos.<br />
Fases del desarrollo<br />
Como tal no se <strong>com</strong>pleta su desarrollo hasta la pubertad. La menarca es la primera menstruación, que es normal<br />
entre los 9-16 años. La telarca es el desarrollo de la mama femenina. La pubarca hace referencia a la aparición del<br />
vello pubiano, y la adrenarca es la aparición del vello axilar. Entre telarca y menarca suelen pasar unos dos años.<br />
Durante la vida fértil, en la mujer se producen los ciclos menstruales cada 28 días desde la pubertad hasta los 50<br />
años, cuando son más irregulares y se habla de climaterio. La menopausia es la última menstruación y a partir de ahí<br />
(posmenopausia) se atrofian todos su órganos lentamente.<br />
Componentes: genitales externos e internos<br />
Se estructura en genitales externos e internos. Los genitales externos son el monte de Venus, la vulva con los labios<br />
mayores y menores, y el clítoris. Los genitales internos son los ovarios, oviductos o trompas de Falopio, útero y<br />
vagina.
Página126<br />
GENITALES EXTERNOS<br />
Monte de Venus<br />
Es una eminencia sobre la sínfisis púbica que tiene una gran cantidad de grasa subcutánea. Se cubre tras la pubertad<br />
de vello pubiano largo con folículos pilosos profundos y oblicuos; el pelo es ensortijado en todas las razas.<br />
Vulva, labios mayores y menores<br />
La vulva está formada por dos pares de pliegues:<br />
Los LABIOS MAYORES son los más externos. Son unos pliegues cutáneos situados bajo el monte de Venus<br />
equivalentes al escroto masculino. Muestran glándulas sebáceas y sudoríparas ecrinas y apocrinas. Esta piel<br />
se asienta sobre una grasa subcutánea importante. Hay miocitos lisos que forman una capa equivalente a la<br />
túnica del dartos del escroto. Desde su cara externa se ve un labio mayor con piel cuya pigmentación<br />
aumenta tras la pubertad, así <strong>com</strong>o la cantidad de vello y desde su cara interna se ve una piel mucho más<br />
lisa, que carece de vello y que muestra abundantes glándulas.<br />
Los LABIOS MENORES son otro par de pliegues cutáneos más finos que encierran el vestíbulo vaginal y que<br />
quedan ocultos por los labios mayores. La piel de la cara externa de los labios menores se pigmenta más a<br />
partir de la pubertad. No presentan vello y la dermis que hay bajo el epitelio está muy vascularizada, es muy<br />
laxa y tiene importante cantidad de fibras elásticas. No hay grasa subcutánea. Encontramos abundantes<br />
glándulas sebáceas que se abren a la cara externa e interna de los labios. Presenta un epitelio estratificado<br />
con papilas altas.<br />
El vestíbulo vaginal es el espacio que queda entre los labios menores. A él se abren la uretra, la vagina y conductos<br />
de las glándulas vestibulares mayores y menores.<br />
Clítoris<br />
Debajo del monte de Venus encontramos el clítoris, que es el equivalente femenino del pene (formación eréctil).<br />
Aumenta de tamaño tras la pubertad y se hincha con la excitación sexual. Está formado por dos cuerpos cavernosos<br />
que acaban en una zona central, el glande. Cada cuerpo cavernoso está rodeado por una vaina fibrocolagenosa, que<br />
en su porción media se fusiona en un tabique central in<strong>com</strong>pleto. Estos cuerpos cavernosos están rodeados por una<br />
piel muy fina sin glándulas ni vello; pero rica en receptores sensitivos (corpúsculos de Pacini).<br />
Himen<br />
Es una membrana mucosa muy fina y delgada que cierra el paso hacia la vagina. Tiene un conjuntivo muy<br />
vascularizado y está tapizada en su cara externa (vulvar) por epitelio plano estratificado queratinizado y en su cara<br />
interna (vaginal) por epitelio plano estratificado no queratinizado. Es una zona de transición entre los genitales<br />
externos e internos.<br />
Glándulas<br />
Las glándulas que podemos encontrar en los genitales externos son sebáceas y sudoríparas, y están en la vulva y el<br />
monte de Venus (entre los labios menores). Las glándulas vestibulares menores se llaman también parauretrales o<br />
de Skene, y las mayores son las de Bartholin o vulvovaginales. Permiten lubrificar el vestíbulo y por tanto la entrada<br />
de la vagina, en especial durante la excitación sexual.
Página127<br />
Las glándulas vestibulares menores se encuentran en la zona que rodea al orificio uretral externo, cerca del clítoris.<br />
Son similares a las glándulas de Littré masculinas, de carácter mucoso y con epitelio prismático pseudoestratificado.<br />
Las glándulas vestibulares mayores son pares y se abren en la cara interna de los labios menores, cerca del orificio<br />
vaginal. Se sitúan a ambos lados del vestíbulo, en la parte inferior de la vagina. Son túbuloalveolares ramificadas y<br />
mucosas cuyo epitelio varía según la zona:<br />
‣ GLÁNDULA: epitelio simple cilíndrico<br />
‣ CONDUCTO: epitelio mucoso o epitelio prismático estratificado<br />
‣ DISTALMENTE: epitelio de transición y células mucosas del himen<br />
Inervación<br />
Los genitales externos tienen abundante inervación sensitiva. Hay terminaciones nerviosas libres en toda su piel.<br />
Además, hay corpúsculos de Meissner en los labios mayores y el monte de Venus, y corpúsculos de Pacini en los<br />
labios mayores y los cuerpos cavernosos del clítoris.<br />
La inervación es esencial para la respuesta fisiológica en la excitación sexual.<br />
OVARIO<br />
Generalidades<br />
Es un órgano par con dimensiones de 2.5-5x1.5-3x0.6-1.5cm. Tiene un color blanco rosado y es intraperitoneal. Se<br />
encuentra sujeto por el mesovario y ligamentos. Antes de la pubertad su superficie es lisa; tras la pubertad la<br />
superficie se va haciendo irregular debido a las cicatrices.<br />
Funciones<br />
1. Gametogénesis<br />
2. Estereidogénesis: proceso endocrino de síntesis de estrógenos que promueven el crecimiento y maduración<br />
de los órganos sexuales femeninos internos y externos, esenciales para el desarrollo de los caracteres<br />
sexuales femeninos, estimulan el crecimiento de los conductos y el estroma de las glándulas mamarias y<br />
regulan la acumulación de tejido adiposo.<br />
3. Síntesis progestágenos, que preparan el útero para el embarazo y la glándula mamaria para la lactancia.<br />
4. Sintetiza y segrega hormonas esenciales para llevar a cabo un correcto ciclo menstrual.<br />
En cuanto a las hormonas ováricas, desde el punto de vista bioquímico tenemos tres tipos de hormonas esteroides<br />
(derivadas del colesterol): los andrógenos, estrógenos y la progesterona.<br />
Dentro de los estrógenos destaca el estradiol, derivado de las células de la granulosa y de las luteínicas foliculares. Es<br />
la principal hormona en la vida fértil, mientras que en la postmenopausia predomina la estrona.<br />
El estradiol es el principal responsable de los caracteres sexuales secundarios femeninos. Estimula el crecimiento de<br />
la mucosa uterina, el de la mucosa vaginal (la engrosa) y es responsable de que el moco que encontramos en la luz<br />
vaginal sea un poco acuoso. En las glándulas mamarias estimula el crecimiento de los conductos y el estroma.<br />
También tiene efectos protectores para la salud de la mujer, fundamentalmente sobre el aparato cardiovascular.<br />
La progesterona la sintetizan las células de la granulosa y el cuerpo lúteo. Predomina en la segunda mitad del ciclo<br />
ovárico. Prepara la mucosa uterina para una posible implantación (fase secretora o progestacional) e inhibe las<br />
contracciones del músculo uterino. Hace que el moco vaginal sea más viscoso y actúa sobre las glándulas mamarias<br />
incrementando su crecimiento. Además, eleva la temperatura corporal basal.
Página128<br />
La mujer también sintetiza andrógenos, fundamentalmente androstenodiona y dehidroepiandrosterona, por las<br />
células de la teca interna. Tienen efecto masculinizante de la mujer y son convertidos en estrógenos por las células<br />
de la granulosa.<br />
Otras hormonas ováricas son la inhibina, activina, relaxina, oxitocina y vasopresina (ADH).<br />
Ciclo<br />
Se pueden establecer tres fases durante la etapa fértil: folicular, ovulatoria y luteínica. En la folicular, los folículos se<br />
desarrollan y aumentan de tamaño, de un folículo primordial a uno terciario. En la fase ovulatoria, el oocito sale<br />
fuera del ovario (ovulación). Con posterioridad describimos la fase luteínica, en la que el folículo que ha ovulado se<br />
transforma en un cuerpo lúteo que funciona <strong>com</strong>o una glándula endocrina.<br />
Estructura histológica<br />
Si observamos un ovario vemos que está tapizado por epitelio germinativo, bajo el que encontramos una lámina<br />
basal y un conjuntivo llamado túnica albugínea. Se divide en una zona cortical que rodea a la médula. Se describe un<br />
hilio, que es la zona de entrada y salida de vasos y nervios. Está unido por el mesovario al ligamento ancho.<br />
Epitelio germinativo<br />
El epitelio germinativo es un nombre equívoco ya que aquí no se forman los oocitos. Se continúa con el mesotelio<br />
del mesovario. En la época fértil es cúbico simple y el resto del tiempo es plano simple. Las células tienen<br />
microvellosidades en la zona apical y algunos cilios, abundantes vesículas de pinocitosis y a veces pueden quedar<br />
dentro de la corteza. Cada vez que la superficie se rompe y cicatriza las células pueden quedar dentro de la corteza y<br />
formar los islotes. Secreta líquido. Los islotes se denominan quistes de inclusión germinales.<br />
La túnica albugínea es conjuntivo denso con fibras y células paralelas a la superficie. Su matriz extracelular es<br />
blanquecina y está poco vascularizada.<br />
Corteza y médula<br />
La corteza se divide en folículos (primordiales, primarios, secundarios, terciarios, cuerpos lúteos y blancos) y tejido<br />
intersticial. Dentro del intersticial se distingue: el estroma (tejido intersticial <strong>com</strong>ún), la teca, glándula intersticial y<br />
células hiliares.<br />
El estroma es un conjuntivo entre los folículos, rico en células, vasos y fibras reticulares, pero no en elásticas, que<br />
sólo aparecen en proximidad a los vasos o formando parte de la media de esos vasos, que se sitúan alrededor de los<br />
folículos en desarrollo, atrésicos y los cuerpos lúteos. De forma dispersa se localizan miocitos lisos.<br />
Parte del estroma, concretamente el que rodea a los folículos, se diferencia para formar la teca. La teca muestra dos<br />
estratos: la interna adquiere carácter secretor y está muy vascularizada y la más externa está algo menos<br />
vascularizada y no es secretora, sino mucho más fibrosa.<br />
La glándula intersticial no es una glándula <strong>com</strong>o tal, sino acúmulos de células con aspecto epitelioide dispersas por<br />
toda la corteza del ovario. Estas células parece que proceden de las tecas hipertrofiadas de folículos atrésicos,<br />
fundamentalmente de gran tamaño. En nuestra especie son poco abundantes, pero en las que tienen muchas<br />
camadas sí son importantes. Las células epitelioides tienen gran cantidad de vesículas lipídicas y sintetizan y segregan<br />
andrógenos que luego se convertirán en estrógenos. Parecen tener un papel más importante y son más abundantes<br />
en la etapa infantil. A partir de la menarca involucionan.<br />
Las células hiliares tienen aspecto epitelioide y son bastante grandes y redondeadas. Abundan cerca del hilio y el<br />
mesovario. Histológicamente son idénticas a las células de Leydig del aparato genital masculino: pálidas y en su<br />
citoplasma abundan las inclusiones lipídicas, ésteres de colesterol, pigmentos lipocrómicos y cristales de Reinke. Se<br />
especula que contienen andrógenos. Estas células se observan en mayor cantidad en la gestación y la menopausia.
Página129<br />
Pueden ser causa de tumores cuando se malignizan o cuando hay desarrollo en gran número y tamaño (hiperplasia).<br />
En estos casos producen masculinización.<br />
La médula del ovario está formada por conjuntivo laxo con enorme desarrollo de vasos sanguíneos tortuosos<br />
(arterias helicinas y plexo venoso pampiniforme). También son importantes los linfáticos y las fibras nerviosas. Nunca<br />
hay folículos.<br />
Folículos: tipo y estructura<br />
Es una estructura de la corteza ovárica en la que siempre tenemos una célula germinal femenina a la que a<strong>com</strong>pañan<br />
otro tipo de células. El grado de desarrollo de la célula y el folículo es variable y la maduración de la célula germinal<br />
femenina y su expulsión del ovario se conoce <strong>com</strong>o ovulación. Al conjunto se le llama folículo y evoluciona y madura<br />
de forma sincrónica.<br />
En los folículos unilaminares la célula germinal está asociada a una sola capa de células a<strong>com</strong>pañantes, y en los<br />
multilaminares hay varias capas.<br />
Según su desarrollo, los folículos primordiales son los más pequeños (no más de 40μm). Son siempre unilaminares y<br />
están en la corteza ovárica, justo por debajo de la túnica albugínea. El oocito (si aparecen dos es un folículo<br />
poliovular) que encontramos es primario y está detenido en la profase I, en diplotena. Tiene de 25-30μm y está en el<br />
centro del folículo, con un núcleo bastante grande y nucléolo. El núcleo no tiene por qué estar centrado en la célula<br />
sino que generalmente es excéntrico. Posee el cuerpo de Balbiani, formado por el Golgi (un dictiosoma de gran<br />
tamaño, mitocondrias y RER). También hay laminillas anulares. Las células foliculares del folículo primordial también<br />
se pueden denominar granulosa (envuelta simple) porque son células epiteliales con una lámina basal.<br />
El folículo primario es más grande (80-100μm). Comienzan siendo unilaminares y acabarán siendo multilaminares,<br />
por lo que el tamaño varía. Según se desarrollan se van hundiendo dentro de la propia corteza, en el estroma. El<br />
folículo primario está constituido por un oocito y una granulosa separados por la zona pelúcida. Por fuera<br />
encontramos la membrana limitante externa y <strong>com</strong>ienza a formarse la teca.<br />
El oocito del folículo primario es bastante similar a la anterior. Sigue detenido en diplotena pero con mayor tamaño.<br />
Los <strong>com</strong>ponentes del cuerpo de Balbiani se han dispersado, aumenta el RER y los ribosomas y hay múltiples<br />
diplosomas pequeños dispersos. Tienen inclusiones lipídicas, pigmento lipocromo y abundantes gránulos corticales.<br />
En su membrana apical tiene gran desarrollo de las microvellosidades que penetran la zona pelúcida.<br />
La zona pelúcida es una matriz extracelular muy refringente que se observa cuando el oocito está creciendo. Está<br />
formada por glucoproteínas (PAS+). Sus <strong>com</strong>ponentes son sintetizados y segregados por las células foliculares y el<br />
oocito y en ella están las microvellosidades de la membrana del oocito y las prolongaciones finas que emiten las<br />
células foliculares.<br />
La granulosa pasa de ser una sola capa a crecer y hacerse las células cúbicas y prismáticas hasta formar varias capas.<br />
Contienen receptores de FSH, que controla su crecimiento y división. Estas células son epiteliales y por tanto no<br />
están vascularizadas y al estar unidas por uniones GAP (y no hermeticcas), no forman una barrera hematofolicular<br />
que separe el oocito de la sangre que hay en los vasos próximos. Además, los nexos que se establecen con el oocito<br />
son fundamentales para permitir la sincronización y maduración de este. Según crecen también aumenta su dotación<br />
de orgánulos: crecen el RER, Golgi, ribosomas y mitocondrias. Estas células son capaces de producir testosterona,<br />
pero previamente necesitan androstenodiona de las células de la teca; finalmente la testosterona se convertirá en<br />
estradiol.<br />
Por fuera de la granulosa encontramos la membrana limitante externa, que es una lámina basal que separa la<br />
granulosa epitelial de la teca, que es un conjuntivo.<br />
La teca se diferencia alrededor de la granulosa a partir del estroma y se observa un estrato interno y otro externo sin<br />
que haya un límite nítido. Inicialmente la teca interna son fibroblastos que están alrededor de la granulosa. Presenta<br />
una gran vascularización, una importante cantidad de fibras, fundamentalmente colágenas, y los fibroblastos
Página130<br />
acabarán transformándose en células secretoras. Tienen desarrollo marcado del REL, mitocondrias con crestas<br />
tubulares e importantes cantidades de lípidos. Presentan receptores para la LH, que regulan la síntesis y secreción de<br />
androstenodiona.<br />
La teca externa es la porción más periférica formando fibras concéntricas alderedor de la teca interna; está formada<br />
por conjuntivo denso muy vascularizado que se continúa con el estroma cortical; también hay miocitos lisos.<br />
El folículo secundario o folículo antral tiene un tamaño considerable pero variable. Tiene los mismos <strong>com</strong>ponentes<br />
del folículo primario, pero entre las múltiples capas de la granulosa se <strong>com</strong>ienzan a abrir huecos. Estos espacios<br />
pequeños irregulares se llaman cuerpos de Call-Exner y están llenos de líquido antral (PAS+). Estas cavidades van<br />
convergiendo de forma que en el folículo secundario maduro hay una única cavidad llamada cavidad antral. El líquido<br />
antral tiene ácido hialurónico, esteroides, GF y gonadotropinas.<br />
En el folículo secundario encontramos el oocito detenido en diplotene. Es una célula muy grande (125-150μm) que<br />
deja de crecer gracias al factor inhibidor de la maduración oocítica (OMI) segregado por la granulosa; si no se<br />
secretara, la meiosis superaría el bloqueo y ocurriría una maduración meiótica espontánea. El oocito ahora se<br />
encuentra situado en el centro de la cavidad antral, pero no en el centro del folículo. A su alrededor está la zona<br />
pelúcida y una cada de células foliculares que se denominan en conjunto corona radiada.<br />
Las células de la granulosa presentan receptor de FSH. En las que forman la corona radiada se aprecia que su<br />
membrana apical forma microvellosidades que crecen en número y tamaño según progresa el desarrollo del folículo<br />
y se dirigen hacia la zona pelúcida. Se establecen nexos entre las microvellosidades de las células foliculares y las de<br />
la membrana del oocito. El conjunto formado por oocito y corona radiada está unido con el resto de células de la<br />
granulosa por una formación llamada cumulus oophurus, disco prolígero, cúmulo oóforo o colina germinal.<br />
Alrededor del folículo secundario y formando parte de él encontramos una teca <strong>com</strong>o la de los primarios pero con un<br />
desarrollo mucho mayor.<br />
El folículo maduro, terciario o de Graaf es una estructura con desarrollo máximo que puede llegar a tener 2cm y<br />
perfectamente visible porque protruye sobre la superficie ovárica. Hasta este momento las células de la granulosa se<br />
dividían de forma activa y crecían en capas; entonces las mitosis se ralentizan mucho y las células presentan<br />
receptores para FSH y LH. La corona radiada es una capa de células cúbicas que empieza a perder la con las células<br />
de la colina germinal. La teca se desarrolla más y el folículo está listo para la ovulación.
Página131<br />
Ovulación<br />
Es la liberación del oocito de un folículo de Graaf junto con la corona radiada que hay a su alrededor. Ocurre de<br />
forma cíclica durante la vida fértil en torno al día 14 tras un pico de LH, bajo control endocrino de LH y FSH.<br />
Justo antes de la ovulación, la colina germinal se rompe de forma que el oocito y corona radiada quedan flotando en<br />
la cavidad antral y el líquido folicular se hace más acuoso (fluido). Justo por encima del folículo de Graaf<br />
encontramos una pequeña cantidad de estroma y la túnica albugínea, que deja de ser perfundida y se torna más<br />
transparente, <strong>com</strong>o hialina; se habla de que aparece un estigma o mácula pelúcida.<br />
Poco después de que aparezca el estigma el epitelio germinal que está por encima se rompe y desintegra. Cuando<br />
hay niveles máximos de estrógenos y se constata un aumento importante en la cantidad de enzimas proteolíticas en<br />
la teca externa y la túnica albugínea, por lo que el estroma se adelgaza, facilitándose la rotura de la pared del ovario.<br />
Fuera y al lado del ovario se constatan cambios en la trompa. La zona más proximal se acerca hacia la superficie del<br />
ovario de forma que cuando llegue el momento de la ovulación el oocito entre en la luz del oviducto en vez de<br />
quedar en la cavidad peritoneal. En un porcentaje bajo los oocitos caen a la cavidad peritoneal y pueden llegar a<br />
fecundarse e implantar. Estas implantaciones que no ocurren en el útero se llaman implantaciones ectópicas. Las más<br />
frecuentes son las de la propia superficie del ovario, intestino y en el saco rectouterino de Douglas. Son<br />
implantaciones de riesgo que se deben detectar de forma precoz. En la mayoría de los casos se interrumpen de forma<br />
natural. Normalmente los oocitos que caen degeneran.<br />
Otro proceso previo a la ovulación es la maduración del oocito, que por fin supera el diplotene; esto permite que se<br />
<strong>com</strong>plete la primera división meiótica. La envuelta nuclear, que hasta entonces estaba intacta, se desintegra gracias<br />
al factor promotor de la ovulación (MPF). La primera división meiótica da dos células desiguales: la que recibe la<br />
mayor parte de orgánulos es el oocito secundario y la otra es el primer corpúsculo polar. Ambas células quedan<br />
juntas rodeadas por la zona pelúcida y corona radiada, y se produce el inicio de la segunda división meiótica. Se<br />
detiene en metafase gracias al factor citostático (CSF) de forma que en la ovulación sale el oocito detenido en<br />
metafase II y por lo tanto cuando se produce la fecundación se fecunda directamente el oocito II. Si esto ocurre, se<br />
reanuda y se <strong>com</strong>pleta la segunda división meiótica y se originan dos células hijas desiguales, el óvulo y un segundo<br />
corpúsculo polar (ambos corpúsculos polares se degenerarán).<br />
Los oocitos primarios son células diploides con doble cromosoma X (44XX), pero los secundarios son haploides con<br />
un cromosoma X (22XX). Al fecundarse se restablece la diploidía pudiendo ser XX o XY.<br />
Cuerpo lúteo<br />
El folículo terciario tras la ovulación se colapsa y las células foliculares y la teca interna se transforman en una<br />
glándula endocrina llamada cuerpo lúteo. Se produce una fuerte contracción de la porción externa de la teca y el<br />
folículo se pliega y ya no se observa una cavidad más o menos clara.<br />
Se produce una ligera hemorragia hacia la cavidad central que forma un coágulo y la membrana limitante externa<br />
desaparece y los vasos de la teca invaden la granulosa; estos vasos invaden la zona del coágulo y se produce una<br />
llegada de fibroblastos, fibras de colágeno y angiogénesis. La zona central se hace muy fibrosa y alrededor granulosa<br />
y teca interna se convierten en las células luteínicas, quedando el centro fibroso y las células luteínicas periféricas.<br />
Estas células luteínicas son de mayor tamaño y su metabolismo se orienta a la síntesis de esteroides. Hay importante<br />
desarrollo de REL, mitocondrias con crestas tubulares y el citoplasma tiene muchas inclusiones lipídicas y luteína<br />
(pigmento lipocromo). Sintetizan y segregan estrógenos y progesterona, según estimulación hormonal. Hay dos tipos<br />
de células luteínicas:<br />
Las más periféricas son las células luteínicas de la teca interna, son más pequeñas (15μm) y se tiñen más y disminuye<br />
su número porque la teca externa va invadiendo el cuerpo lúteo creando tabiques. Así, para localizarlas tendremos<br />
que ponernos al lado de esos tabiques. Estas células sintetizan progesterona y androstenodiona y responden a la LH.<br />
El resto del cuerpo lúteo está formado por células luteínicas foliculares, más numerosas y mayores en tamaño.<br />
Sintetizan y segregan progesterona y estrógenos en respuesta a FSH y LH. Carecen de la enzima para sintetizar<br />
estradiol, por lo que parten de la androstenodiona de las células tecoluteícas.
Página132<br />
La progesterona y andrógenos estimulan el crecimiento de la mucosa y el crecimiento y actividad de las glándulas del<br />
endometrio y preparan el endometrio para la implantación y su desarrollo es máximo en torno al día 20.<br />
Si en el día 20 no ha habido fecundación, el cuerpo se denominará cuerpo lúteo de la menstruación y se degradará<br />
por luteólisis con procesos apoptóticos. Habrá un cese en la perfusión que llevará a una situación de hipoxia que se<br />
a<strong>com</strong>paña de una llegada masiva de linfocitos T que sintetizan TNF-γ, que activa la llegada de macrófagos que<br />
sintetizan TNF; así las células luteínicas acaban muriendo. Se calcula que el cese de actividad de estas células está en<br />
torno al día 26 y al degenerar forman el cuerpo blanco.<br />
Si en torno al día 20 se produce fecundación se habla del cuerpo lúteo de la gestación. Sigue creciendo hasta medir<br />
unos 4cm e invade prácticamente todo el ovario. Mantiene la actividad de síntesis hormonal de estrógenos y<br />
progesterona, pero ahora bajo el control de la gonadotropina coriónica humana (hCG) del trofoblasto. Esta actividad<br />
endocrina es fundamental para que el endometrio se mantenga preparado para la ovulación. El cuerpo lúteo es<br />
esencial en las primeras semanas de vida y se irá sustituyendo por la placenta, hígado y corteza suprarrenal<br />
femenina. A partir del sexto mes hay una clara involución del cuerpo lúteo (luteólisis) que no implica menstruación<br />
porque los niveles de FSH no caen. También se sabe que el cuerpo lúteo segrega relaxina. Fundamentalmente se<br />
detecta en el primer trimestre de la gestación inhibiendo las contracciones peristálticas espontáneas del útero<br />
relajando la musculatura y facilitando el parto. Estimula la síntesis de colagenasa e inhibe la síntesis de colágenos,<br />
relajándose los ligamentos de la sínfisis del pubis y el ablandamiento del cuello uterino de cara al parto.<br />
Cuerpo blanco<br />
El cuerpo blanco o albicans pierde tamaño y se va hundiendo en el estroma de la corteza ovárica y persiste durante<br />
años. Se denomina así porque las células periféricas incrementan su actividad secretora y disminuyen su contenido<br />
en lípidos, cesando la síntesis de estrógenos. Las células periféricas se hacen secretoras de colágeno, por lo que cada<br />
vez hay más colágeno que se agrupa formando tabiques fibrosos que tabican el cuerpo lúteo. La glándula endocrina<br />
pasa a ser una masa ovoide fibrosa de color hialino pobre en células y rica en colágeno.<br />
Desarrollo folicular<br />
En la etapa embrionaria, en la tercera semana las células germinales primitivas migran hacia la cresta gonadal,<br />
pasando la pared del tubo digestivo y llegando al blastema gonadal. Pueblan esta cresta e invaden los cordones. Sin<br />
embargo, las células germinales en este caso quedan rodeadas por las células epiteliales derivadas del epitelio<br />
peritoneal (celómico), que formarán la granulosa. Así se forman los folículos primordiales. Este desarrollo es<br />
independiente de los niveles de gonadotropinas. Las células siguen llegando y se dividen de forma activa. Durante la<br />
10-11 semana ya se han detenido en diplotene y se habla de folículos primarios. Se calcula que en el 5º mes de vida<br />
todas las células han entrado ya en diplotene, y en el mes 6-7 hay unos 7 millones de oocitos.<br />
Al nacer sólo queda un millón y a la pubertad sólo llegarán unos 400.000. En la pubertad los folículos ya son<br />
dependientes de gnRH, FSH y LH. En cada ciclo menstrual un pequeño grupo de folículos primarios se activa y se<br />
desarrollan a folículos secundarios. En principio solo uno se convertirá en folículo dominante. Se cree que eso<br />
depende del nivel de receptores para la FSH de la granulosa. Tiene mayor crecimiento y es el que madurará para<br />
convertirse en un folículo de Graaf. El desarrollo es dependiente de gonadotrofinas. Poco antes de la ovulación el<br />
ovocito del folículo dominante prosigue la primera división meiótica y se detiene en metafase II. A lo largo de la vida<br />
sólo maduran unos 400 oocitos. Tras el tiempo fértil el ovario ha perdido todos los oocitos y la mujer entra en<br />
menopausia.<br />
Atresia<br />
En la mujer hay degeneración absoluta de los folículos formados en el desarrollo embrionario. No es patológico y<br />
afecta a más del 99% de los folículos formados en el embrión. Afecta más a los folículos pequeños, pero pueden<br />
sufrirla cualquier tipo de folículo desde el desarrollo embrionario y después del nacimiento, aunque a los 40 años la<br />
velocidad de degeneración dosminuye. Un folículo primordial y primario no deja huella, pero el secundario se<br />
transforma en un cuerpo fibroso difícil de distinguir del cuerpo blanco. Se queda en el estroma ovárico durante<br />
mucho tiempo formando una cicatriz.
Página133<br />
Los folículos primarios y primordiales sufren atresia por degeneración del oocito. Entra en apoptosis y después las<br />
células foliculares degeneran. Se retrae y el espacio que queda por la retracción es invadido por el estroma que lo<br />
rodea.<br />
Cuando degenera un folículo secundario se constata inicialmente en la pared. La degeneración del oocito es<br />
secundaria a la de la teca y la granulosa. Las células foliculares presentan un factor NAIP (proteína nerviosa<br />
inhibidora de apoptosis) que inhibe la apoptosis de estas células. Se expresa en todos los folículos pero cuando va a<br />
empezar a sufrir atresia un folículo se interrumpe su síntesis.<br />
Un folículo atrésico tiene un oocito excéntrico y el núcleo es picnótico. Hay una clara alteración del citoplasma del<br />
oocito y las células foliculares ya que están en apoptosis. La zona pelúcida se mantiene aparentemente intacta y los<br />
vasos que estaban a nivel de la teca invaden la granulosa y la colina germinal, vascularizando esa zona. Un signo más<br />
tardío de atresia es la dispersión de las células foliculares y la llegada de macrófagos, fibroblastos y neutrófilos a la<br />
granulosa y más tarde la invasión del antro por tejido conjuntivo. La membrana limitante externa se engruesa y sufre<br />
plegamientos, llamándose membrana vítrea o esmerilada. Se constata mayor separación entre granulosa y teca. Por<br />
último, la teca se hipertrofia y las células epitelioides tienen un aspecto que recuerda bastante a las luteínicas, que se<br />
encargan de sintetizar estrógenos (metabolismo lipídico).<br />
Los folículos atrésicos en estado avanzado (o cuerpos lúteos atrésicos) recuerdan mucho al cuerpo blanco. El<br />
estroma invade el folículo en degeneración y los cordones de células de la teca se fragmentan y dispersan. Además,<br />
parece que las células de la glándula intersticial proceden de la teca de estos folículos atrésicos.
Página134<br />
OVIDUCTO O TROMAS DE FALOPIO<br />
Características<br />
Es un tubo muscular de unos 12cm con los extremos abiertos: uno hacia la cavidad peritoneal y el otro hacia la<br />
cavidad uterina. Proporcionan el entorno adecuado para la fecundación; es donde se forma el cigoto y <strong>com</strong>ienza la<br />
segmentación.<br />
Partes<br />
Cada zona tiene diferencias histológicas y son: una primera zona es el pabellón o infundíbulo, luego tendríamos la<br />
ampolla, el istmo más corto y el segmento intramural, que es el más corto de todos.<br />
El infundíbulo es el extremo abierto hacia la cavidad peritoneal y presenta flecos denominados fimbrias. Justo antes<br />
de la ovulación las fimbrias aumentan su perfusión y aumentan de tamaño, lo que permite que el infundíbulo se<br />
acerque a la superficie del ovario que va a ovular para asegurar que el óvulo liberado entre dentro y no se caiga.<br />
La ampolla tiene un mayor diámetro con luz amplia y unas paredes relativamente delgadas. En esta zona ocurre la<br />
fecundación. Aquí el óvulo es transportado de forma lenta. El óvulo fecundado puede implantarse en la mucosa del<br />
oviducto. Este embarazo ectópico no progresa porque el oviducto se rompe produciendo una hemorragia interna.<br />
El istmo es una zona larga que presenta una luz muy estrecha y unas paredes musculares bastante gruesas que<br />
permite contracciones rítmicas para el avance del óvulo hacia el útero.<br />
El segmento intramural une el oviducto con la luz uterina, atravesando la pared uterina.<br />
Estructura histológica<br />
La luz es estrecha y laberíntica y la mucosa está llena de pliegues; son mayores en la zona de la ampolla, así <strong>com</strong>o<br />
más <strong>com</strong>plejos y se ramificados. En el istmo son más bajos y están menos desarrollados y mínimos en el segmento<br />
intramural.<br />
La estructura en capas (mucosa, muscular y adventicia) sufre cambios cíclicos.<br />
El epitelio es prismático simple cuya altura decrece al avanzar hacia el útero, siendo máxima a la altura en la ampolla;<br />
hay células basales, ciliadas y secretoras.<br />
Las células ciliadas se encargan de facilitar el transporte del óvulo a nivel del infundíbulo y la ampolla. El número de<br />
células ciliadas, el número de cilios y la altura de los cilios varían de forma cíclica siendo máximos en el momento de<br />
la ovulación, aunque después irán perdiendo peso y crecen tras la menstruación y el parto. Los cambios en estas<br />
células son muy marcados en infundíbulo y ampolla, y se controlan por el estradiol y la progesterona. Si alguna de las<br />
hormonas falla (<strong>com</strong>o en la ovariectomía) se atrofia el epitelio del oviducto y las células ciliadas desaparecen. El<br />
epitelio se recupera al administrar estradiol.<br />
Las células secretoras son más bajas tras la ovulación. Están provistas de microvellosidades apicales no muy altas y<br />
segregan fundamentalmente un fluido rico en agua, potasio, cloro y algunas proteínas séricas, que proporciona<br />
nutrientes al oocito tras la ovulación y en los primeros días de la segmentación si hay fecundación.<br />
La lámina propia es bastante laxa y destacan las fibras reticulares. Hay fibroblastos y células del sistema inmune, así<br />
<strong>com</strong>o nódulos. Está muy vascularizada y en la mujer gestante se aprecia un aumento en el tamaño de las células.<br />
La capa muscular está muy desarrollada (gruesa) y regulada por endocrino.. Tenemos dos estratos, pero en las<br />
porciones más próximas al útero hay tres. Los miocitos se disponen de forma espiralada en una capa interna circular<br />
y otra externa longitudinal con contracciones peristálticas máximas en la ovulación. También son responsables del<br />
avance del embrión hacia el útero.<br />
Por último tenemos una adventicia que forma una serosa externa. Se corresponde con el peritoneo.
Página135<br />
Cambios cíclicos<br />
En la ovulación hay máximo desarrollo de los cilios y su batido, y de los vasos y la perfusión sobre todo a nivel de la<br />
ampolla, donde aumenta la turgencia que permite el desarrollo de las fimbrias y el acercamiento hacia el ovario.<br />
También hay aumento importante en las contracciones peristálticas desde el infundíbulo hacia el útero.<br />
ÚTERO<br />
Generalidades<br />
Es un órgano impar hueco situado en la zona de la pelvis menor. Se describe en forma de pera, aplanado en el plano<br />
AP. Su pared es gruesa por el desarrollo de la capa muscular.<br />
Funciones<br />
Recibe el blastocisto y permite su implantación. Después sufrirá cambios que permitan que permanezcan las<br />
relaciones vasculares y el embrión se nutra y crezca; una vez maduro el feto, el miometrio contribuye a su expulsión.<br />
Cuerpo, istmo y cérvix<br />
Anatómicamente se distinguen los dos tercios superiores que son el cuerpo y el tercio inferior o cérvix, entre los que<br />
se dispone una zona estrecha o istmo.<br />
En el cuerpo ocurre la implantación. Dentro de él hay una zona superior a los orificios en los que desembocan las<br />
trompas, el fondo. El orificio interno permite <strong>com</strong>unicar la luz del cuerpo con el cérvix.<br />
El istmo es una región muy corta y estrecha que está en la zona superior del cuello. Histológicamente es igual al<br />
cuerpo pero con una mucosa más delgada. En la mujer gestante no se distingue y pasa a integrarse en el cuerpo.<br />
El cuello o cérvix es una parte en forma de tubo con los extremos más estrechos. Es simétrico y tiene una pared<br />
bastante delgada cuyo orificio externo <strong>com</strong>unica el cuello con la vagina; en principio es redondo, pero tras un parto<br />
natural adquiere la forma de hendidura transversal. La porción del cuello que sobresale en la vagina se denomina<br />
porción vaginal. En las mujeres que ya han tenido embarazos el cuello adquiere la forma de un tonel o barril.
Página136<br />
Estructura histológica del cuerpo<br />
Hay tres capas: la mucosa se denomina endometrio, la muscular miometrio (muy importante) y la serosa se<br />
denomina perimetrio.<br />
Endometrio, ciclo<br />
El endometrio muestra cambios histológicos a lo largo de la vida:<br />
En la infancia tienen epitelio cúbico bajo con un estroma poco desarrollado con glándulas poco formadas e inactivas.<br />
En la época postmenopáusica tiene epitelio cúbico bajo, con un estroma más denso y frecuentemente con quistes<br />
de las glándulas.<br />
En la época fértil se puede dividir en dos estratos funcionales: la capa más superficial es la capa que va a<br />
experimentar cambios cíclicos (control hormonal9 y que en cada uno de estos cambios se desprende y tiene que<br />
volver a regenerarse y una capa basal que contacta con el miometrio y no experimenta cambios, sino que siempre se<br />
conserva y permite la regeneración de la capa superficial en cada ciclo.<br />
El epitelio es prismático simple recubierto por una cubierta proteica protectora. En él tenemos células ciliadas y<br />
células secretoras.<br />
Debajo encontramos una lámina propia que ocupa un grosor importante. En principio es bastante laxa y recuerda<br />
mucho al mesénquima. Tiene cambios cíclicos en su <strong>com</strong>posición, aunque siempre hay gran cantidad de glándulas<br />
tubulares simples. En principio las glándulas están restringidas a esta capa, pero en alguna patología <strong>com</strong>o la<br />
adenomiosis invaden el miometrio. La secreción propia, que varía según el momento del ciclo, permite cambios en<br />
los espermatozoides (capacitación) y también son esenciales para lograr la implantación de un blastocisto. Las<br />
porciones profundas de las glándulas están en la capa basal y son las que permiten la regeneración de las glándulas y<br />
el epitelio superficial.<br />
La capacitación es un proceso muy corto en nuestra especie (minutos) que consiste en una serie de cambios en lso<br />
espermatozoides para adaptarlos al canal reproductor femenino y fecundar un ovocito, la pérdida de la cubierta de<br />
glucoproteínas de la membrana plasmática del acrosoma, la disposición de determinadas moléculas de la membrana,<br />
lo que permite la activación de ciertos receptores para ligandos de la zona pelúcida y la hiperactivación de los<br />
espermatozoides para que naden más rápido.<br />
La lámina propia está muy vascularizada desde niveles más inferiores. A nivel del miometrio las arterias uterinas<br />
forman las arterias arqueadas y entre la zona del miometrio y la capa profunda del endometrio forman las arterias<br />
basales. Las ramas ascienden hasta alcanzar la capa funcional (arterias espirales) y son muy tortuosas; estas se<br />
caracterizan porque en un momento dado del ciclo sufren contracciones y dilataciones de forma rítmica. Después<br />
encontramos un plexo capilar superficial.<br />
Aunque se hable de fases, el ciclo es un proceso continuo que dura 28 días. Se suele dividir en tres periodos, la fase<br />
proliferativa o reparadora, secretora y menstrual. Es importante la correlación entre las fases del ovario y del útero.<br />
FASES CICLO MENSTRUAL<br />
La fase proliferativa ocurre a partir del día 4 y dura unos 10 días hasta el momento de la ovulación. En este periodo el<br />
endometrio mide <strong>com</strong>o máximo 5mm y sólo tiene capa basal. Responde a los estrógenos del ovario y se regenera la<br />
capa funcional. Se caracteriza por un gran número de mitosis a nivel del estroma y en los extremos desgarrados de<br />
las glándulas. Estas células epiteliales en proliferación regeneran las glándulas y el epitelio de revestimiento. Se<br />
consigue aumentar las glándulas en número y longitud de forma que acaban siendo rectas y estrechas. También<br />
prolifera la lámina propia, aumentando la sustancia fundamental y el colágeno por las células estromales, hasta que<br />
el estroma es metacromático. Durante los diez días también se recuperan los vasos superficiales que se han perdido,<br />
aunque el tercio superior aún no mostrará vasos. Las arterias espirales se regeneran pero tendrán una trayectoria no<br />
muy tortuosa. Las células epiteliales de las glándulas empiezan a acumular cantidades importantes de glucógeno en<br />
su citoplasma basal.<br />
Tras la ovulación <strong>com</strong>ienza la fase secretora (16-27). El endometrio crece hasta 6-7mm, dándose un notable aumento<br />
de grosor. Responde a la progesterona y estrógenos del cuerpo lúteo. Sigue habiendo mitosis pero a menor ritmo.<br />
Todo el endometrio está vascularizado, las glándulas se hacen más tortuosas (aserradas) y más grandes con una luz
Página137<br />
bastante amplia y secreción de moco viscoso acidófilo; además, aumenta la cantidad de secreción. En cuanto al<br />
estroma, se constata una formación de edema previo a la decidualización, que se produce si se da la implantación.<br />
En caso de que no haya implantación se da paso a la fase premenstrual o isquémica (1 día). En el estroma se produce<br />
una infiltración leucocitaria e involuciona y se retrae el endometrio. Las glándulas reducen su actividad y su secreción<br />
es bastante espesa. En la capa funcional se muestran alteraciones en la irrigación porque las arterias espirales se<br />
contraen y dilatan de forma intermitente durante unas horas. Hay momentos en los que el estrato superficial no<br />
tiene suficiente riego y entra en isquemia e hipoxia, y cesa la secreción glandular. Aumenta el número de leucocitos y<br />
el estroma se adelgaza y se vuelve menos edematoso.<br />
Las arterias finalmente se rompen en la fase menstrual (0-4). Esto supone una hemorragia de sangre arterial y<br />
venosa alteradas que llega a la luz y se elimina por la vagina; la menstruación es el sangrado del útero que conlleva la<br />
eliminación del endometrio preparado para una posible implantación. Hay pérdida de células epiteliales, conjuntivas<br />
y secreciones glandulares. Dura cuatro días y queda una mucosa desgarrada ya que se ha desprendido todo el<br />
estrato superficial. También se da una inhibición del proceso de coagulación. La capa basal no sufre estos cambios y<br />
se mantiene inalterada con todos sus vasos e irrigación perfectos.<br />
Cuando un blastocisto alcanza la cavidad uterina se implanta, lo que provoca cambios en el endometrio<br />
(decidualización). Este endometrio tiende a hacerse más hiperplásico, edematoso y secretor. Responde a la hCG del<br />
trofoblasto, que estimula el cuerpo lúteo a que siga secretando estrógenos y progesterona. En esta situación no<br />
habrá fase menstrual.<br />
Las células deciduales son células conjuntivas en las que se producen cambios metabólicos y morfológicos. Se hacen<br />
más grandes, son pálidas y su citoplasma tiene abundantes inclusiones lipídicas y de glucógeno. Se activan los<br />
receptores de estrógenos y progesterona. Tienen múltiples funciones: proporcionan los nutrientes necesarios para el<br />
desarrollo del embrión que se va a implantar, controlan el carácter invasivo del trofoblasto, intervienen<br />
decisivamente en evitar un rechazo inmunológico de los tejidos embrionarios y fetales por las células de la madre y<br />
segregan prostaglandinas, relaxina y una prostaglandina decidual similar a la hipofisiaria.<br />
También hay ciclos anovultarios en los que no hay fase secretora porque no ha habido ovulación ni cuerpo lúteo. La<br />
fase proliferativa es más amplia y continúa hasta que se elimina.<br />
La endometriosis es la implantación de tejido endometrial fuera de la pared uterina. Puede estar en la trompa,<br />
ovarios o en lugares más distantes <strong>com</strong>o intestino grueso, vejiga o a nivel pulmonar. Este tejido es capaz de<br />
permanecer en ellos y proliferar, así <strong>com</strong>o responder al control hormonal. Termina sangrando. En ocasiones puede<br />
quedar enmascarado porque las hemorragias cíclicas pueden quedar atrapadas. Otras veces sí se produce el<br />
sangrado (<strong>com</strong>o dismenorrea o dolor, menorragia o sangrado excesivo, o menometrorragia o sangrado entre ciclos) y<br />
en muchas ocasiones es difícil de detectar la endometriosis porque no hay un marcador específico. Otra patología<br />
importante es el carcinoma endometrial, el tumor maligno más frecuente del aparato genital femenino.<br />
Miometrio y perimetrio<br />
El miometrio es la capa de mayor grosor. Está formada por miocitos con una longitud de unos 20μm. Junto a ellos<br />
hay importante cantidad de conjuntivo en el que destacan las fibras elásticas. Se habla de tres estratos: superficial y<br />
profundo con fibras longitudinales y oblicuas, y una capa media circular. En esta capa media circular hay mayor<br />
presencia de vasos sanguíneos y linfáticos y se denomina capa vascular. Buena parte de las arterias tienen paredes<br />
muy gruesas que se pueden contraer fuertemente en la mujer que ha dado a luz para cortar hemorragias posparto<br />
peligrosas.<br />
Esta capa sufre importantes cambios en la mujer gestante. Hay un aumento muy considerable del número y tamaño<br />
de los miocitos, que se alargan hasta 600μm y en el tejido conjuntivo asociado al músculo <strong>com</strong>ienza a acumularse<br />
agua; de todos modos su grosor es menor porque el útero se está estirando. Esta capa responde a la relaxina<br />
segregada por el ovario y la placenta, inhibiendo las contracciones musculares. En cambio, la oxitocina<br />
(neurohipófisis) estimula las contracciones y es fundamental en el parto.<br />
Tras el alumbramiento hay reducción importante del tamaño de los miocitos, aunque no recobran el tamaño<br />
original. Su número tampoco se reduce drásticamente, por lo que su número es mayor que en la que nunca ha<br />
estado embarazada. Así, el útero tiene un tamaño mayor.
Página138<br />
Los miomas son tumores benignos del miometrio. Generalmente son redondeados y su crecimiento depende de los<br />
niveles hormonales. Es muy frecuente que estas células presenten alteraciones en el número de cromosomas y dan<br />
síntomas muy variables.<br />
Por último, el perimetrio es una adventicia asociada al peritoneo que se conoce <strong>com</strong>o mesotelio. Recubre el cuerpo<br />
del útero y posee un conjuntivo submesotelial.<br />
Estructura histológica del istmo y cérvix<br />
El istmo está en la parte superior del cuello. Su mucosa es más fina que la del cuerpo, pero equivalente, y sufre<br />
cambios cíclicos menos acentuados que a nivel del cuerpo. El epitelio es más bajo y tiene menos glándulas.<br />
En cuanto al cérvix, su luz está habitualmente ocluida por la presencia de un moco en toda la época fértil de la mujer,<br />
que presenta cambios cíclicos en su <strong>com</strong>posición.<br />
Ectocérvix y endocérvix<br />
La mucosa del cuello se denomina endocérvix. Tiene perfil irregular debido a numerosos pliegues transversos<br />
llamados plicae palmatae. Se aprecia la formación de unas hendiduras tubulares que parecen glándulas tubulares<br />
ramificadas. Es mucho más fino que el endometrio y apenas hay cambios cíclicos y lo normal es que no se desgarre ni<br />
sangre en el ciclo (no hay arterias helicinas en el estroma).<br />
El endotelio es prismático simple y se distinguen células ciliadas y secretoras: las ciliadas tienen cilios que baten hacia<br />
la vagina para eliminar el moco y las secretoras muestran cambios cíclicos regulados por estrógenos. Las células<br />
secretoras son más bajas en la fase proliferativa y sus núcleos son basales. Responden al aumento de estrógenos y se<br />
vuelven cada vez más altas a la vez que su citoplasma se llena de gránulos de mucígeno. En la fase secretora vierten<br />
la secreción y vuelven a ser células más bajas para repetir de forma cíclica los cambios. En los gránulos encontramos<br />
lisozima, mucinas y proteínas bactericidas, que actúa <strong>com</strong>o lubricante vaginal y <strong>com</strong>o tapón y barrera protectora<br />
frente a la entrada de patógenos ascendentes.<br />
En el epitelio, las zonas tubulares invaginadas del cérvix se corresponden con glándulas tubulares en forma de cripta.<br />
Esta zona se puede dilatar y su secreción quedar retenida formando los quistes de Naboth.<br />
El moco del canal cervical muestra cambios cíclicos en su viscosidad y pH. En los días u horas previas a la ovulación la<br />
sustancia se hace muy acuosa y clara y es muy abundante, con un pH alcalino que permite el acceso de los<br />
espermatozoides al tracto genital superior. Si se toma una muestra de la secreción en estos días es muy extensible y<br />
en un portaobjetos se ve que cristaliza en forma de hoja de helecho. En esta secreción hay altos niveles de sodio,<br />
potasio y cloro. Tras la ovulación, las células secretoras disminuyen su actividad y hay menos secreción, que se hace<br />
espesa y viscosa y el pH se hace ácido. En estos días tenemos una sustancia mucosa que sella el útero impidiendo la<br />
entrada de nuevos espermatozoides para proteger al blastocisto para que implante. En la mujer gestante el tapón<br />
cierra y protege el ambiente uterino hasta el inicio del parto cuando se elimina (trabajo del parto). Se muestra un<br />
sangrado señal de que el parto se está iniciando.<br />
Debajo del epitelio encontramos un estroma de conjuntivo bastante denso con miocitos lisos dispersos. Está<br />
vascularizado pero sin arterias helicinas.<br />
Debajo hay una capa muscular más fina que el miometrio. En este caso asociados a los miocitos hay conjuntivo denso<br />
fibroso con fibras colágenas y elásticas, que la hacen bastante dura, aunque se va a perder en el caso de embarazo<br />
justo al final de la gestación. Se constatan cambios de forma que se produce un reblandecimiento, lo que permite el<br />
ablandamiento y la dilatación del cuello para que pase la cabeza del feto. En la parte posterior del cérvix<br />
encontramos una serosa.<br />
La porción vaginal del cuello es la que sobresale a la vagina. Está revestida por mucosa lisa que se denomina<br />
ectocérvix. En esta porción vaginal el epitelio no es prismático simple sino plano estratificado no queratinizado y se<br />
continúa con el epitelio propio de la vagina. En este epitelio las células tienden a acumular glucógeno.<br />
La zona de transción se denomina unión escamosocolumnar del cérvix. En esta zona se observa el límite brusco del<br />
epitelio del endocérvix y el exocérvix y hay un aumento de las glándulas, por lo que es la zona más frecuente de<br />
quistes de Naboth ya que el epitelio estratificado cubre las zonas de salida de la secreción. Esta unión es<br />
clínicamente importante porque está implicada en algunas patologías. El punto exacto de esa unión varía y su<br />
localización es dependiente de las hormonas maternas. En la infancia coincide con el orificio externo, aunque estos
Página139<br />
epitelios responden a los estrógenos, por lo que a partir de la pubertad el endocérvix empieza a invadir la zona más<br />
allá del orificio externo. En esa situación se habla de que aparece un ectropión o una erosión cervical de modo que el<br />
epitelio de esta región es mucho más rojizo (eritroplasia, mancha roja). Responde a factores hormonales y va a más<br />
en cada embarazo. Una mujer que tenga varios embarazos irá invadiendo la porción vaginal.<br />
En esta zona se producen con mayor frecuencia alteraciones celulares, hay focos de displasia. Los más graves son los<br />
carcinomas in situ (neoplasia intraepitelial cervical, CIN), que quedan dentro del propio epitelio y puede progresar e<br />
infiltrar otros tejidos. Este tipo de alteraciones son muy fácilmente detectables mediante citología cérvico-vaginal. Es<br />
una prueba económica y de fácil acceso. También permite detectar la infección por el virus del papiloma humano<br />
(VPH). Hay una alta correlación de infección con este virus y que en la mujer se produzca un cáncer de cérvix.<br />
VAGINA<br />
Generalidades<br />
La vagina es el órgano copulador femenino. Es importante porque a la hora del parto forma el canal más inferior. Se<br />
extiende desde el orificio externo uterino al vestíbulo. Es tubular, fibromuscular y de unos 8-9cm y en reposo está<br />
aplastada y la entrada la cubre el himen.<br />
Estructura histológica<br />
La superficie de la mucosa muestra pliegues transversales o arrugas. El desarrollo de la mucosa depende de<br />
hormonas (estrógenos): la infancia y tras la menopausia el grosor es menor, y tras el climaterio está atrofiada. El<br />
moco que cubre la luz de la vagina no procede de glándulas propias de la vagina (no tiene) sino de las glándulas<br />
cervicales y de los genitales externos (glándulas de Bartholin). Contribuye en un aporte de agua y líquidos desde los<br />
capilares de la lámina propia normalmente en la excitación sexual.<br />
El epitelio es plano estratificado no queratinizado, <strong>com</strong>o el ectocérvix. Tiene distintas zonas:<br />
La más profunda tiene células altas con núcleos redondeados y citoplasma muy basófilo que cuando ascienden a la<br />
zona media van aplanándose, el citoplasma pierde basofilia, los núcleos <strong>com</strong>ienzan a retraerse y aparecen<br />
importantes inclusiones de glucógeno. Las más superficiales son muy planas, con núcleos picnóticos, citoplasma<br />
eosinófilo, inclusiones de glucógeno y queratohialina. Las células que se exfolian de forma constante serán objeto de<br />
transformación por parte de la flora vaginal. La bacteria más importante es el bacilo de Döderlein (Lactobacillus<br />
vaginalis), que forma ácido láctico a partir de glucógeno y contribuye al pH ácido de la vagina. Ese ácido es<br />
importante ya que protege frente a patógenos, pero no protege en las ETS.<br />
Cuando disminuye el ácido láctico, <strong>com</strong>o a partir del climaterio, hay tendencia al aumento en la frecuencia de las<br />
infecciones vaginales. Las infecciones ocurren fundamentalmente por estafilococos, hongos y protozoos, que<br />
necesitan un pH alcalino. Esto origina una inflamación no solo de la mucosa vaginal, sino de la piel de la vulva y se<br />
habla de vulvovaginitis. Es de fácil detección y tratamiento mediante un frotis vaginal. Para el tratamiento hay que<br />
restaurar el pH ácido y usar antibióticos y sulfonamidas.<br />
Debajo del epitelio hay una lámina propia con una zona superficial laxa rica en células y con importante cantidad de<br />
linfocitos y leucocitos que aumentan el número al aproximarnos al inicio del ciclo. En determinadas situaciones<br />
pueden formar nódulos importantes. La zona más profunda (submucosa) es un conjuntivo mucho más denso en el<br />
que las fibras colágenas y elásticas predominan. Es un conjuntivo bastante vascularizado con venas abundantes de<br />
paredes finas que permiten una cierta erección en la excitación sexual. El número de terminaciones nerviosas es muy<br />
pobre, fundamentalmente de dolor.<br />
Tiene una capa muscular importante (CI y LE). El externo es más grueso y se desarrolla una zona inferior que<br />
funciona <strong>com</strong>o esfínter, el músculo bulboesponjoso.<br />
Por último hay una adventicia gruesa. El estrato más interno está formado por conjuntivo muy denso con fibras<br />
elásticas que otorgan a la vagina resistencia y elasticidad. La parte más externa tiene un conjuntivo mucho más laxo.<br />
Los dos estratos están muy vascularizados con sangre y linfa.
Página140<br />
Cambios cíclicos<br />
El epitelio es dependiente de estrógenos y muestra cambios cíclicos. En la fase proliferativa el número de capas del<br />
epitelio estratificado va creciendo y las células se hacen más eosinófilas, almacenan más glucógeno y al final de la<br />
fase hay niveles elevados de glucógeno y tasa de descamación importante.<br />
En la ovulación el epitelio tiene un grosor máximo, las células se han cargado de gránulos de queratohialina y los<br />
núcleos de las células superficiales son picnóticos.<br />
Tras la ovulación (fase secretora) el número de capas disminuye, así <strong>com</strong>o el contenido de glucógeno. Hay mayor<br />
proporción de células basófilas y mayor número de leucocitos y bacterias en la vagina.<br />
Si se realizan citologías vaginales se pueden obtener distintos resultados que se correlacionan con las diferentes<br />
fases. En la ovulación debe haber predominio de células eosinófilas, en la fase scretora las basófilas y antes de la<br />
ovulación el número de ambas es el mismo.<br />
Las citologías se realizan mediante el método Papanicolaou. Se tiñen con hematoxilina, naranja G y eosina azur. Se<br />
usa para frotis vaginales y de cérvix. Se distinguen patologías, cambios hormonales y situaciones patológicas<br />
referentes a la flora vaginal. Los Pap cervicovaginales se usan también para diagnosticar precozmente el cáncer de<br />
cuello de útero y el de carcinoma endometrial. Las células cancerosas se detectan fácilmente y se puede determinar<br />
su lugar de origen.<br />
PLACENTA<br />
Implantación<br />
Tras la ovulación y fecundación se inicia la segmentación. El blastocisto desciende y en el día 6 llega al útero. Se<br />
produce el fenómeno de la implantación en la parte superior de la pared posterior del cuerpo uterino. En la primera<br />
fase se produce una adhesión inestable. Luego viene una penetración en el endometrio. Simultáneamente al proceso<br />
ocurre la decidualización (transformación del endometrio en decidua).<br />
Fases<br />
En la adhesión (inicial e inestable) el blastocisto contacta con la superficie del endometrio. Las células muestran en<br />
su membrana el HB-EFG, que va a resultar el ligando de un receptor expresado por las células del trofoblasto<br />
(receptor para HB-EFG). Sólo lo expresan en determinados días, igual que el ligando. Cuando el blastocisto llega el<br />
receptor se une al ligando y se produce un contacto inestable con él y el heparán sulfato. Las células trofoblásticas<br />
<strong>com</strong>ienzan a desarrollar microvellosidades que crecen para meterse entre los pinópodos de las células del<br />
endometrio.<br />
En la segunda fase o penetración (estable) el trofoblasto va a penetrar en el endometrio. Muestra una diferenciación<br />
en una capa interna que se divide activamente (citotrofoblasto), y una capa externa o sincitiotrofoblasto, que es una<br />
masa multinuclear.<br />
El sincitiotrofoblasto erosiona el endometrio, lo que permite que el blastocisto avance y penetre. En esta<br />
penetración se habla de invasión intersticial, es decir, en el avance del blastocito en el endometrio juegan un papel<br />
importante enzimas proteolíticas que erosionan el endometrio y se segregan por el sincitiotrofoblasto. En la invasión<br />
endovascular se rompen los vasos del endometrio, por lo que las arterias espirales se rompen y hay una apertura de<br />
la circulación. Por último, en la invasión por fusión los núcleos de las células del estroma materno pasan a ser<br />
incorporados en el sincitiotrofoblasto en su erosión. Como consecuencia de la rotura de los vasos maternos la sangre<br />
se extravasa y quedan lagunas de sangre materna en el sincitio. Conforme avanza se forma una auténtica red lacunar<br />
con lagunas conectadas entre ellas. Se establece una circulación úteroplacentaria primitiva.
Página141<br />
Vellosidades coriónicas<br />
Seis días después del inicio de la implantación (días 11-13) el mesodermo extraembrionario induce la proliferación<br />
del citotrofoblasto de forma que sus células se meten dentro del sincitio. Estas proliferaciones se llaman vellosidades<br />
primarias. Otros seis días después (día 16), el mesodermo que ha inducido los cordones, además de seguir<br />
induciendo la proliferación se mete en las vellosidades y se habla de vellosidades secundarias con mesodermo en el<br />
centro. En el día 20 se produce una diferenciación celular en el mesodermo y se forman capilares y células<br />
sanguíneas. Así, las vellosidades terciarias son en las que dentro del mesodermo de la vellosidad hay vasos.<br />
Estas células de las vellosidades siguen proliferando y llega un momento en que penetran tan profundamente que<br />
sobrepasan el sincitio y llegan al endometrio. En ese punto no avanzan sino que proliferan hacia los lados formando<br />
un límite que rodea todo el blastocisto, la corteza citotrofoblástica. Esta masa tiene un límite que permite que el<br />
blastocisto se fije al endometrio. Cuando una vellosidad se ancla al endometrio se denomina vellosidad de anclaje.<br />
Las más cortas laterales son las vellosidades libres o terminales. Están flotando en el espacio intervelloso, lleno de<br />
sangre (red lacunar).<br />
El día 21 el corazón <strong>com</strong>ienza a latir. Esto permite el bombeo de sangre por todo el embrión. La circulación<br />
embrionaria alcanza las vellosidades y es un circuito cerrado. La sangre de las vellosidades terminales va a<br />
intercambiar gases y nutrientes con la sangre materna de las lagunas. La sangre materna ha penetrado en las lagunas<br />
por diferencia de presiones. Una vez se produce el intercambio de gases y nutrientes, la sangre materna retorna a la<br />
circulación cerrada por las venas uterinas. La sangre del embrión oxigenada y cargada de nutrientes retorna al<br />
embrión por la vena umbilical. Las arterias umbilicales llevan la sangre sin oxígeno hacia las vellosidades terminales.<br />
Decidualización<br />
En los días 2-7 en el estroma <strong>com</strong>ienza la fase primaria de decidualización. Se forma una zona decidual primaria en<br />
la que las células tienen aspecto epitelioide, alta tasa de proliferación y segregan proteínas <strong>com</strong>o fibronectina,<br />
laminina, entactina y colágeno I, III, IV y V. Además, expresan proteínas morfogenéticas óseas 2 y 7, FGF, Wnt4 y<br />
proteína Hedgehog. Simultáneamente las células del epitelio del endometrio reducen la cantidad de desmosomas<br />
que las unen. Empiezan los primeros signos de apoptosis. En la decidua los vasos aumentan su permeabilidad en el<br />
lecho de implantación.<br />
Durante la penetración da una fase secundaria de decidualización. Se detecta en las células un cambio en el<br />
metabolismo que permite que se expresen otras proteínas, <strong>com</strong>o las metaloproteinasas (MMP), inhibidores tisulares<br />
de las metaloproteinasas, inhibidores de la remodelación de la matriz decidual en presencia de prostaglandina 2 y<br />
activadores del plasminógeno.<br />
La decidua es el endometrio del útero grávido. Se denomina así porque en el parto se elimina entera menos la capa<br />
basal. Entre sus funciones están la nutrición y protección del embrión. Las células se cargan de lípidos y glucógeno.<br />
Esta capa permite controlar la invasión del trofoblasto, concretamente del sincitio. Además, evita el rechazo de la<br />
madre respecto del embrión, que equivale a un alotransplante. El sincitio es una masa embrionaria que no expresa<br />
nunca MHC II. Además, las zonas más limítrofes tampoco tienen MHC I. Sin embargo, sí expresan moléculas que<br />
pueden provocar rechazo, las C, E y G de los HLA. Esto podría provocar un ataque de los NK. Sin embargo, la forma G<br />
de la HLA se une a unos receptores símil de Ig de forma que se evita el ataque de las células NK. Las deciduales<br />
sintetizan y segregan prostaglandinas e inmunosupresores que inhiben las células NK. Además, los leucocitos<br />
infiltrados segregan IL2 para evitar el rechazo materno del embrión. Las células deciduales también expresan<br />
citosinas con efecto depresor de los linfocitos T. Además, en esta región la función de los LT está dificultada debido a<br />
un déficit en triptófano. También se cree que los LT durante la gestación tienen menor número de receptores. Por<br />
último la hCG, estrógenos y corticosteroides tienen efecto inmunosupresor.<br />
La decidua tiene función endocrina. Segrega prolactina decidual, similar a la conocida y que ejerce efecto trófico<br />
sobre el cuerpo lúteo. También tiene relaxina y prostaglandinas, que inhiben el crecimiento y mitosis de las células<br />
del endometrio.
Página142<br />
Tipos de decidua<br />
La decidua basal es la porción de la decidua inmediatamente por debajo del blastocisto implantado. Tiene<br />
vellosidades. Es el <strong>com</strong>ponente materno de la placenta y se fija a la placa coriónica. La decidua capsular es la parte<br />
de la decidua que corresponde a la parte sobre el blastocisto implantado y el corion. También presenta vellosidades.<br />
El resto del endometrio forma la decidua parietal.<br />
Inicialmente se forman vellosidades coriónicas en todo el saco corióncio. A partir del segundo mes de vida las de la<br />
decidua capsular degeneran y sólo quedan las de la decidua basal. El corion asociado a la decidua capsular se<br />
denomina corion liso o leve. El que sí presenta vellosidades, asociado a la decidua basal, se denomina corion<br />
frondoso y es el <strong>com</strong>ponente fetal de la placenta. Con el desarrollo, la decidua capsular acaba degenerando y el<br />
corion leve se fusiona con la decidua parietal de forma que la luz uterina desaparece por el crecimiento del feto.<br />
Placenta: características anatómicas e histológicas<br />
La placenta es un órgano en forma de disco circular que aumenta de grosor por el crecimiento de las vellosidades.<br />
Alcanza su desarrollo máximo al quinto mes de vida. Pesa 0.5kg y mide 2.5cm de grosor.<br />
Anatómicamente, la cara fetal es una cara lisa muy brillante por el amnios que la cubre. En el centro tendríamos la<br />
inserción del cordón umbilical. La cara materna no es homogénea sino que está dividida en 10-20 unidades en forma<br />
de cuenco llamadas lóbulos o placentomos. Cada lóbulo se separa del vecino por tabiques derivados de la decidua<br />
basal que se extienden hacia la placa coriónica.<br />
Histológicamente presenta una placa coriónica en la cara fetal y una placa basal en la cara materna. Entre medias<br />
están las vellosidades coriónicas en el espacio intervelloso. La placa coriónica es el mesodermo apoyado sobre el<br />
amnios. La placa basal es la decidua basal más la capa basal del endometrio. Desde ahí salen los tabiques que dividen<br />
la placenta en placentomos o lóbulos. Las vellosidades coriónicas están bañadas en sangre materna lacunar.<br />
El amnios es epitelio cúbico simple con frecuentes metaplasias. Bajo él tenemos un conjuntivo. La zona más próxima<br />
carece de vasos y luego tenemos un mesodermo en continuidad con el conjuntivo del corion. Ahí sí hay grandes<br />
vasos, ramas de las arterias y vena umbilicales. Como límite de la placa coriónica encontramos un sincitio<br />
trofoblástico continuo que contacta con la sangre del espacio intervelloso. Los tabiques placentarios tienen distinta<br />
longitud y altura y son siempre in<strong>com</strong>pletos. Nunca llegan a alcanzar la placa coriónica.<br />
La placa basal es la placa formada por el citotrofoblasto continuo que delimita y ancla el blastocisto al endometrio<br />
(corteza citotrofoblástica). Además tenemos la decidua basal con células deciduales, y en menor medida fibroblastos<br />
y linfocitos. Hay una matriz muy laxa con colágeno IV, laminina, fibronectina y heparán sulfato. Es frecuente que las<br />
células del citotrofoblasto penetren y se encuentren aisladas <strong>com</strong>o células X, muy basófilas.<br />
Dentro de cada lóbulo hay un número variable de vellosidades de anclaje (hasta 10). Un cotiledón es el espacio del<br />
lóbulo en el que sólo hay una vellosidad de anclaje con sus ramificaciones.<br />
El fibrinoide es un depósito que recuerda a la lámina basal, muy eosinófilo. Tiene fibrinas, inmunoglobulinas y<br />
algunas células trofoblásticas muertas. Crece según avanza el desarrollo y puede incluso llegar a casi reemplazar<br />
vellosidades enteras. En la placa coriónica encontramos los fibrinoides de Langhans. En la superficie basal y<br />
vellosidades tenemos fibrinoide de Rohr y el de Nitabuch está cerca de la capa basal del endometrio.<br />
Hay dos tipos de fibrinoide. El tipo fibrina se origina cuando hay algún tipo de lesión del sincitiotrofoblasto, que<br />
origina coagulación de la sangre materna. A partir de los coágulos (con fibrina) se formaría el fibrinoide. Tiene<br />
función de barrera y protección. El fibrinoide tipo matriz es una secreción de citotrofoblasto.
Página143<br />
Membrana placentaria<br />
La vellosidad terciaria es la unidad funcional de la placenta. Si se extendieran tendríamos 15m 2 . Hasta el quinto mes<br />
las vellosidades están formadas por un eje de mesodermo con células mesenquimales que son las que segregan la<br />
matriz. En su momento diferenciarán a fibroblastos y a células son las de Hofbauer, fagocíticas. También hay<br />
capilares. Luego encontramos una lámina basal y un epitelio, el citotrofoblasto. Es una capa de células epiteliales<br />
unidas por desmosomas entre ellas y al sincitio y que sufren muchas mitosis. El sincitio está en contacto con la<br />
sangre materna y puede dar microvellosidades al espacio intervelloso.<br />
A partir del cuarto mes la estructura de la microvellosidad se simplifica y hay menos distancia entre los capilares<br />
fetales y la sangre materna. El sincitio se va adelgazando. Se agrupan los núcleos para que ocupen menos. Los<br />
núcleos agrupados se denominan brotes sincitiales o nudos y sobresalen hacia la sangre materna. Con el paso del<br />
tiempo los brotes se desgajan y se arrastran por la sangre materna. Acaban degradándose en los vasos pulmonares.<br />
El citotrofoblasto desaparece en buena parte. Algunas células quedan justo debajo del sincitio, pero ya no es una<br />
capa continua. Por último, la cantidad de mesodermo disminuye pero aumenta mucho el número y tamaño de<br />
capilares, que tienden a situarse inmediatamente por debajo del sincitio. El espacio que queda entre las dos sangres<br />
disminuye. Esta barrera se conoce <strong>com</strong>o membrana placentaria. Así, la membrana placentaria es el tejido fetal que<br />
separa la sangre fetal de la materna.<br />
Funciones de la placenta<br />
Permite la nutrición del embrión y feto, y la excreción de los productos del metabolismo. Permite el intercambio de<br />
gases entre la sangre materna y fetal, la transferencia de inmunoglobulinas de la madre al feto. También actúa <strong>com</strong>o<br />
órgano endocrino.<br />
Con la nutrición y la excreción se relacionan elementos de pequeño tamaño <strong>com</strong>o agua, iones, vitaminas, glucosa,<br />
ácidos grasos y aminoácidos. No la pasan moléculas de gran tamaño <strong>com</strong>o péptidos y proteínas, ni triglicéridos. Se<br />
eliminan bilirrubina libre y urea. Hay transporte activo y pasivo.<br />
Los gases se intercambian por difusión simple para eliminar el CO 2 del feto. También la atraviesan el oxígeno y el<br />
monóxido de carbono. Permite la oxigenación de la sangre fetal.<br />
Las IgG pasan la membrana placentaria y le confieren al feto inmunidad pasiva.<br />
Como órgano endocrino sintetiza progesterona, en especial el sincitiotrofoblasto. Sin embargo no pasa directamente<br />
a estrógenos sino que sale a la circulación fetal y llega hasta la glándula suprarrenal feta. En la corteza es<br />
transformada en andrógenos (DHEA y DHEAS). Los andrógenos retornan a las vellosidades terciarias y a partir de<br />
ellos el sincitio sintetiza estrógenos, esenciales en el mantenimiento de la gestación. La unidad fetoplacentaria es la<br />
colaboración entre la corteza suprarrenal fetal y la placenta.
Página144<br />
El sincitio también sintetiza hCS (lactógeno placentario). Estimula el crecimiento del feto y tras el parto la<br />
lactogénesis. Es la responsable de la diabetes de un importante número de mujeres mientras están embarazadas.<br />
Después del parto dejan de ser diabéticas.<br />
La hCG es la que da resultado positivo en las pruebas de embarazo y se sintetiza ya en el día 6. Estimula la secreción<br />
de T4 y es esencial para mantener el cuerpo lúteo al principio. Permite el desplazamiento lúteo-placentario.<br />
El citotrofoblasto sintetiza el IGF-I y II. Actúan sobre el propio citotrofoblasto estimulando las mitosis, el crecimiento<br />
y la diferenciación de la capa.<br />
El EGF se sintetiza al principio por el citotrofoblasto. También estimula las mitosis de las células del trofoblasto.<br />
Después quien se encarga de sintetizarlo es el sincitiotrofoblasto para mantener la función del trofoblasto<br />
diferenciado.<br />
La relaxina facilita el reblandecimiento del cuello uterino y sus ligamentos antes del parto. La sintetizan las células<br />
deciduales.<br />
Por último, la leptina se detecta en grandes cantidades al final del embarazo y la sintetiza el sincitio. Controla las<br />
reservas de nutrientes de la madre para el feto y su transporte a través de la placenta.<br />
MAMA<br />
La mama es un órgano exclusivo de los mamíferos, diseñado para poder alimentar y nutrir a las crías, que son muy<br />
inmaduras. Son en realidad glándulas cutáneas modificadas.<br />
Embriología<br />
Hay dos hormonas que juegan un papel fundamental en el desarrollo de esta glándula mamaria: el lactógeno<br />
placentario y los estrógenos, que actúan sobre el epitelio y el mesénquima.<br />
La formación de la mama presenta dos fases: la formación del pezón y la formación de la glándula mamaria.<br />
En la sexta semana del desarrollo embrionario, se produce una acumulación de células epiteliales que forman una<br />
depresión, el pezón invertido. Estas células se agrupan en dos líneas paralelas a lo largo de las cuales encontramos<br />
estas acumulaciones. Estas zonas engrosadas son las líneas mamarias, y se extienden en la zona ventral, desde la<br />
axila hasta la zona inguinal.<br />
Cada una de estas acumulaciones conforma una yema o botón epidérmico primario, que van profundizando y<br />
ramificándose, de modo que de cada yema primaria se forman de 15 a 20 cordones epiteliales macizos, que<br />
conforman las yemas secundarias.<br />
Estos cordones <strong>com</strong>ienzan a ahuecarse y finalmente se observa que en los cordones se desarrollan los alveolos. Cada<br />
cordón ahuecado es entonces un conducto galactóforo.<br />
Al final del desarrollo mamario solo quedan dos yemas primarias a nivel pectoral, que corresponden a los pezones.<br />
Pueden suceder polimastias o politelias, que a veces no se descubren hasta la pubertad.<br />
Tras el nacimiento, el pezón invertido protruye y la areola se eleva según se desarrollan las glándulas en su interior.<br />
En el recién nacido, las mamas son de tamaño mayor que en la infancia, lo que se correlaciona con una posible<br />
secreción grasa con α-lactalbúmina (leche de brujas), que responde al efecto que ejercen los estrógenos<br />
placentarios.<br />
Al poco tiempo, esos estrógenos desaparecen y las mamas involucionan (sobre todo el parénquima). En varón, la<br />
mama casi no se desarrolla, y en mujeres el desarrollo se produce por acción de varias hormonas.<br />
El aumento de estrógenos en la pubertad, junto con la prolactina, generan un estímulo en el conjuntivo y en el<br />
parénquima epitelial. Se observan una serie de cambios cíclicos debidos a la progesterona. Una mama solo alcanza<br />
su desarrollo máximo en una mujer gestante y durante la lactancia.
Página145<br />
La ramificación de los conductos es dependiente de factores segregados por adipocitos.<br />
Las células epiteliales producen además células mioepiteliales que migran a la periferia.<br />
El mesodermo también responde a estímulos del epitelio. Las yemas segregan la PTH-RP, responsable de que el<br />
mesénquima de alrededor se diferencie en conjuntivo, tejido adiposo y mésculo liso. Existen datos que muestran que<br />
una mutación en el gen que codifica para PTH-RP puede bloquear el desarrollo mamario porque impide que la yema<br />
profundice y se desarrolle.<br />
La interacción entre el epitelio y el mesénquima es fundamental en el desarrollo de la mama. Las células epiteliales<br />
proliferan inducidas por el mesénquima que las rodea.<br />
Cada yema primaria origina una mama, mientras que cada botón secundario origina un lóbulo mamario.<br />
En el caso de un feto varón, este sistema de conductos involuciona <strong>com</strong>o consecuencia de los altos niveles de<br />
testosterona. La gine<strong>com</strong>astia es un trastorno en que en el varón se desarrollan las mamas tanto por proliferación y<br />
ramificación del sistema de conductos, y por aumento del tejido fibrocolágeno periductal. Se produce cuando faltan<br />
receptores de testosterona (síndrome de insensibilidad los andrógenos), en caso de desequilibrio entre andrógenos<br />
testiculares y estrógenos de corteza suprarrenal (cirrosis hepática, tratamientos de cáncer de próstata) y en el<br />
síndrome de Klinefelter (47, XXY).<br />
Estructura Histológica<br />
Pezón o papila mamaria<br />
El pezón y la areola están recubiertos de una epidermis muy arrugada, un epitelio plano estratificado queratinizado,<br />
y papilas dérmicas muy abundantes y profundas, haciendo que haya mucha vascularización y por ello tengan<br />
tonalidad rosácea.<br />
En pubertad y gestación, la papila y la areola se hacen más prominentes y pigmentados, ya que los estrógenos<br />
estimulan a melanocitos y hay una melanogénesis mayor. Encontramos una gran cantidad de vasos linfáticos<br />
Estos haces musculares se distribuyen de manera longitudinal o radial.<br />
Encontramos inervación sensitiva que actúa en el reflujo de erección de la papila.
Página146<br />
La papila tiene unos orificios dispuestos anularmente, cada uno de los cuales da paso a un conducto galactóforo.<br />
Cada conducto drena un lóbulo mamario. Los orificios están tapados con queratina.<br />
Areola<br />
En la areola mamaria encontramos varios tupos de glándulas: sebáceas no asociadas a folículos, sudoríparas y de<br />
Montgomery (una mezcla entre sudoríparas y mamarias).<br />
Están en la areola y son muy prominentes, de manera que se puede observar donde se sitúan. Presentan una<br />
secreción lubricante de tipo graso que aumenta durante la lactancia.<br />
Lóbulos mamarios<br />
Cada botón primario corresponde a una mama, y cada botón secundario a un lóbulo de dicha mama. Cada mama<br />
tiene entre 15 y 20 lóbulos.<br />
Los lóbulos mamarios se disponen radialmente, separados por tejido conjuntivo denso interlobular, con abundantes<br />
adipocitos.<br />
Los lóbulos amaraos están unidos a la zona profunda de la dermis mediante los ligamentos suspensorios o de<br />
Cooper, que se anclan en la fascia del musculo pectoral.<br />
Cuando la mama se desarrolla, el ligamento se distiende. Cuando la mama reduce su volumen, el ligamento no se<br />
retrae.<br />
Cada lóbulo tiene una glándula tubuloalveolar ramificada, que en origen era una sudorípara y ha sido ramificada. Así,<br />
cada mama tiene entre 15 y 20 glándulas<br />
Lobulillos mamarios<br />
Un lobulillo es una parte de la glándula drenada por un conducto. Entre los lobulillos hay tejido conjuntivo<br />
interlobulillar, denso. Este conjuntivo puede entrecruzarse con el conjuntivo de otros lóbulos, y debe ser tenido en<br />
cuenta en cirugías.<br />
Mama Inactiva<br />
Estroma<br />
En la mama inactiva, el estroma es muy abundante. Se <strong>com</strong>pone de conjuntivo interlobular, interlobulillar e<br />
intralobulillar. Este último es un conjuntivo más laxo, sin adipocitos, con escasas células inmunitarias y muchos<br />
capilares.<br />
Parénquima<br />
El parénquima en la mama inactiva está muy reducido. Si esta dsarrollado el sistema de vías (dpte de estrógenos)<br />
pero no el secretor (dpte de progesterona)<br />
Las vías excretoras de la mama son el conducto galactóforo y el seno lactífero, a partil del cual se produce una<br />
ramificación dicotómica, dando conductos terminales, cada uno de los cuales drena un lobulillo. De estos conductos<br />
se da una ramificación irregular que forma los conductillos intralobulillares, entre los que encontramos tejido<br />
conjuntivo intralobulillar.<br />
Conducto galactóforo: está tapizado por un epitelio estratificado cúbico que cuando alcanza la superficie se<br />
convierte en epitelio estratificado plano. Tiene una longitud de unos 3 mm.
Página147<br />
Seno lactífero: es una dilatación del conducto galactóforo, y está tapizado por epitelio estratificado cúbico.<br />
Constituye un reservorio de leche.<br />
A partir del seno hay un sistema de vías llamados conductos terminales o intralobulillares que se ramifican de forma<br />
dicotómica, y su epitelio pasa de ser estratificado cúbico a ser simple primatico y finalmente simple cúbico. A lo largo<br />
de todas estas vías encontramos células mioepiteliales (IHQ: Actina, p63).<br />
Cada lobulillo drena por un conducto terminal. Dentro de cada lobulillo encontramos los conductos intralobulilalres,<br />
que terminan en fondo de saco en el caso de la mama inactiva.<br />
Unidades secretoras: cambios en el ciclo menstrual<br />
La unidad secretora es el alveolo, y en una mama inactiva no existen, si bien en la pubertad hay cambios cíclicos en el<br />
final de los conductillos intralobulilalres. Coincidiendo con los picos de progesterona o de estrógenos, en el día 16 del<br />
ciclo menstrual, los extremos de los conductos se dividen y aumentan de tamaño y tienen signos de células<br />
secretoras.<br />
Estos conductillos empiezan a dilatarse, de modo que hay una pequeña luz y en el conjuntivo intralobullilar se<br />
empieza a acumular líquido y GAG. Aumenta además el tamaño de los lobulillos.<br />
Estos cambios que se detectan cíclicamente no afectan a toda la mama. Algunos lobulillos se hipertrofian y las yemas<br />
se desarrollan. La mama aumenta ligeramente de tamaño, provocando molestias.<br />
Si en ese ciclo los niveles de progesterona caen por falta de fecundación, el proceso queda parado hasta el siguiente<br />
ciclo menstrual, cuando se hipertrofiarán otros conductos que no lo hicieron en el primer ciclo, y los antiguos<br />
conductos hipertrofiados involucionarán.<br />
Si hay fecundación, la progesterona es responsable del desarrollo amplio y generalizado de las yemas en toda la<br />
mama.<br />
Mama en el embarazo<br />
En el embarazo, la mama sufre una serie de cambios para preparar la lactancia<br />
Estos cambios se ven favorecidos por el lactógeno placentario, estrógenos, prolactina y progesterona.<br />
Se desarrollan los extremos de los conductos, disminuyendo la cantidad de conjuntivo y aumentando la de tejido<br />
adiposo. Se observa asimismo la infiltración de células inmunitarias<br />
El parénquima glandular <strong>com</strong>ienza a crecer. Se siguen ramificando los conductos y aparecen los alveolos.<br />
En el primer trimestre, aumenta la vascularización, y con ello la pigmentación, de la papila y la areola. Los<br />
conductillos terminales proliferan por hiperplasia y se observan en ellos signos de células secretoras.<br />
En el segundo trimestre estas células son ya claramente células secretoras. Se observan luces abiertas al final de los<br />
conductillos Aumenta el número de células inflamatorias. El tejido conjuntivo intralobulillar es el que más prolifera.<br />
En la última parte del embarazo, las células secretoras son muy activas. Se produce así una secreción abundante que<br />
se acumula en alveolos y conductos. Los alveolos se encuentran en este momento en su desarrollo máximo.
Página148<br />
Las células secretoras son eosinófilas, con un RER abundante, un aparato de Golgi en posición supranuclear,<br />
mitocondrias, REL, vesículas de secreción e inclusiones lipídicas.<br />
La leche es el producto de secreción de la mama. Algunos <strong>com</strong>ponentes se secretan por merocrinia y los elementos<br />
grasos por apocrinia; las moléculas grasas forman lóbulos de gran tamaño en la zona apical del citoplasma. Las<br />
células plasmáticas del conjuntivo intralobulillar secretan Ig, que se liberan al epitelio alveolar y por transcitosis<br />
llegan a la leche a través de las células epiteliales.<br />
Mama en la lactancia<br />
Después del parto, se produce una caída brusca de los niveles de estrógenos y progesterona. La prolactina, que será<br />
la nueva encargada de estimular la síntesis de leche, requiere días para mostrar su efecto lactogénico. La GH, la PTH<br />
y glucocorticoides también tienen efecto lactogénico. La oxitocina, por su parte, se encarga de la expulsión de leche.<br />
Durante la lactancia se mantiene un desarrollo máximo de los alveolos, que presentan un epitelio cúbico con células<br />
mioepiteliales. Inicialmente segregan calostro y después leche madura.<br />
Los alveolos no están en el mismo estado funcional en toda la mama. Hay zonas con alveolos dilatados llenos de<br />
leche, con epitelio bajo. Otras zonas tienen alveolos de luz estrecha y un epitelio alto, sin reservorio de leche.<br />
Las células mioepiteliales tienen receptores para oxitocina y son muy sensibles a ella, de modo que la oxitocina<br />
promueve la expulsión de la leche.<br />
Calostro<br />
Es una secreción láctea amarillenta que se produce en los primeros días de lactancia. Es muy alcalina, rica en<br />
proteínas, también contiene grasa. Contiene sodio y cloro, así <strong>com</strong>o vitamina A. Contiene además IgA, que da<br />
inmunidad pasiva al recién nacido.<br />
Leche madura<br />
Es un alimento específico para cada especie de mamífero. Se segrega aproximadamente 1100 ml al día (mas de 2 l si<br />
hablamos de parto múltiple).<br />
Contiene elementos nutritivos, inmunológicos, protectores y estimuladores del crecimiento.<br />
· Proteínas: caseína, -lactalbúmina, PTH-RP, lactoferrina, lisozima<br />
· Lípidos: triglicéridos, colesterol<br />
· Azúcares: lactosa, que atrae agua hacia la vesícula secretora<br />
· IgA dimérica: 5 meses después del parto la leche aún tiene muchas Igs
Página149<br />
CICLO SECRETOR<br />
1. Secreción<br />
Partimos de una zona con alveolos pequeños, una luz pequeña y vacía, y un epitelio alto y plegado. Son células de<br />
citoplasma eosinófilo y núcleo basal.<br />
Los alveolos <strong>com</strong>ienzan a crecer, el epitelio disminuye haciéndose cúbico bajo y se estría. Se desarrollan las organelas<br />
y se almacena la secreción en las luces; la luz se llena de material granulado eosinófilo en el que flota grandes gotas<br />
lipídicas.<br />
Para que la leche que se ha formado por acción de la prolactina se expulse, se precisa de oxitocina.<br />
Tras el parto caen los niveles de prolactina, así que no se puede producir la secreción espontáneamente, por lo que<br />
la succión del bebe es fundamental, ya que estimula receptores de la papila, generando impulsos que llegan al<br />
hipotálamo, el cual deja de segregar un factor que promueve la inhibición de la prolactina, lo que hace que la<br />
adenohipófisis segregue prolactina en grandes cantidades, lo que permite iniciar el ciclo de secreción.<br />
La neurohipófisis <strong>com</strong>ienza a generar oxitocina, ya que si la leche no se expulsa, se detiene el ciclo.<br />
La producción y secreción de leche es continua, mientras que la expulsión es episódica y precisa de la participación<br />
del bebé. Amamantar constantemente hace que la lactancia se prolongue 2 o 3 años.<br />
2. Involución de la mama<br />
Si la lactancia se interrumpe, se produce en la mama una <strong>com</strong>presión de los vasos sanguíneos. La sangre que fluye<br />
por estos vasos no llega a las células mioepiteliales, por lo que tampoco llega la oxitocina y no se produce la<br />
expulsión de leche aunque la leche se sigue produciendo. Esta leche que es almacenada y no es expulsada acaba<br />
siendo reabsorbida por el epitelio alveolar, que empieza a entrar en apoptosis. Se sintetizan una serie de enzimas<br />
que están destinadas a degradar las células epiteliales, produciéndose un colapso alveolar. La fagocitosis de estos<br />
elementos epiteliales en degeneración corre a cargo de macrófagos y no se trata de una reacción inflamatoria. Como<br />
consecuencia, disminuye el volumen del parénquima glandular y aumenta el del estroma, es decir, la mama retorna<br />
al estado de reposo similar al de la mama inactiva.<br />
La prolactina tiene, además de la estimulación de la síntesis de leche, otra serie de efectos. La secreción masiva de<br />
prolactina inhibe la secreción de la hormona liberadora de gonadotropina y de LH, lo que hace que no se produzca<br />
ovulación, de ahí que la lactancia sea un método anticonceptivo.<br />
Cuando una mama involuciona tras la lactancia, el aspecto que se recobra es muy similar al de la mama inactiva, pero<br />
hay ciertas diferencias: restos que nos revelan que ha habido una gestación y una lactancia. En primer lugar, los<br />
alveolos no desaparecen del todo, si bien la mayoría vuelven a ser conductos que acaban en fondos de saco.<br />
Durante un tiempo prolongado, se encuentran restos de la secreción en los conductos.<br />
Mama en la menopausia<br />
Cuando la mujer llega a la menopausia, las mamas sufren un proceso de involución que hace que vuelvan a un<br />
estado prepuberal. Hay pues una atrofia significativa del parénquima. El estroma también se atrofia y pierde<br />
densidad celular, así <strong>com</strong>o fibras colágenas y elásticas.<br />
Patología: mama del varón<br />
Estructura fibroadiposa con el <strong>com</strong>ponente ductal de las glándulas, poco desarrollado.
Página150<br />
En ambos sexos, la mama presenta una estructura inicial igual hasta la pubertad, cuando las hormonas sexuales<br />
femeninas (estrógenos, prolactina, progesterona, somatotropina y glucocorticoides) permiten el desarrollo mamario,<br />
mientras que en el hombre continuará en fase de ductos.<br />
Gine<strong>com</strong>astia: patología exclusiva de varones por desarrollo mamas femeninas.<br />
Trastornos mamarios<br />
BENIGNOS<br />
Mastopatía fibroquística: causada por desequilibrios hormonales, presenta un significativo aumento del estroma,<br />
apareciendo quistes en los conductos. Cursa con dolor intenso.<br />
Fibroadenoma y displasia mamaria benigna: son masas de crecimiento lento, formadas por tejido epitelial y<br />
conjuntivo. Los conductos mamarios principales se dilatan y aparecen nódulos. Son trastornos indoloros.<br />
MALIGNOS<br />
La mama es uno de los lugares más frecuentes de cáncer en la mujer. La mayor parte tiene <strong>com</strong>o origen una<br />
transformación de células epiteliales de las vías excretoras. Los clínicos diferencian entre carcinoma ductal (si la<br />
metaplasia aparece entre el conducto galactóforo y el conducto terminal) e intraductal (si el cáncer se da en células<br />
de los conductillos intralobulillares o en los alveolos).<br />
Los genes BRCA1 y BRCA2 codifican para proteínas supresoras de tumores y en un porcentaje significativo de casos,<br />
estos marcadores indican predisposición a cáncer ovárico y mamario. Es por ello que se chequean estos marcadores<br />
frecuentemente, especialmente en pacientes con antecedentes familiares. Se re<strong>com</strong>ienda la realización de una<br />
mamectomía si se detectan.
Página151<br />
Generalidades<br />
APARATO REPRODUCTOR MASCULINO<br />
El aparato reproductor masculino se encarga de producir constantemente espermatozoides, los nutre y permite su<br />
introducción en el aparato genital femenino. Sintetiza y segrega andrógenos.<br />
Los genitales externos son el escroto y el pene. Los genitales internos son las gónadas, las vías espermáticas y las<br />
glándulas accesorias (próstata, vesículas seminales y glándulas bulbouretrales o de Cowper).<br />
ESCROTO<br />
Es una estructura par con forma de bolsa cutánea fibromuscular, dentro de la cual encontramos una cavidad<br />
revestida por una túnica vaginal. La piel es pilosa (contiene folículos pilosos). Dentro de cada escroto se aloja un<br />
testículo, que desciende de la cavidad abdominal por el conducto inguinal.<br />
Fallos de este descenso dan lugar a la criptorquidia.<br />
TESTÍCULOS<br />
Los testículos son órganos pares. Cada uno pesa 20 g. y se alojan fuera de la cavidad abdominal. Su cara posterior se<br />
encuentra asociada al epidídimo. Dentro del escroto, el testículo está sostenido por el cordón espermático.<br />
Los testículos son los órganos encargados de producir espermatozoides y testosterona.<br />
Embriología y desarrollo de las gónadas<br />
El desarrollo de las gónadas está en estrecha relación con el desarrollo del aparato urinario y ocurre en la cavidad<br />
abdominal.<br />
El mesodermo intermedio origina las crestas urogenitales que es tapizado por mesotelio celómico. Las células<br />
germinales primordiales provienen del saco vitelino y llegan hasta estas crestas, poblándolas.<br />
Estas células germinales proliferan e inducen a las células del mesotelio celómico y las crestas a que se dividan.<br />
Dentro de la gónada, se desarrollan unos cordones sexuales que acaban siendo cordones testiculares. Desde estas<br />
células se forman los túbulos seminíferos y rectos y la rete testis.
Página152<br />
En el feto, se produce el descenso testicular:<br />
Primera fase: los testículos<br />
descienden por un mecanismo<br />
controlado por el péptido MIS,<br />
segregado por las células de Sertoli.<br />
Segunda fase inguinoescrotal, no<br />
regulada por MIS, se produce un<br />
control por andrógenos. Esta fase<br />
depende del péptido CGRP,<br />
relacionado con la calcitonina.<br />
Se pueden producir fallos en el descenso de las gónadas:<br />
CRIPTORQUIDIA<br />
Se produce cuando el testículo no desciende. Es un trastorno asintomático que se debe tratar, porque si ninguno de<br />
los dos testículos desciende se produce esterilidad. Además, los testículos que quedan en la pared abdominal tienen<br />
una alta incidencia de malignización.<br />
La espermatogénesis requiere una temperatura inferior a la corporal. Para ello, los testículos salen de la cavidad<br />
abdominal y disponen del plexo venoso pampaniforme, donde la sangre circula y permite disipar el calor dentro de la<br />
gónada.<br />
Además, para que el descenso térmico no sea excesivo, los músculos cremáster y dartos elevan los testículos hacia la<br />
pared abdominal.<br />
El descenso del testículo puede ser <strong>com</strong>pleto o en ascensor (primero baja pero luego sube).<br />
Para diagnosticar la criptorquidia se pueden evidenciar uno de las consecuencias derivadas del problema: la atrofia<br />
testicular, que justifica la infertilidad.<br />
En un corte histológico lo las estructuras tubulares correspondientes a los túbulos seminíferos mantienen un diámetro<br />
similar. Si hay menos diámetro se puede hablar de atrofia testicular. El tamaño aproximado de todos los túbulos<br />
seminíferos es semejante y normalmente hay un equilibrio entre las células intersticiales de Leydig y los TS, pero en<br />
ocasiones el aumento del tejido intersticial hace disminuir el diámetro de los TS, viéndose muchas células de Leydig y<br />
túbulos seminíferos disminuidos. Esto es un indicativo de atrofia testicular.<br />
Histología General<br />
El testículo dispone de una cápsula muy gruesa: la túnica albungínea.<br />
En la zona posterior, esta túnica se engrosa hacia el interior,<br />
formando el mediastino testicular.<br />
Desde la cápsula parten una serie de tabiques que permiten dividir la<br />
gónada en secciones piramidales. Cada una de estas secciones<br />
corresponde a un lobulillo testicular. Existen en torno a 250-300<br />
lobulillos.<br />
Dentro de cada lobulillo, tenemos 3 o 4 túbulos seminíferos y entre<br />
medias de éstos, tejido intersticial.<br />
Se pueden distinguir varias capas en el testículo, que de fuera a<br />
dentro son:
Página153<br />
1. Escroto: piel pilosa y sebácea (basal hiperpigmentada)<br />
2. Túnica Dartos: tejido conjuntivo laxo con fibras musculares lisas del músculo Dartos que se encuentra adherido al<br />
escroto<br />
3. Tejido celular subcutáneo<br />
4. Fascia espermática externa<br />
5. Músculo cremáster: fibras musculares esqueléticas del músculo cremáster (del oblicuo < y transverso), que<br />
tienden a elevar los testículos con el frío.<br />
6. Fascia espermática interna<br />
7. Túnica vaginal propia del testículo (peritoneal): hojas parietal y visceral. Se forma al descender el testículo desde<br />
la cavidad abdominal arrastrando el peritoneo (las dos capas) al testículo.<br />
Más interna se dispone la túnica albugínea, un tejido conjuntivo denso irregular con rica inervación pero pobre<br />
vascularización. Se considera la primera verdadera primera capa del testículo.<br />
Hacia dentro la túnica albugínea, en la zona medial que corresponde al hilio del testículo, se produce un<br />
engrosamiento que se denomina MEDIASTINO o ANTRO DE HIGMORE, desde el cual se emiten una serie de tabiques<br />
que van a <strong>com</strong>partimentalizar al testículo en 250 lobulillos.<br />
Túbulos seminíferos<br />
Tienen un grosor de unos 150 μm y una longitud de 80 cm.<br />
Durante buena parte del desarrollo son cordones macizos, hasta que se abren a los 6 o 7 años. Histológicamente,<br />
presentan un epitelio estratificado, una lámina basal y un conjuntivo delgado con fibroblastos, células mioides y<br />
fibras colágenas.<br />
Cada uno de los 250 lobulillos (delimitados por los tabiques del antro de Higmore) va a tener de 1-4 túbulos<br />
seminíferos (existiendo aproximadamente 1000 túbulos seminíferos en ambos testículos), que <strong>com</strong>ienzan en forma<br />
de fondo de saco ciego y de forma tortuosa van hacia el mediastino donde finalmente se abren (están abiertos en<br />
sus dos extremos). Se van a unir entre ellos y van a dar lugar a unos tubos cortos y rectos denominados tubos rectos.<br />
Epitelio germinativo<br />
Se forma por dos tipos celulares: las células somáticas o de Sertoli, y las células espermatogénicas (espermatogonias,<br />
espermatocitos y espermátidas). Estas últimas se desplazan desde la zona basal hacia la luz.<br />
Células de Sertoli o Nodrizas<br />
Morfología<br />
Son células postmitóticas, minoritarias en adultos (10%). En la infancia y la vejez, son mayoritarias en el túbulo<br />
seminífero. Se apoyan en la lámina basal, y llegan hasta la luz.<br />
Tienen un núcleo claro y una membrana plasmática segmentada. Presentan un marcado nucléolo, así <strong>com</strong>o un<br />
desarrollo muy marcado de los orgánulos. Los cristales de Charcot-Böttcher muestran la presencia en su interior de<br />
filamentos.<br />
Son células altas, no prismáticas, con prolongaciones laterales que contactan unas células de Sertoli con otras,<br />
formando nichos o criptas donde se alojan las células espermatogénicas. Esto permite que las células de Sertoli creen<br />
microambientes específicos que favorecen determinadas fases de la espermatogénesis.
Página154<br />
Las células de Sertoli presentan un importante desarrollo de uniones estrechas de adhesión y nexos. Su REL está<br />
relacionado con uniones estrechas.<br />
Las células de Sertoli en el niño son las células mayoritarias, pero con la edad disminuye su porcentaje y van<br />
apareciendo otro tipo de células, constituyendo finalmente solo el 10% de las células dentro del túbulo seminífero<br />
en el adulto.<br />
Compartimentos. Barrera hematotesticular<br />
Las células de Sertoli presentan dos <strong>com</strong>partimentos<br />
<br />
<br />
Compartimento basal: lo constituyen las primeras prolongaciones de las células de Sertoli. En él encontramos<br />
espermatogonias y espermatocitos primarios recién formados.<br />
Compartimento adluminal: se encuentra por encima del anterior y aloja espermatocitos secundarios y<br />
espermátidas.<br />
Cada <strong>com</strong>partimento tiene un microambiente diferente y cerrado gracias a la presencia de la barrera<br />
hematotesticular.<br />
Esta barrera trata de separar estos dos <strong>com</strong>partimentos que a su vez tratan de diferenciar entre el ambiente propio,<br />
que permite el acceso de sangre y linfa, y las células del túbulo seminífero.<br />
Estos dos ambientes se crean ya que las células espermatogénicas sufren un proceso de diferenciación. Los gametos<br />
son células extrañas a nuestro organismo, por lo que serían atacados por el sistema inmunitario si no existiese eta<br />
separación. Además, a la hora de formar los gametos, hay que evitar que lleguen tóxicos, bacterias, etc., lo que<br />
conllevaría un desarrollo anormal de los gametos.<br />
En las células de Sertoli encontramos receptores para FSH y testosterona. Ambas son imprescindibles en<br />
espermatogénesis.<br />
Componentes de la barrera Hematotestiular:<br />
• Las células de Sertoli<br />
• La lámina basal del propio túbulo seminífero<br />
• La membrana de las células endoteliales del vaso<br />
Funciones<br />
· Sostén para células espermatogénicas<br />
· Establecen la barrera hematotesticular<br />
· Protegen y nutren a las células espermatogénicas<br />
· Inducen la espermatogénesis<br />
· Facilitan la migración de las células espermatogénicas<br />
· Fagocitan los cuerpos residuales que se dan en espermatogénesis<br />
· Facilitan la espermiación (liberación de espermátidas maduras por contracción)<br />
· Producen estrógenos<br />
· Excretan líquido testicular<br />
· Sintetizan ABP, transportadora de andrógenos, que fija la testosterona. ABP NO es un receptor.<br />
· Sintetiza otras proteínas que regulan la FSH: inhibina (inhibe síntesis de FSH), subunidades de activina (estimula<br />
síntesis de FSH).<br />
· Durante etapas fetales, sintetizan factor de regresión de Müller.
Página155<br />
Espermatogénesis<br />
La espermatogénesis en el varón se inicia en la pubertad y se mantiene toda la vida. Al dividirse, las células<br />
experimentan citocinesis in<strong>com</strong>pleta y quedan unidas por puentes, lo que implica una sincronización entre las fases.<br />
Se obtienen cuatro gametos por cada espermatogonia.<br />
Proceso<br />
Las células germinales se detectan desde la tercera semana en la pared del saco vitelino. Son células grandes, con<br />
prolongaciones, núcleo redondeado en que destacan uno o dos nucléolos, citoplasma con glucógeno, mitocondrias<br />
con crestas tubulares, polisomas y RER. Presentan fosfatasa alcalina.<br />
A partir de la cuarta semana migran y a la vez proliferan. En este proceso atraviesan el mesenterio para alojarse en la<br />
parte dorsal. Encontramos hasta 5000 células que pueblan el blastema gonadal.<br />
Este proceso migratorio es posible gracias a la interacción del factor de célula madre con el receptor c-kit. Si el<br />
ligando o el receptor faltan, tenemos un déficit de células germinales primordiales. Este ligando se descubrió en<br />
otros procesos, afectando a hematopoyesis, diferenciación de melanocitos y mastocitos.<br />
Las células germinales inducen la proliferación de células del mesonefros y el epitelio celómico. Se forman las crestas<br />
gonadales y unos cordones donde se sitúan las células que vienen migrando. Inicialmente, tenemos una gónada<br />
indiferenciada con cordones, en los que encontramos células de Sertoli y células germinales primordiales. Bajo ellas,<br />
encontramos una lámina basal y por fuera un estroma gonadal indiferenciado. A partir de la séptima semana de vida,<br />
esta gónada se desarrolla hacia la vía masculina. Hablamos entonces de gonocitos, no de células germinales, y los<br />
cordones dejan de ser sexuales para pasar a ser cordones testiculares, que constituyen los túbulos seminíferos. El<br />
estroma se transforma en conjuntivo intersticial.<br />
Inicialmente, estas células germinales y gonocitos, se sitúan en zona central de cordón. Son células grandes, con un<br />
solo nucléolo y mitocondrias con crestas tubulares. Presentan fosfatasa alcalina y c-kit. Se siguen dividiendo dentro<br />
de los cordones y a partir de semana 12 se desplazan hacia la base del cordón (hacia la lámina basal). Parte de las<br />
células que originan los gonocitos dan lugar a espermatogonias fetales. Son células más pequeñas que los gonocitos,<br />
su cromatina está mucho más condensada y tienen una gran reserva de glucógeno. Las espermatogonias tienen<br />
mitocondrias con crestas no tubulares, unidas entre sí mediante un elemento electrodenso, RNA+. Las<br />
espermatogonias también se dividen.<br />
Hacia la semana 16, <strong>com</strong>ienza la degeneración celular en las gónadas. Dura 4 semanas y afecta a cualquier tipo<br />
celular, pero especialmente a los gonocitos. De esta manera, desde la semana 21-22 los cordones celulares tienen<br />
<strong>com</strong>o célula principal la espermatogonia fetal. Estas espermatogonias fetales son reguladas por un factor promotor<br />
de la mitosis y otro inhibidor. Predomina el inhibidor.<br />
Durante el resto de la gestación no hay cambios en los cordones testiculares. Al nacer, los túbulos seminíferos que<br />
tienen sobre todo células de Sertoli. También encontramos gonocitos, espermatogonias fetales y espermatogonias A.<br />
Todas estas células se asientan sobre la lámina basal, bajo la cual hay miofibroblastos. Los gonocitos se siguen<br />
detectando hasta los 6 meses. Las espermatogonias fetales duran hasta 4 años, de modo que encontramos túbulos<br />
seminíferos que contienen sobre todo espermatogonias A. Las mitosis que experimentan estas células duran unos<br />
pocos meses y son dependientes de gonadotropinas (FSH y LH). Estas gonadotropinas disminuyen su nivel hasta que<br />
cesan sus mitosis en el sexto mes. Después se produce un reposo de 2 o 3 años, apareciendo una oleada de mitosis<br />
de origen desconocido (no dependen de gonadotropinas) que da lugar a las espermatogonias B.<br />
Antes de la pubertad, el testículo crece y madura. Se alargan y ensanchan los túbulos seminíferos y se ahuecan hacia<br />
los 9 años. Al final de esta etapa, hay un gran crecimiento de células germinales regulado por gonadotropinas, cuyos<br />
niveles aumentan irregularmente. Hay una tercera oleada proliferativa, cuando se estabilizan los niveles de<br />
gonadotropinas y se consigue ya espermatogénesis <strong>com</strong>pleta (12 ó 13 años).
Página156<br />
TIPOS CELULARES<br />
Espermatogonias<br />
Son células diploides. Cuando <strong>com</strong>ienzan a dividirse, las células que resultan están unidas por puentes.<br />
· Espermatogonia A: núcleos más ovalados, nucléolos periféricos, se dividen en densas y claras. Las densas tienen<br />
núcleo heterocromático, y las claras, eucromático.<br />
· Espermatogonia B: núcleo redondo, con cromatina densa. La secuencia de aparición de las espermatogonias y<br />
demás células espermatogéncias es la siguiente: A densa, A clara, B, espermatocitos primarios (tras diferenciación).<br />
Menos los espermatocitos primarios, todas las las espermatogonias están en el <strong>com</strong>partimento basal. Ascienden<br />
luego al <strong>com</strong>partimento luminal.<br />
Espermatocitos primarios<br />
Son células diploides. Los espermatocitos primarios, que derivan de la mitosis de las espermatogonias B, atraviesan<br />
la barrera hematotesticular para alcanzar el <strong>com</strong>partimento adluminal. Se establecen nuevas uniones bajo ellas y se<br />
degradan las existentes.<br />
Estos espermatocitos primarios son los que entran en la primera división meiótica, en la que se separan los<br />
cromosomas homólogos y hay una selección aleatoria de cromosomas paternos y maternos. Esta división es lenta y<br />
la profase ocupa la mayor parte del tiempo y pasa por los distintos estadios.<br />
Espermatocitos secundarios<br />
Se forman tras la primera división. Son células haploides. Su contenido en ADN es la mitad del original, y de forma<br />
inmediata se pasa a una segunda división sin periodo S, en la que se separan las cromátidas hermanas de forma<br />
rápida. Cada espermatocito forma las espermátidas.<br />
Espermátidas<br />
Son células haploides y no sufren ya nuevas divisiones mitóticas y meióticas, pero distan de ser los gametos<br />
originales. Para convertir la espermátida en espermatozoide, se necesita un proceso de diferenciación: la<br />
espermiogénesis.<br />
Inicialmente, las espermátidas son redondas, con gran cantidad de citoplasma, y se alojan en nichos limitados por las<br />
células de Sertoli. Cuando las prolongaciones avanzan, hay un cambio de forma de las células hasta que se obtienen<br />
células más alargadas, quedando la cabeza apuntando hacia el fondo del epitelio. Una vez que el proceso de<br />
espermiogénesis termina, se produce la espermiación, es decir, salida de la espermátida a la luz.
Página157<br />
Espermiogénesis<br />
Las espermátidas desarrollan un flagelo, forman un acrosoma y desarrollan un manguito, a la vez que ocurren<br />
cambios en el núcleo que tienen que ver con la condensación del DNA, para lo que se una a unas histonas que<br />
permiten mayor empaquetamiento. A la vez, el DNA no puede ser degradado porque los sistemas de reparación se<br />
eliminan, todo ello a<strong>com</strong>pañado de la pérdida progresiva del tamaño, generando restos que son fagocitados por las<br />
células de Sertoli.<br />
Este proceso da lugar a unas células aún inmóviles, inmaduras e incapaces de fecundar. Dura unos 74 días, al cabo de<br />
los cuales todavía tienen que sufrir un proceso de maduración.<br />
Las células madre son sensibles a agentes nocivos (déficits nutricionales, infecciones generales o locales,<br />
temperatura elevada, hormonas esteroideas, fármacos, mutágenos, pesticidas, HC halogenados de cadena corta,<br />
radiación...).<br />
Cuando la temperatura es superior a 35 Cº se puede producir: varicocele (dilatación anormal de las venas del cordón<br />
espermático), criptorquidia, torsión del cordón espermático, infarto hemorrágico, tratamiento antitumoral,<br />
parotiditis o inflamación de testículos en varones postpuberales.<br />
Túbulo seminífero<br />
El tubo seminífero presenta un epitelio organizado en diferentes capas, con distintos estadios de desarrollo y no<br />
distribuidas al azar. Las células se unen por puentes que permite una <strong>com</strong>unicación instantánea, que a su vez<br />
permite su sincronización.<br />
Asociación celular<br />
Una asociación celular es una <strong>com</strong>binación de células o tipos celulares, cada uno en su respectivo estadio, y que<br />
aparece simultáneamente en un momento dado en el tubo seminífero. El número de asociaciones es específico de<br />
cada especie (por ejemplo, en el varón humano hay seis asociaciones y en la rata, catorce) Las asociaciones van<br />
cambiando a lo largo del tiempo.<br />
Ciclo<br />
Un ciclo es una serie <strong>com</strong>pleta de asociaciones celulares que se continúan unas con otras en un punto del tubo<br />
seminífero.<br />
También se puede definir <strong>com</strong>o el tiempo que tarda una secuencia <strong>com</strong>pleta de asociaciones en cambiar en un punto<br />
concreto. Lo que dura un ciclo depende de la especie. Para que se segregue una espermátida, se necesitan varios<br />
ciclos y en el particular del hombre se precisan entre 4 y 6 ciclos (cada ciclo dura 16 días, haciendo un total de 74<br />
días).<br />
Onda
Página158<br />
Una onda es una sucesión de asociaciones celulares a lo largo del tubo seminífero. Mientras que las ondas en<br />
roedores son lineales, en el hombre son helicoidales; así, en un corte transversal se observan varios estadios.<br />
Tejido intersticial<br />
El tejido intersticial del testículo está formado por células de Leydig, aunque también hay macrófagos. Está muy<br />
vascularizado.<br />
Células de Leydig<br />
Pueden estar aisladas o en pequeños grupos.<br />
Son grandes, de núcleo redondeado y citoplasma eosinófilo.<br />
Si observamos este citoplasma, vemos abundantes orgánulos,<br />
sobre todo REL, mitocondrias con crestas tubulares e<br />
inclusiones lipídicas o de lipofuscina, en las que aparecen<br />
unas estructuras cristalinas con proteínas de función poco<br />
clara llamados cristales de Reinke. Estas células provienen de<br />
fibroblastos que se transforman entre la semana 8ª y 10ª del<br />
desarrollo embrionario. Son esenciales porque sintetizan y<br />
segregan andrógenos.<br />
Son activas durante unas pocas semanas o meses del<br />
neonato. Tras nacer, sufren una regresión y en la pubertad se<br />
activan, siendo responsables de la síntesis del 95% de la<br />
testosterona.<br />
Hay dos hormonas que regulan la síntesis de andrógenos: LH,<br />
que estimula la producción de testosterona; y prolactina, que<br />
induce el receptor para la LH. Sintetizan también la<br />
androstenediona y la dehidroepiandrosterona (DHEA).<br />
La hiperprolactemia se da cuando inhibe la función reproductora masculina, al reducirse la secreción de<br />
gonadotropinas. Se produce una disminución de la síntesis de testosterona, disminución de la Espermatogénesis y<br />
disfunción eréctil con infertilidad.<br />
Andrógenos<br />
Testosterona<br />
El 95% de la testosterona viaja unida a una globulina específica. El resto (5%) es sintetizado por células de la corteza<br />
suprarrenal. Esta testosterona atraviesa el tubo seminífero y alcanza la luz. Esta testosterona debe quedarse ahí,<br />
función que corre a cargo de la proteína ABP. Esto sucede porque interesa que la testosterona quede ahí, ya que es<br />
esencial para la espermatogénesis.<br />
La testosterona afecta a los conductos de Wolff que persisten y se diferencian en epidídimo, conducto deferente,<br />
conductos eyaculadores y vesículas seminales.<br />
DHT<br />
DHT es un andrógeno que se forma a partir de la testosterona para lo que se forma la 5-α-reductasa. Se forman el<br />
tubérculo, el pliegue, la tumefacción y el seno genitales; que originan el pene, el escroto y la próstata.<br />
Individuos con déficit en 5-α-reductasa tienen genitales internos masculinos, pero no desarrollan genitales externos<br />
masculinos y son confundidos con mujeres al nacer.<br />
Si no tenemos DHT, las estructuras forman genitales externos femeninos.
Página159<br />
Funciones de los andrógenos<br />
· En el feto: regulan la diferenciación de genitales externos e internos de tipo masculino y el crecimiento y desarrollo<br />
de estos genitales y la diferenciación del SNC.<br />
· En la pubertad: estimulan el inicio de la espermatogénesis y además se desarrollan los caracteres sexuales<br />
secundarios y se estimula la secreción de glándulas sebáceas.<br />
· En el adulto: mantenimiento de la espermatogénesis, caracteres sexuales secundarios y vías espermáticas.<br />
Tiene que ver con la conducta del individuo y afectan al desarrollo.<br />
Desarrollo sexual<br />
El sexo, desde el punto de vista genético, está determinado en la fecundación por la ausencia o presencia de<br />
cromosoma Y.<br />
Desde el punto de vista gonadal, depende del gen SRY, que codifica para el factor determinante testicular.<br />
Según una interpretación hormonal, el sexo viene determinado por la aparición de testosterona, DHT y MIS en el<br />
embrión masculino.<br />
La gónada es indiferenciada hasta la semana 7 del desarrollo, momento en que el gen SRY controla la diferenciación<br />
de gónada hacia el testículo: la corteza gonadal involuciona y la médula forma el testículo.<br />
Los pacientes con disgenesia gonadal XY presentan estructuras gonadales poco desarrolladas. Se observa una<br />
ausencia de virilización (persistencia de estructuras de origen mülleriano).<br />
MIS produce la regresión del conducto de Müller y si no existe se forman genitales femeninos.<br />
El síndrome de insensibilidad androgénica (SIA) se produce por un defecto del gen que codifica para el receptor de<br />
andrógenos.<br />
Presenta ausencia del desarrollo de conducto de Wolff e involución del conducto de Müller. Los testículos no<br />
descienden y hay genitales internos funcionales. Se desarrollan genitales externos femeninos y se encuentran<br />
elevadas concentraciones de andrógenos.<br />
En ausencia de andrógenos, los conductos de Wolff involucionan.<br />
Si en un feto hembra hay elevadas concentraciones de andrógenos, persisten los conductos de Müller y los<br />
conductos de Wolff.<br />
VÍAS ESPERMÁTICAS<br />
Origen embriológico<br />
La testosterona es necesaria para estimular el conducto mesonéfrico de Wolff. La parte más proximal del conducto<br />
se transforma en el conducto epididimario, muy contorneado. La parte más distal origina el conducto deferente. La<br />
parte extrema de la zona distal forma los conductos eyaculadores y las vesículas seminales. La zona de contacto con<br />
la gónada se transforma en los conductos eferentes.<br />
Túbulos rectos<br />
Son la primera vía de salida de los espermatozoides, en el mediastino testicular. Entre los túbulos seminíferos y los<br />
rectos, encontramos una porción corta donde solo encontramos células de Sertoli formando un epitelio simple.<br />
El túbulo recto es un tubo de luz estrecha, bastante corto, y está formado por células epiteliales cúbicas bajas con<br />
microvellosidades y algún cilio. Presentan un gran desarrollo de uniones occludens en el dominio apical. Puede<br />
aparecer alguna célula mioide.<br />
Rete testis<br />
Desde el mediastino testicular, pasamos a otros canales que forman una red laberíntica muy anastomosada con<br />
elevados niveles de andrógenos, más concentrados que en sangre. A este nivel seguimos estando en mediastino<br />
testicular, es decir, hay niveles de testosterona importantes. Estos canales están tapizados por un epitelio cúbico<br />
simple con células con cilios y microvellosidades. Debajo, encontramos un conjuntivo con células mioides. Entre los
Página160<br />
canales hay abundantes vasos linfáticos y sanguíneos, y también células de Leydig. Todos estos canales convergen<br />
para formar conductillos eferentes que atraviesan la túnica albugínea para pasar al epidídimo.<br />
Conductillos eferentes<br />
Estas vías se encuentran muy enrolladas, formando los conos eferentes. La punta del cono se orienta hacia el<br />
mediastino testicular. Estos conductillos eferentes presentan una luz muy amplia, con un margen festoneado debido<br />
a que se alternan células de distinta altura, por lo que se dice que el epitelio es prismático simple o<br />
pseudoetratificado. Encontramos un mínima cantidad de conjuntivo y miocitos lisos circulares.<br />
En los conductillos eferentes encontramos dos tipos celulares:<br />
· Células principales: se encuentran en el epitelio más bajo. Tienen función de absorción, para lo que tienen<br />
microvellosidades, vesículas endocíticas, invaginaciones tubulares con mitocondrias. Absorben gran parte del líquido<br />
de la luz.<br />
· Células ciliadas: son más altas y presentan cilios. Se encargan de batir el contenido luminal hacia el epidídimo. Este<br />
flujo de líquido es favorecido por las células musculares y los cilios.<br />
Epidídimo<br />
Estructura y morfología<br />
Es un conducto único al que convergen todos los conductillos eferentes. El epidídimo mide más de 5 cm., y se<br />
encuentra enrollado sobre sí mismo apretadamente hasta conseguir un volumen de 5 cm. A lo largo de este tubo se<br />
consigue la maduración de los espermatozoides y además permite almacenarlos temporalmente.<br />
El epidídimo se divide en cabeza, cuerpo y cola. Por ser tan largo, no tiene las mismas características en toda su<br />
longitud: es más estrecho al principio y más amplio en la cola. Además, presenta un epitelio más alto al principio y va<br />
siendo más bajo según avanzamos. Algunas mutaciones afectan a la formación de los segmentos del conducto (son<br />
los genes Bmp 4, 7 y 8).<br />
Esquema Diferencias Epitelio en Epidídimo<br />
En la cabeza encontramos los conos eferentes y estroma, así <strong>com</strong>o altas concentraciones de andrógenos.<br />
El epidídimo está tapizado por un epitelio pseudoestratificado, con núcleos en dos niveles. Debajo encontramos una<br />
lámina propia delgada y una capa muscular que va creciendo según avanzamos. Inicialmente, solo encontramos<br />
miocitos dispuestos circularmente, y a partir del cuerpo aparecen dispuestos longitudinalmente. En la cola<br />
encontremos tres estratos: uno longitudinal, otro circular, y un último longitudinal. Finalmente encontramos una<br />
adventicia.<br />
En el epitelio encontramos dos tipos celulares: las células principales y las células basales.
Página161<br />
Células principales<br />
Son células prismáticas, y van perdiendo altura según avanzamos. Su membrana presenta microvellosidades largas y<br />
ramificadas (esterocilios). Encontramos vesículas endocíticas y pinocíticas, así <strong>com</strong>o lisosomas, RER, REL, Golgi e<br />
invaginaciones de la membrana basal.<br />
Estas células absorben el 90% del líquido testicular y fagocitan cuerpos residuales no eliminados por las células de<br />
Sertoli, así <strong>com</strong>o espermatozodies en malas condiciones.<br />
Segregan moléculas que inmovilizan espermatozoides (productos ácidos <strong>com</strong>o carnitina y ácido siálico) y facilitan su<br />
maduración (glucoproteínas y glicerofosforilcolina).<br />
Transporte de espermatozoides<br />
En este transporte juegan un papel fundamental las contracciones de la capa muscular. En las zonas proximal y<br />
media, se dan contracciones rítmicas y espontáneas (peristaltismo) que mueven los espermatozoides. En la zona más<br />
distal, encontramos contracciones más intensas, sincronizadas con las contracciones del conducto deferente. Estas<br />
contracciones responden a un control adrenérgico y se desencadenan en la estimulación sexual. Más breve en zona<br />
inicial y más largo en zona terminal.<br />
Maduración de espermatozoides<br />
Este proceso se produce en la zona media. Necesita 10-12 días y está regulada por testosterona. Los<br />
espermatozoides sufren diferentes cambios en la cromatina (se estabiliza), en la membrana plasmática (cambia su<br />
carga y adquiere proteínas que actúan <strong>com</strong>o receptores), en la capacidad de unión a la zona pelúcida y en la<br />
movilidad (la adquiere en modo anterógrado).<br />
Almacenamiento de espermatozoides<br />
Esta función ocurre en la zona final, es limitado, y si no hay eyaculación, los espermatozoides se eliminan en orina.<br />
Células basales<br />
Aparecen entre ellas linfocitos intraepiteliales o del halo, por primera vez en todo el testículo.<br />
Conducto deferente<br />
Estructura<br />
Es un tubo con una importante pared muscular, con una longitud de 45 cm. Inicialmente, se encuentra dentro del<br />
cordón espermático junto con arterias, nervios, el plexo venoso pampaniforme y el músculo cremáster. Va<br />
ascendiendo hasta la cavidad abdominal y rodea la vejiga, para terminar en la ampolla.<br />
Tiene una luz muy irregular debido a la importante capa muscular. Es una luz más estrecha que en la cola del<br />
epidídimo.<br />
Encontramos pliegues longitudinales de la mucosa, una adventicia de conjuntivo laxo y adipocitos.<br />
Está tapizado por un epitelio pseudoestratificado que pierde altura. Las células son de dos tipos:<br />
• Células Basales (mayoritarias)<br />
• Células Principales o Cilíndricas muestran estereocilios (favorecen la absorción) y van desapareciendo hacia<br />
distal.<br />
Encontramos una lámina propia delgada de fibras elásticas. Después aparece una importante pared muscular con<br />
tres estratos, siendo más grueso el medio (circular). Esta muscular le da gran consistencia al conducto. Presenta una<br />
inervación adrenérgica en sus miocitos. Estos tres estratos no son independientes, sino que los miocitos se disponen<br />
helicoidalmente. Esta disposición permite el proceso de eyaculación gracias a unos movimientos rápidos y alternos<br />
de aspirado y eyección.
Página162<br />
En una vasectomía no se corta todo el cordón espermático, sino únicamente el CD, ya que no se pueden cortan los<br />
plexos venosos pampiniformes porque su función es esencial para la “ambientación” de los espermatozoides (muy<br />
sensibles a cambios de tª), ya que enfrían al testículo (recordemos que los testículos deben mantenerse a 2º menos<br />
que la tª corporal normal, es decir, a 35º, por eso se alojan fuera de la cavidad abdominal); por tanto se abre el<br />
cordón espermático y una vez abierto se localiza el CD, se tira de él, se corta a ambos lados y se anuda.<br />
Eyaculación<br />
Antes de la eyaculación, los conductos eyaculadores están cerrados. Solamente se produce la contracción en la zona<br />
donde se disponen longitudinalmente: el conducto se acorta pero se ensancha. Aumenta la luz, cae la presión y se<br />
produce una aspiración por la zona más próxima del epidídimo.<br />
En eyaculación suceden dos hechos sincronizados: se abren los conductos eyaculadores y toda la musculatura del<br />
conducto deferente se contrae (no solo longitudinal). Es una contracción rítmica de control adrenérgico que hace<br />
que la luz, antes ensanchada se cierre, incrementando la presión y provocando el avance de los espermatozoides.<br />
Ampolla deferente<br />
La ampolla está formada por un epitelio semejante al del CD, con células cúbica/cilíndricas dependiendo del<br />
momento en el que se encuentre, y recibe al conducto de secreción de la vesícula seminal. De esta dilatación sale el<br />
conducto eyaculador.<br />
El CD termina en una ampolla donde vierten su contenido las vesículas seminales, y de esta ampolla parte el<br />
conducto eyaculador, el cual vierte su contenido en una porción de la uretra prostática denominada montículo<br />
seminal o vero montanum y desde ahí (desde la ampolla) el conducto deferente se hace intraprostático y pasa a<br />
denominarse conducto eyaculador.<br />
Conducto eyaculador<br />
Este conducto es mucho más delgado que el deferente (se forma a partir de la ampolla deferente) y penetra en la<br />
próstata y permite que la secreción de espermatozoides y la secreción de las vesículas seminales lleguen a la uretra.<br />
Tiene una mucosa delgada, epitelio estratificado simple, lámina laxa con fibras elásticas y solo una capa muscular en<br />
la zona más proximal.<br />
Uretra<br />
La uretra se divide en tres porciones: prostática, membranosa y peneana.<br />
En la prostática se vierte la secreción de las vesículas prostáticas y en la peneana la de las glándulas de Cowper.<br />
En la porción prostática, la uretra presenta epitelio de transición o urotelio, que es un epitelio prismático<br />
estratificado y glándulas de Littré (mucosas) e intraepiteliales. En la uretra peneana, estas glándulas son más grandes<br />
y en el cuerpo esponjoso del pene se sitúan entre las trabéculas. Sintetizan y segregan un producto que contiene<br />
sobre todo glucosaminoglicanos.<br />
A nivel de la uretra membranosa hay dos esfínteres, uno de musculatura lisa y otro de musculatura esquelética.
Página163<br />
PENE<br />
Estructura e histología<br />
El pene es el órgano copulador masculino. Está recubierto por una piel<br />
fina y pigmentada, con terminaciones nerviosas sensitivas, y deslizable.<br />
Presenta mucha musculatura lisa que se continúa con el dartos del<br />
escroto. Esta musculatura se organiza en dos estratos longitudinal y<br />
circular. En la base del pene encontramos vello. El tejido subcutáneo del<br />
pene es una zona libre de grasa.<br />
El extremo del pene o glande está recubierto por un pliegue cutáneo, el<br />
prepucio. Hay aquí glándulas sebáceas de Tyson o prepuciales que<br />
secretan un sebo denominado esmegma. El pene presenta una doble<br />
inervación, simpática y parasimpática, además de otra sensitiva. Permite<br />
las respuestas de erección y eyaculación.<br />
Histológicamente, el pene se forma por tres cuerpos cilíndricos: dos<br />
dorsales (cuerpos cavernosos) y uno ventral (esponjoso). Son cuerpos de<br />
tejido eréctil y cada uno de ellos está rodeado por una fascia muy fibrosa<br />
de tejido conjuntivo.<br />
Cuerpo cavernoso<br />
Los cuerpos cavernosos están rodeados por la fascia llamada túnica albugínea, muy gruesa y fibrosa. En la zona<br />
media, las túnicas albugíneas que rodean los cuerpos se fusionan, formando un tabique. Este tabique no es <strong>com</strong>pleto<br />
a lo largo de todo el pene, y en la zona anterior es más delgado.<br />
Su estructura está formada por cavernas: espacios vasculares tremendamente anastomosados y muy irregulares en<br />
cuanto a su forma. Cada caverna se reviste por un endotelio. Entre las cavernas quedan trabéculas de tejido<br />
conjuntivo, en las cuales son abundantes los miocitos. Las cavernas más grandes se sitúan en la zona central. En la<br />
periferia, son más pequeñas y estrechas y se <strong>com</strong>unican con un plexo venoso.<br />
Cuerpo esponjoso<br />
Aquí la túnica albugínea es más delgada, con abundantes fibras elásticas. Las cavernas son similares y también<br />
encontramos un plexo venoso. En la zona terminal ya no tenemos cavernas, y ya aparece la uretra. Gran desarrollo<br />
de musculatura.<br />
La sangre llega a las cavernas desde las arterias helicinas y son drenadas por canales venosos.<br />
Vascularización<br />
Cuerpos cavernosos<br />
Los cuerpos cavernosos del pene están irrigados por arterias helicinas, que proceden de la arteria profunda de cada<br />
cuerpo cavernoso. Las arterias hemicilas tienen una túnica media muy gruesa, con miocitos de inervación<br />
parasimpática. Los miocitos se encuentran también en la íntima, <strong>com</strong>o almohadillas que se disponen<br />
longitudinalmente. La regulación de esta musculatura provoca el cierre de las cavernas.<br />
La sangre sale hacia vénulas postcavernosas que se unen formando un plexo venoso periférico de venas de paredes<br />
finas.<br />
Existen también numerosas anastomosis arteriovenosas que conectan la arteria profunda del pene con el sistema<br />
venoso periférico. La sangre de este plexo pasa a unas venas de mayor calibre, que atraviesan la albugínea y van<br />
hasta la vena dorsal profunda del pene.
Página164<br />
Cuerpo esponjoso<br />
Las cavernas en el cuerpo esponjoso se irrigan de manera similar, pero el drenaje va a la vena dorsal superficial del<br />
pene, que recoge también sangre de la piel.<br />
Erección<br />
El tejido eréctil es en realidad un fenómeno vascular, que se inicia <strong>com</strong>o respuesta a unos estímulos que llegan al<br />
SNC. Esta estimulación sexual hace que el sistema nervioso parasimpático acabe relajando la musculatura de las<br />
arterias helicinas y los miocitos de las trabéculas. Esto permite la erección. El sistema nervioso simpático controla el<br />
proceso de emisión y el cierre del cuello vesical, esencial para la eyaculación.<br />
El neurotransmisor fundamental de la erección es el NO.<br />
Algunos autores hablan de una secreción directa de NO por fibras parasimpáticas, mientras que otros dicen que<br />
estas fibras segregan ACh y activa así el endotelio, que es el que sintetiza y secreta NO. En cualquier caso, estos<br />
niveles de NO difunden, activando los miocitos. Estos miocitos presentan uniones gap que permiten una activación<br />
sincitial de la mayoría de los miocitos.<br />
El NO estimula entonces la cascada de la adenilato ciclasa, incrementando el AMPc y relajándose la musculatura. La<br />
sangre accede a las cavernas y se <strong>com</strong>primen las venas pequeñas, se estanca la sangre y se produce la erección. Esta<br />
erección es menor en el cuerpo esponjoso porque no debe ocluirse la uretra.<br />
La erección termina cuando la fosfodiesterasa degrada el AMPc.<br />
Se aumentan entonces los niveles de calcio y se contraen los miocitos, aumentando el tono en trabéculas y la presión<br />
de las almohadillas de las arterias. Se reduce la entrada de sangre a las cavernas, la sangre drena y la erección<br />
termina.<br />
La disfunción eréctil se produce por múltiples causas y, si el parasimpático no está afectado, se puede tratar<br />
utilizando el sildenafil (Viagra), que bloquea la fosfodiesterasa.<br />
PENE FLÁCIDO: las cavernas están colapsadas en los cuerpos cavernosos y perfundidas en el esponjoso.<br />
PENE ERECTO: todas las cavernas están llenas de sangre, que queda retenida en las cavernas. Aumenta el volumen<br />
del pene, y además las cavernas presionan las venas impidiendo el drenaje.<br />
GLÁNDULAS<br />
Hay tres glándulas accesorias al aparato genital masculino: vesículas seminales, próstatas y glándula de Cowper.<br />
Vesículas seminales<br />
Histología<br />
Son órganos pares, pequeños. Derivan de la ampolla del conducto deferente y no son más que un conducto muy<br />
retorcido (15 cm de longitud cuando está desenrollado). El nombre se dio porque se pensaba que era un lugar donde<br />
se almacenaban espermatozoides, cosa que no es así.<br />
Tiene una luz muy irregular, tapizada por mucosa con pliegues que se ramifican.<br />
El epitelio es seudoestratificado, y es dependiente de testosterona (necesita de testosterona para mantenerse).<br />
Aparecen dos tipos de células en esta mucosa:<br />
- Basales<br />
- Cilíndricas o principales: secretan fructosa, flavinas y demás <strong>com</strong>ponentes del esperma.<br />
Se apoya sobre una lámina propia laxa con una importante capa muscular de dos estratos: uno circular interno y otro<br />
longitudinal externo. Finalmente encontramos una adventicia.<br />
El epitelio de estas vesículas seminales va disminuyendo de altura conforme avanza la edad (es cúbico bajo en<br />
personas mayores). Presenta actividad secretora apocrina, muchos orgánulos, inclusiones de lipofuscina, gránulos<br />
eosinófilos de secreción. Estas vesículas funcionan desde la pubertad.
Página165<br />
Secreción<br />
La secreción de las vesículas seminales es una secreción espesa y alcalina, que conforma entre el 60% y el 80% del<br />
esperma y forma la parte final del eyaculado. Contiene fructosa (aporte metabólico de espermazoides),<br />
prostaglandinas, proteínas, aminoácidos, potasio y flavinas. Las flavinas son citocromos muy útiles en medicina legal<br />
porque son fluorescentes y evidencian la eyaculación. Esta secreción nutre a los espermatozoides y algunas proteínas<br />
son responsables de la coagulación del semen.<br />
La secreción sale por los conductos situados entre el deferente y los eyaculadores, que atraviesan la cápsula de la<br />
próstata y se abren a eyaculador. La pared de la vesícula se contrae en eyaculación para añadir la secreción de las<br />
vesículas a la parte final del eyaculado.<br />
Próstata<br />
Generalidades<br />
Es un órgano impar con forma de castaña y un peso de 20 g.,<br />
situado bajo la vejiga urinaria.<br />
Dentro encontramos los conductos eyaculadores y la uretra<br />
prostática.<br />
La próstata está rodeada por una cápsula no especialmente<br />
gruesa, pero sí muy firme y consistente.<br />
Contiene un estroma que contiene abundantes miocitos que<br />
le dan consistencia y lo convierten en un órgano palpable.<br />
El estroma contiene además tejido conjuntivo denso irregular que se organiza con las fibras musculares de manera<br />
aleatoria pero formando capas concéntricas alrededor de la luz uretral.<br />
El parénquima son glándulas túbuloalveolares que drenan por conductos excretores o a la uretra directamente, de<br />
manera que encontramos más glándulas que conductos. Las secreciones van a la uretra en el colículo seminal.<br />
Regiones anatómicas<br />
Anatómicamente, la próstata tiene 4 regiones: periférica, central, periuretral y transicional. Son regiones<br />
concéntricas.<br />
La región periférica es de mayor tamaño y es la que se inflama y maligniza con mayor facilidad.<br />
La zona central es más resistente a la infección y malignización. Es donde se incluyen los conductos eyaculadores, y<br />
tiene un origen embriológico diferente<br />
Cuando las zonas transicional y periuretral proliferan en exceso, se produce la hiperplasia benigna prostática (HBP).<br />
Hay un desarrollo descontrolado de glándulas, que forman nódulos y <strong>com</strong>primen la uretra causando dificultad en la<br />
micción (retención de orina, cistitis…). Proliferan <strong>com</strong>o consecuencia de una estimulación de DHT, se forma a partir de<br />
testosterona por la 5-α-reductasa. Esta patología se trata con inhibidores de esa enzima.<br />
Regiones histológicas<br />
Histológicamente, hablamos de 3 regiones concéntricas en la próstata, en función del tipo de glándulas: periférica<br />
(glándulas principales), submucosa (glándulas submucosas ramificadas y de conductos largos) y mucosa (glándulas<br />
mucosas, con conductos más cortos).<br />
Solo las glándulas submucosas y principales tienen conductos, más o menos largos. Desembocan a ambos lados de<br />
las crestas uretrales.<br />
El epitelio de la próstata es inicialmente seudoestratificado y en la uretra es ya urotelio.
Página166<br />
Cuerpos amiláceos o calcáreos<br />
La luz de la próstata presenta los cuerpos amiláceos. Tienen un tamaño de 1-2 mm, son eosinófilos, con una<br />
estructura laminar concéntrica. Se forman cuando, alrededor de algún resto, se precipitan elementos de la secreción<br />
prostática, <strong>com</strong>o glicoproteínas y calcio.<br />
Glándulas<br />
Las glándulas de la próstata tienen una trayectoria contorneada y un tamaño acinar irregular. Su epitelio es variable<br />
(liso, con pliegues…) y generalmente cilíndrico seudoestratificado (en realidad biestratificado). En general, cuanto<br />
más activa es la glándula, mayor altura tiene el epitelio. Su actividad es dependiente de testosterona.<br />
Se describen 3 tipos celulares:<br />
· Principales o secretoras: presentan abundante RER y aparato de Golgi debido a su función secretora<br />
· Basales: encargadas de la renovación. Están por fuera de las luces alveolares (zona periférica) y son escasas,<br />
aplanadas y con núcleos hipercromáticos; si no se ven bien tendremos que recurrir a técnicas de IHQ (CK34B12).<br />
Son muy importantes a la hora de determinar si el crecimiento glandular es normal hiperplásico o tumoral displásico,<br />
es decir, su presencia/ausencia es importante para valorar la existencia de un carcinoma de próstata. Las células<br />
basales nos sirven para detectar la presencia de tumores prostáticos. Al igual que las células mioepiteliales en la<br />
mama (un acino creciendo sin presencia de células mioepiteliales detectables con las técnicas de IHQ actina de<br />
músculo liso o P6 , probablemente el crecimiento fuera neoplásico o cancerígeno), si vemos luces, un crecimiento<br />
epitelial glandular y no detectamos a las células basales, significa que el crecimiento es neoplásico o tumoral.<br />
· Endocrinas: sintetizan serotonina<br />
Secreción prostática<br />
Es una secreción de color blanquecino, lechosa, muy fluida y alcalina. Forma parte de la porción inicial del eyaculado.<br />
Contiene elementos <strong>com</strong>o FAP (una fosfatasa específica), PSA (antígeno prostático específico), ácido cítrico y<br />
fibrinolisina. Menos abundantes son las prostaglandinas, poliaminas (poliamina espermina con valor diagnóstico),<br />
proteasas, amilasas, inmunoglobulinas y el zinc.<br />
La PSA es la molécula marcador del carcinoma de próstata. Supuso un enorme avance ya que el cáncer prostático era<br />
asintomático y se detectaba muy tarde. En varones sanos, PSA está circulante en niveles bajos y procede sobre todo<br />
del hígado. Si se detectan altos niveles de PSA, ello indica un desarrollo maligno de la próstata.<br />
Esta secreción neutraliza el pH acido de la vagina. Aporta nutrientes, estabiliza la cromatina y aporta <strong>com</strong>ponentes<br />
bacteriostáticos. Hace que el semen más líquido.<br />
La próstata <strong>com</strong>o glándula solo es activa desde la pubertad.<br />
Hasta entonces está indiferenciada: es más pequeña y apenas se observan conductos. El desarrollo de epitelio y las<br />
glándulas es posible al elevarse los niveles de testosterona, momento en que se desarrollan los adenómeros y las<br />
células secretoras.<br />
Glándulas de Cowper o bulbouretrales<br />
Son glándulas pares, pequeñas, situadas detrás de la uretra membranosa. Tienen un conducto excretor que<br />
desemboca en la uretra peneana proximal, en el cuerpo esponjoso. Son glándulas túbuloalveolares de secreción<br />
mucosa, con un conducto excretor tapizado por un epitelio prismático simple.<br />
La secreción de estas glándulas es un líquido claro, que precede a la emisión del semen. El pH alcalino de esta<br />
secreción neutraliza el pH ácido de la orina y actúa <strong>com</strong>o lubricante de la uretra peneana. Contiene sobre todo<br />
galactosa y ácido siálico, aunque también encontramos galactosamina, ácido galactourónico y metilpentosa.
Página167<br />
SEMEN<br />
Es una secreción alcalina, lo que le permite neutralizar el pH ácido de la uretra y el del medio vaginal. El semen se<br />
forma de líquido testicular, secreciones del epidídimo, del conducto deferente, la próstata, las vesículas seminales y<br />
las glándulas de Cowper. El 10% del semen corresponde a espermatozoides que se encuentran en una concentración<br />
de 100 millones por mililitro.<br />
Se considera que cuando el número de espermatozoides disminuye a 20 millones/ml, es signo de infertilidad a causa<br />
de oligospermia.<br />
En individuos sanos, el 20% de los espermatozoides eyaculados son anormales y el 25% inmóviles.<br />
La criopreservación es el almcenamiento de espermatozoides; si es en líquido se mantiene su capacidad fertilizante,<br />
lo que posibilita la inseminación artificial y la FIV (Fecundación In Vitro) con semen congelado.
Página168
Página169<br />
APARATO URINARIO<br />
El aparato urinario está formado por los riñones y las vías urinarias, es decir, los uréteres, la vejiga urinaria y la<br />
uretra.<br />
Funciones<br />
El aparato urinario se encarga de la producción de la orina, controlar su <strong>com</strong>posición, almacenarla termporalmente y<br />
finalmente evacuarla. En el varón también permite la evacuación del semen.<br />
Además, se encarga de regular la homeostasis del agua y los electrolitos. Esto implica un control sobre el volumen<br />
del líquido extracelular y su osmolaridad. Ligado a esto, el aparato urinario es responsable del control del equilibrio<br />
ácido-base. En esto intervienen los procesos de filtración, reabsorción y excreción de electrolitos (aminoácidos,<br />
glucosa, péptidos, sodio, cloro, calcio, PO 3 - y agua).<br />
También sintetiza y segrega factores y hormonas <strong>com</strong>o la eritropoyetina (EPO) y la renina y activa la vitamina D 3 en<br />
1,25-hidroxicolecalciferol. Parte de estas hormonas se relacionan con el control de la tensión arterial.<br />
Participa en el metabolismo porque elimina restos celulares, productos extraños (<strong>com</strong>o medicamentos) y catabolitos,<br />
en lo que intervienen la filtración y la excreción (catabolitos de proteínas, purinas y productos hidrogenados).<br />
RIÑÓN<br />
Es un órgano par con forma de haba y de medidas 11x5x4cm y pesa unos 120-300g.<br />
Está situado en la cavidad retroperitoneal y presenta, especialmente en su cara cóncava donde se localiza el hilio<br />
renal, tejido graso, y en el polo superior se sitúa la glándula suprarrenal.<br />
El seno renal está formado por vasos, nervios y la pelvis renal.
Página170<br />
Vascularización<br />
La arteria renal penetra por el hilio y se ramifica formando las<br />
arterias interlobulares, que ascienden por un territorio denominado<br />
columnas de Bertin. Al llegar al límite entre corteza y médula, las<br />
arterias interlobulares dan paso a las arterias arciformes, que tienen<br />
una trayectoria horizontal. Después encontramos las arterias<br />
interlobulillares, que sí son verticales y ascienden hasta la cápsula.<br />
Cada territorio está irrigado por una de las arterias interlobulillares y<br />
no hay anastomosis entre ellas. Cuando se produce una obstrucción<br />
arterial habrá una necrosis focal. Así, en cada infarto renal muere una<br />
región del riñón muy delimitada.<br />
Las arterias interlobulillares dan dos ramas. Una circula entre los<br />
túbulos corticales y oxigena la zona. La otra rama va a penetrar<br />
dentro del corpúsculo renal. Esta rama se denomina arteriola<br />
aferente. Dentro del corpúsculo renal da paso al glomérulo (capilares<br />
glomerulares), un amasijo de capilares en los que no se produce<br />
intercambio de oxígeno y dióxido de carbono. La sangre sale del<br />
corpúsculo por la arteriola eferente, que dará paso a una segunda<br />
red capilar, los vasos rectos o arterias rectas que circulan entre los<br />
túbulos de las nefronas y los conductos colectores, parte de los<br />
cuales están en zona cortical y parte en zona medular; en esta zona sí se produce intercambio de gases.<br />
Al final llegaremos a unos vasos rectos venosos y a las venas arciformes, que están en el límite entre corteza y<br />
médula. Estas venas también reciben la sangre de las arterias que irrigaban los espacios corticales por drenaje de las<br />
venas interlobulillares. Forman las venas interlobulares y salen por la vena renal a través del hilio.<br />
ESTRUCTURA HISTOLÓGICA<br />
Encontramos un estroma formado por la cápsula y el intersticio, y un parénquima con elementos epiteliales que se<br />
divide en corteza y médula.<br />
La cápsula es <strong>com</strong>pleta y en el hilio se introduce y forma la cubierta del seno. En la zona externa encontramos<br />
fibroblastos y fibras colágenas. Sin embargo, en la parte interna hay miofibroblastos con capacidad contráctil que<br />
permite resistir los cambios de tensión y volumen del hilio.<br />
Corteza y Médula<br />
La corteza es una zona rojiza en la que está la mayor parte del volumen sanguíneo (90-95%). Rodea por <strong>com</strong>pleto a la<br />
médula y tiene aspecto granuloso porque en ella están los corpúsculos renales o corpúsculos de Malpigio. Si<br />
observamos la corteza veremos que no toda ella muestra dicho aspecto granuloso, sino unas zonas con estriaciones<br />
verticales perpendiculares a la cápsula que se denominan rayos medulares de Ferrein (aprox. 500).<br />
La médula es la zona rodeada por la corteza. Es mucho más gruesa que ésta (el doble) y es más pálida. No tiene<br />
aspecto granuloso sino que hay estriaciones abundantes. Está formada por 8-18 estructuras en forma de pirámide,<br />
las pirámides de Malpigio separadas entre sí por unas zonas con aspecto similar a la corteza denominadas columnas<br />
de Bertin. Las pirámides tienen la base cubierta por la corteza y la cúspide orientada hacia el seno renal.<br />
Histológicamente la médula se divide en externa e interna; la médula externa a su vez se divide en franja externa e<br />
interna.
Página171<br />
Lóbulo y Lobulillo Renal<br />
Un lóbulo renal es una pirámide de Malpigio junto con el tejido cortical que la rodea. En algunas especies el riñón<br />
sólo tiene un lóbulo, <strong>com</strong>o en la rata y el conejo; sin embargo, en el hombre el riñón es multilobular, aunque esta<br />
lobulación sólo es bien visible hasta el primer año de vida y luego <strong>com</strong>ienza a hacerse difusa.<br />
Un lobulillo renal hace referencia a elementos que son sólo corticales. Para un anatómico es la parte de la corteza<br />
flanqueada por dos arterias interlobulillares ascendentes. Si nos fijamos en sus elementos, es la parte de la corteza<br />
en la que tenemos un conducto colector y las nefronas que drenan a él. En el eje del lobulillo o rayo medular<br />
encontramos el conducto colector y las porciones rectas de las nefronas. A ambos lados del eje y dentro del lobulillo<br />
hay zonas corticales en las que localizamos los corpúsculos y las zonas contorneadas de la corteza; esta zona lateral<br />
se denomina laberinto cortical.<br />
Túbulo Urinario<br />
La unidad renal es un túbulo urinario. Un túbulo urinario está formado por una nefrona y un tubo colector, de<br />
distinto origen embriológico: la nefrona procede del metanefros y el tubo colector del botón ureteral. Entre las<br />
unidades renales encontramos tejido intersticial, que es conjuntivo bastante laxo en el que hay vasos sanguíneos,<br />
linfáticos y fibras nerviosas.<br />
Nefrona<br />
Una nefrona (aprox 2 millones en cada riñón) es un tubo largo que tiene un extremo proximal ciego y un extremo<br />
abierto que permite la salida de la orina al tubo colector. El extremo proximal ciego está dilatado e invaginado ya que<br />
dentro de él queda alojado el glomérulo (ovillo capilar); tiene una capa parietal y una visceral que forman juntas la<br />
cápsula de Bowman. El segundo tramo de la nefrona es el túbulo proximal, que tiene una porción contorneada y una<br />
porción recta. Después tenemos el segmento delgado, que es mucho más fino. Da paso al túbulo distal, inicialmente<br />
presenta una porción distal recta y luego una porción contorneada; además, contiene la mácula densa.<br />
Esquema Nefrona Extendida<br />
El asa de Henle es la suma del segmento delgado con las dos porciones rectas, la del túbulo proximal y la del distal.<br />
Hay nefronas que tienen un asa de Henle corta y otras que la tienen muy larga, lo que permite diferencias fisiológicas
Página172<br />
importantes. En las cortas el segmento delgado descendente gira y da paso al túbulo recto distal. En las largas, el<br />
segmento delgado es descendente y ascendente.<br />
Corpúsculo Renal<br />
La cápsula de Bowman (de la nefrona) y el glomérulo forman el corpúsculo renal. En él se define un polo vascular por<br />
el que entra la sangre por la arteriola aferente y sale por la eferente. En el extremo opuesto tenemos el polo<br />
urinario, que se continúa con el túbulo proximal y en el que drena la primera orina.<br />
La cápsula de Bowman está formada por varias estructuras. Tiene 2 capas u hojas que son:<br />
<br />
<br />
Hoja externa o parietal de la cápsula hay epitelio plano simple sobre una lámina basal con fibras reticulares.<br />
Hoja interna o visceral (en contacto con el glomérulo) está formada por células epiteliales modificadas<br />
llamadas podocitos. Entre ambas capas queda el espacio urinario, en el que se recoge la primera orina<br />
(ultrafiltrado).<br />
Los podocitos son células epiteliales con forma estrellada en las que el núcleo hace protrusión hacia el espacio<br />
urinario. Sus prolongaciones se ramifican en secundarias y terciarias, cuando se llaman pedicelos y están<br />
interdigitados y formando entre ellas las hendiduras o ranuras de filtración. Tienen un rico citoesqueleto y se<br />
encuentran adosadas a los capilares del glomérulo. Una de sus funciones es estimular el crecimiento del endotelio y<br />
mantener las fenestraciones, para lo que sintetizan GEGF (factor de crecimiento endotelial glomerular).<br />
Aunque todas las nefronas son similares hay diferencias histológicas:<br />
Los corpúsculos corticales están en la corteza en la zona más superior, cerca de la cápsula.<br />
<br />
Los corpúsculos yuxtamedulares (que están más cerca de la médula) se forman primero en el desarrollo<br />
embrionario y sufren antes atresia fisiológica (daño por el paso de los años).<br />
Túbulo Proximal<br />
El túbulo proximal está en continuidad con la capa parietal de la cápsula de Bowman por su polo urinario. Mide<br />
14mm y está recubierto por epitelio cúbico simple. Posee una pars convoluta y una pars recta.<br />
El túbulo contorneado proximal está exclusivamente a nivel de la corteza. El epitelio es cúbico simple con un núcleo<br />
redondeado con tendencia a situarse en el tercio basal y citoplasma eosinófilo. La membrana plasmática apical tiene<br />
un número considerable de microvellosidades, lo que permite visualizar al óptico el ribete en cepillo con glicocálix<br />
PAS+. Aparecen gran número de vesículas de endocitosis, lisosomas, cuerpos de Rhodin, RER, REL y Golgi, y<br />
abundantes zonulae occludens bastante permeables. Las células se interdigitan lateralmente. Al óptico se ven<br />
estriaciones en la zona basal del citoplasma que son repliegues de la membrana basal (bastoncitos Haiden-Hain). Son<br />
paralelas entre sí y perpendiculares a la lámina basal. Entre ellas hay mitocondrias con grandes crestas.<br />
El túbulo recto proximal se mete en la franja externa de la médula externa. El epitelio es cúbico simple con<br />
tendencia a ser algo más bajo. Las células tienen microvellosidades menos abundantes (ribete en cepillo menos<br />
desarrollado) y hay menor desarrollo de los pliegues basales y de las interdigitaciones laterales, así <strong>com</strong>o menor<br />
número de mitocondrias.<br />
Segmento Delgado
Página173<br />
El segmento delgado es la porción de la nefrona de menor diámetro. Siempre está en médula y su longitud es muy<br />
variable. En las nefronas corticales es exclusivamente descendente y después de girar da paso al túbulo recto distal,<br />
mientras que en las yuxtamedulares desciende, gira y asciende, alcanzando la médula interna. Su diámetro es de<br />
15μm. Su epitelio es plano simple y es importante no confundirlo con capilares.<br />
De forma clásica se definen cuatro tipos celulares diferentes en este segmento delgado. Las células tipo IV son<br />
mucho más planas y finas mientras que las tipo II son algo menos planas, más altas. Todas tienen desarrollo escaso<br />
de orgánulos, que destacan algo más en las II y III. Las tipo I están sólo en las nefronas de asa corta (corticales),<br />
mientras que los otros tres tipos están en determinadas regiones del segmento delgado de las nefronas<br />
yuxtamedulares (las II por todo el segmento delgado, las III en la zona de médula interna y las IV en la curvatura y<br />
segmento ascendente).<br />
Túbulo Distal<br />
El túbulo distal es ascendente y se vuelve a meter en la corteza. El de cada nefrona se aproxima a su corpúsculo de<br />
origen. Su luz es mayor que la del túbulo proximal y se observa una transición gradual en cuanto al epitelio, llegando<br />
a ser cúbico simple de nuevo. El túbulo distal tiene una pars recta, una pars convoluta y una zona entre medias que<br />
es la pars maculata (mácula densa).<br />
El túbulo recto distal nos lleva de nuevo a la corteza. Sus células sufren una transición gradual a cúbicas y tienen<br />
escasas microvellosidades, por lo que no se observa ribete en cepillo. Tampoco hay vesículas endocíticas. El núcleo<br />
redondo tiende a estar en el tercio apical de la célula y el citoplasma sigue siendo eosinófilo. Tiene desarrollo poco<br />
llamativo de orgánulos (escasos RER y Golgi) pero tienen abundantes zonulae occludens e interdigitaciones laterales.<br />
También hay estriaciones basales. Abundan los pliegues basales con mitocondrias verticales dispuestas entre ellos.<br />
La mácula densa es una porción corta del túbulo distal. Está junto al corpúsculo de origen, pegada a la arteriola<br />
aferente. Muestra un epitelio prismático con 20-30 células bastante estrechas con un gran número de núcleos muy<br />
juntos que parecen superpuestos. La membrana plasmática presenta algunas microvellosidades y un cinocilio. La<br />
polaridad está invertida (núcleo apical y Golgi basal). Estas células epiteliales carecen de los pliegues basales típicos<br />
del túbulo recto pero sí tienen prolongaciones basales, que van atraviesan la lámina basal. La mácula densa parece<br />
funcionar percibiendo cambios en la concentración de sodio (NaCl)<br />
de la orina del túbulo distal.<br />
El túbulo contorneado distal está en la corteza. Es relativamente<br />
corto con una luz amplia. Drena al conducto colector. Muestra<br />
epitelio cúbico simple con células sin ribete en cepillo con alguna<br />
microvellosidad corta. El núcleo es apical y el citoplasma menos<br />
eosinófilo que hasta ahora. Tienen pliegues basales e<br />
interdigitaciones laterales con mitocondrias verticales.<br />
Entre el túbulo colector y el túbulo contorneado distal está el túbulo<br />
reuniens. Es corto y se encuentra tapizado por epitelio plano simple.<br />
Esquema Nefrona Plegada
Página174<br />
Conducto Colector<br />
El conducto colector tiene una zona cortical revestida por epitelio simple plano que pasa a cúbico y una zona<br />
medular en la que el epitelio simple cúbico va pasando a prismático. A cada conducto colector drenan unos 10<br />
túbulos reuniens. Por último, los túbulos colectores van convergiendo para formar los conductos papilares de Bellini.<br />
En él encontramos las células principales pálidas y las intercalares oscuras.<br />
Las células principales tienen microvellosidades cortas y gruesas. Poseen un cinocilio en cuya membrana hay<br />
policistina 1 y 2; la policistina 2 es un canal para calcio y la policistina 1 es una proteína que interacciona con<br />
carbohidratos específicos y se relaciona con la adhesión célula-célula o célula-matriz. El cinocilio es un sensor<br />
mecánico del flujo y del contenido luminal. En la poliquistosis renal autosómica dominante alguno de los genes de las<br />
policistinas 1 o 2 están mutados. Se forman masivamente quistes renales y los conductos colectores acaban<br />
dilatándose. Puede llegar a repercutir a la porción distal de la nefrona. Se trata con diálisis o transplante.<br />
Estas células tienen escasas vesículas, mitocondrias pequeñas y abundante pliegues en la membrana basal, en los<br />
quepliegues encontramos acuaporinas 3 y 4. La membrana luminal es rica en canales para el sodio y potasio y la<br />
basolateral tiene abundancia de bombas Na/K. La aldosterona controla todos estos canales. Los canales de sodio<br />
también son regulados por el ANP (péptido natriurético auricular), que los inhibe, por lo que se elimina más sodio<br />
por la orina. Con estas dos hormonas se regula la reabsorción de sodio y la secreción de potasio.<br />
Estas células principales tienen abundantes acuaporinas 2 indispensables para la reabsorción de agua, regulada por<br />
vasopresina (ADH). Cuando hay niveles altos de ADH la orina excretada estará muy concentrada. Si la ADH está en<br />
niveles bajos (no se necesita conservar agua) la orina estará muy diluida. El rango de variación de eliminación de<br />
volumen de agua va de 0.5 a 23L, desde la sobrehidratación hasta la deshidratación. El volumen de agua reabsorbido<br />
en el conducto colector es independiente de sodio. Permite el 20-30% de la reabsorción de agua.<br />
En algunas situaciones la regulación por ADH no es posible. En la diabetes insípida hay déficit de ADH. Hay pérdida<br />
masiva de agua y los niveles de sodio en sangre son siempre muy altos (poliuria, polidipsia e hipernatremia) ya que<br />
los conductos colectores no reabsorben agua adecuadamente. En la diabetes insípida nefrógena no funcionan las<br />
acuaporinas 2 o el receptor para la ADH está mutado. Un exceso de ingesta de agua también es negativo para el<br />
organismo ya que puede inhibir la secreción de ADH, lo que conduciría a la formación de grandes volúmenes de orina<br />
hipoosmótica.<br />
Las células intercalares inicialmente son muy abundantes pero tienden a disminuir su número al acercarnos a la<br />
médula y no se observan en la médula interna. Poseen microvellosidades, micropliegues, interdigitaciones laterales y<br />
carecen de pliegues basales. Tienen muchas mitocondrias y se encargan de la regulación del equilibrio ácido-base.<br />
Reabsorben iones potasio y secretan hidrogeniones e iones bicarbonato.<br />
Los conductos colectores de Bellini son muy gruesos (100-200μm) y terminan abriéndose en la punta de la papila<br />
por el área cribosa. Están tapizados por epitelio prismático simple y las células tienen desarrollo notable de las<br />
uniones estrechas.
Página175<br />
Barrera de filtración<br />
En el riñón hay dos estructuras que actúan en la filtración. El sistema filtrante (glomérulos) permite un ultrafiltrado a<br />
nivel del corpúsculo. El resto de la nefrona (sistema tubular, es decir, túbulo contorneado proximal, asa de Henle,<br />
túbulo contorneado distal y tubo colector) modifica la <strong>com</strong>posición del ultrafiltrado.<br />
La barrera de filtración glomerular separa la sangre glomerular del espacio urinario. Encontramos un endotelio<br />
capilar, una lámina basal glomerular y una capa de podocitos, sobre todo la zona de los pedicelos.<br />
El endotelio tiene células muy planas con una membrana plasmática con carga negativa importante asociada a<br />
glucoproteínas. Es una de las localizaciones típicas de capilares fenestrados. En los huecos entre las células se puede<br />
observar el diafragma, que se puede perder en los preparados. Los capilares son muy permeables para moléculas<br />
pequeñas (agua, urea, glucosa y proteínas pequeñas).<br />
La lámina basal es muy gruesa (300nm), aunque se engruesa aún más en patologías <strong>com</strong>o la diabetes. Se forma por<br />
la fusión de la lámina basal subendotelial y la asociada a la capa de podocitos; así, tanto las células endoteliales <strong>com</strong>o<br />
los podocitos contribuyen a formarla y mantenerla con una tasa de renovación muy elevada y constante.<br />
Aunque su origen son dos láminas basales tiene sólo tres estratos con los <strong>com</strong>ponentes típicos (colágeno IV,<br />
fibronectina, laminina y heparán sulfato).
Página176<br />
1. Lámina rara interna: tiene heparán sulfato, lo que le da una carga negativa.<br />
2. Lámina densa: colágeno IV, laminina y fibronectina<br />
3. Lámina rara externa<br />
Las prolongaciones de los podocitos (pedicelos) están firmemente ancladas mediante integrinas, en especial α 3 β 1 , y<br />
están interdigitados dejando espacios, las ranuras de filtración, que miden unos 6nm. Al MEC muestran un material<br />
membranoso llamado diafragma de la ranura, que contiene P de adhesión nefrinas de dos tipos: Neph1 y Neph2. No<br />
es lo mismo que el diafragma de las fenestraciones.<br />
El síndrome nefrótico congénito es mortal y en él el gen que codifica para la nefrina está mutado. Hay salida masiva<br />
de proteínas (proteinuria) porque no funciona el diafragma o está ausente. Se produce distensión abdominal,<br />
hipoalbuminemia, hiperlipidemia y edema.<br />
La barrera de filtración glomerular retiene los elementos de la sangre y las grandes moléculas. En el paso a través de<br />
la barrera hay tres criterios: tamaño, carga y forma de la molécula. Las moléculas sin carga con menos de 4nm pasan<br />
de forma libre. Si no tienen carga pero superan los 8nm no pasan la barrera. Cuanta más carga negativa tenga la<br />
molécula más dificultad tendrá para atravesar la barrera. Por último, cuanto más rígida es una molécula menos<br />
facilidad tendrá para llegar al espacio urinario.<br />
Orina<br />
Los riñones reciben el 20% del gasto cardiaco, importante para filtrar la sangre. Se filtran unos 125mL/min, pero la<br />
orina final no tiene ese volumen ya que ese ultrafiltrado (180L/día) se reabsorbe en un 99%. Así, diariamente<br />
formaremos 1.5L de orina.<br />
Hay muchos productos que atraviesan la barrera glomerular, formándose un ultrafiltrado u orina primaria<br />
básicamente por diferencias de presión entre la sangre glomerular y la hidrostática del espacio urinario. Se produce<br />
una síntesis de esta orina primaria a 125mL/día (1mL/min), de los que 124mL serán reabsorbidos en otra parte de la<br />
nefrona.<br />
El ultrafiltrado pasa al túbulo proximal, en el que se reabsorbe el 70-80% de agua filtrada, aminoácidos, glucosa,<br />
vitaminas, sodio, potasio y cloro. Hay una disminución importante en el volumen de la orina. Los elementos pasan a<br />
la sangre de los capilares rectos peritubulares. Para llevar a cabo la reabsorción se da transporte pasivo<br />
(acuaporinas), transporte activo (bombas ATPasa) y endocitosis mediada por receptor. Los electrolitos reabsorbidos<br />
(<strong>com</strong>o NaCl y glucosa) crean un gradiente osmótico transcelular que hace que el agua salga de las células por<br />
ósmosis a través de la zonulae occludens y aumente la presión hidrostática del <strong>com</strong>partimento intercelular.<br />
El síndrome de Fanconi es una nefropatía hereditaria o adquirida. En el paciente no se reabsorbe un determinado<br />
elemento, <strong>com</strong>o un aminoácido o glucosa, en el túbulo contorneado proximal. Hay una menor actividad de la ATPasa<br />
Na/K dependiente de magnesio.<br />
Cuando la orina primaria llega al segmento delgado y <strong>com</strong>ienza a atravesar la rama descendente es isoosmótica<br />
respecto del plasma. Sin embargo, cuando sale por la rama ascendente será hipoosmótica.<br />
En el asa de Henle las dos ramas funcionan de forma distinta: la descendente es permeable al agua pero<br />
impermeable a la urea y al NaCl y la ascendente es impermeable al agua y más permeable al NaCl y la urea. En esta<br />
porción de la nefrona se reabsorbe el 10% del agua filtrada y una importante cantidad de sodio (20%). Los diuréticos<br />
inhiben la ATPasa Na/K, inhibiendo así la reabsorción de NaCl y aumentando la excreción de agua y NaCl.
Página177<br />
La orina entra isoosmótica respecto al plasma y sale hipoosmótica, generando un intersticio hiperosmótico. En el asa<br />
de Henle se consigue que la orina que sale esté mucho más concentrada que al principio (se produce orina<br />
hipertónica). El grado de concentración depende de la longitud del asa de Henle (segmento delgado): cuanto más<br />
largo sea más concentrada podrá ser la orina. Desde la unión corteza-médula hasta la punta de la papila se concentra<br />
cada vez más, produciéndose un gradiente de hipertonicidad.<br />
El túbulo contorneado distal es muy permeable al sodio pero impermeable al agua excepto en presencia de ADH.<br />
Cuando aparece, las células son permeables al sodio y al agua. En esta porción se reabsorbe sodio y bicarbonato y se<br />
añade a la orina iones potasio e hidrogeniones, lo que contribuye a que la orina sea un fluido ácido. Las células<br />
logran la conversión del amoniaco en ion amonio.<br />
Tejido intersticial<br />
Es el estroma que proporciona sostén al órgano y se encuentra entre túbulos y corpúsculos. Este tejido conjuntivo<br />
laxo logra el sostén por abundantes fibras colágenas y proteoglucanos. La cantidad de tejido intersticial es mayor en<br />
médula (hasta un 20% en la interna) que en la corteza (7%), donde también hay fibroblastos y APC. En la médula<br />
aparecen las células intersticiales cargadas de lípidos.<br />
Los fibroblastos se encargan de sintetizar los materiales del intersticio y de renovarlo. También sintetizan la EPO. La<br />
nefritis intersticial cursa con una reacción inflamatoria desencadenada por la activación de estos fibroblastos en la<br />
que participan macrófagos y linfocitos. Puede desarrollarse por intoxicación por fármacos nefrotóxicos (metales<br />
pesados) o por un problema inmunológico.<br />
Las células intersticiales cargadas de lípidos se sitúan en hileras paralelas a los túbulos, lo que permite un mayor<br />
grado de <strong>com</strong>pactación de la médula. Estas células son irregulares y tienen prolongaciones largas que se dirigen<br />
hacia la lámina basal y contactan con otras células. Los núcleos de las células intersticiales aparecen dispuestos en<br />
hileras transversales (<strong>com</strong>o los peldaños de una escalera). Tienen desarrollo importante del citoesqueleto de actina,<br />
del RER y del Golgi. Los filamentos de actina han sugerido que las células son capaces de contraerse, por lo que se<br />
piensa que intervienen en la regulación del flujo sanguíneo en la zona papilar, que es donde son más abundantes. Se<br />
cree que ese flujo se encuentra bajo control hormonal.<br />
Estas células tienen gran número de gotas lipídicas, fundamentalmente prostaglandina E 2 . Los lípidos están<br />
implicados en la síntesis de medulipina I, que se activa en medulipina II. La medulipina II es un vasodilatador que<br />
regula a la baja la presión arterial. También se ha sugerido que estos lípidos favorecerían la eliminación de sodio.<br />
Mesangio<br />
El mesangio es una estructura intraglomerular formada por los elementos que encontramos situados entre los<br />
capilares del glomérulo. Hay una matriz mesangial y células mesangiales.<br />
La matriz mesangial es una estructura laxa PAS+ en la que encontramos <strong>com</strong>ponentes de las láminas basales <strong>com</strong>o<br />
colágeno IV, V, VI, fibronectina, laminina y perlecán. Es más fibrosa que la lámina basal típica y esta matriz la<br />
sintetizan las células mesangiales.<br />
Las células mesangiales tienen núcleos densos de gran tamaño. Tienen prolongaciones con actina y miosina muy<br />
abundantes. Sufren mitosis. También destaca la presencia de abundantes lisosomas y pigmentos <strong>com</strong>o la lipofuscina.<br />
Las prolongaciones llegan hasta la lámina basal y penetran alcanzando las fenestraciones de los capilares. En<br />
realidad, estas células proceden de pericitos de la capa visceral de la cápsula de Bowman que se modifican. Son<br />
contráctiles, capaces de fagocitar de forma activa y también secretan.<br />
Dan sostén al ovillo de capilares (esqueleto intraglomerular). Sintetizan los <strong>com</strong>ponentes de la matriz mesangial,<br />
renuevan la lámina basal y fagocitan los elementos que podían quedar retenidos en la barrera de filtración
Página178<br />
glomerular. Sintetizan prostaglandinas, interleucinas (sobre todo IL1), PDGF y endotelinas. Las endotelinas actúan<br />
sobre la pared de las arteriolas aferentes y eferentes y permiten su contracción. Tienen receptores para la<br />
angiotensina II, que lleva a una importante vasoconstricción. Así, una función demostrada de estas células es que<br />
bajo el control de la angiotensina II regulan el flujo sanguíneo que entra y sale del glomérulo.<br />
Intervienen en eliminar los <strong>com</strong>puestos retenidos o proteínas del <strong>com</strong>plemento que han acudido por una reacción<br />
inflamatoria. Así, la acumulación de inmunoglobulinas y moléculas del <strong>com</strong>plemento en la matriz mesangial induce la<br />
producción de citosinas por las células mesangiales. Producen una reacción inflamatoria que puede llevar a la<br />
oclusión de la luz capilar (glomerulonefritis).<br />
Aparato yuxtaglomerular<br />
Este aparato es un órgano endocrino que encontramos dentro del propio riñón. Está constituido por varías<br />
estructuras y se localiza en el polo vascular de los corpúsculos, delimitado por la mácula densa, las arteriolas<br />
aferente y eferente, el propio mesangio y glomérulo. Funciona de forma que proporciona información<br />
(retroalimentación tubuloglomerular) sobre la orina que se está formando. Lleva a una autorregulación del flujo<br />
sanguíneo renal y por tanto la filtración glomerular de la sangre.<br />
Está formado por:<br />
<br />
<br />
<br />
La mácula densa o células modificadas del túbulo contorneado distal<br />
El mesangio extraglomerular con células mesangiales extraglomerulares, del lacis o de Goormaghtigh<br />
(controlan la filtración en la superficie)<br />
Células mioepiteliales de Ritter o yuxtaglomerulares.<br />
Las células más altas del túbulo distal se asientan sobre una lámina basal discontinua. Presentan prolongaciones<br />
basales que se meten en la lámina basal y alcanzan las células yuxtaglomerulares. La función de la mácula densa es<br />
percibir cambios en la concentración de sodio en la orina que pasa por el túbulo distal.<br />
El mesangio extraglomerular es un espacio en contacto con el mesangio propiamente dicho, dentro del glomérulo.<br />
Las células mesangiales extraglomerulares están unidas fuertemente por uniones GAP y también se unen con las<br />
células del mesangio. Estas células también contribuyen en la percepción de los niveles de sodio de la orina.<br />
Las células yuxtaglomerulares son miocitos modificados de la túnica media de las arteriolas, fundamentalmente de<br />
la aferente pero no de manera exclusiva. Adquieren un aspecto de células epitelioides redondeadas. Se tiñen poco y<br />
tienen desarrollado el RER y Golgi. Hay gránulos tipo I y tipo II que en realidad equivalen a las dos fases de síntesis de<br />
la hormona renina.
Página179<br />
La renina permite convertir el angiotensinógeno en angiotensina I. Cuando alcanza los capilares pulmonares se<br />
obtiene angiotensina II. La angiotensina II es un vasoconstrictor muy potente que regular la homeostasis de sodio y<br />
hemodinámica renal. Estas células permiten la síntesis, almacenamiento y secreción del vasoconstrictor, por lo que<br />
regulan el flujo sanguíneo local. Eleva la presión sanguínea sistólica y diastólica. Además, la angiotensina II estimula<br />
la síntesis de aldosterona, que interviene en la rama ascendente del asa de Henle y en el túbulo colector y aumenta<br />
la reabsorción de sodio y agua. Aumenta así el volumen sanguíneo y la tensión arterial. La angiotensina II también<br />
influye en la reabsorción de sodio del túbulo contorneado distal y en la estimulación de la síntesis de ADH, que<br />
aumenta la reabsorción de sodio y agua.<br />
Las células yuxtaglomerulares funcionan <strong>com</strong>o barorreceptores de la presión arterial. Cuando la presión se eleva se<br />
inhibe la liberación de renina, que se acumula en forma de gránulos. También interviene la mácula densa, que<br />
percibe cambios en el sodio. Cuando disminuyen los niveles de sodio se estimula la secreción de renina. El SNS regula<br />
la renina (NE y dopamina estimulan la secreción de renina).<br />
Equilibrio hídrico<br />
Hay pérdidas constantes de agua por la piel (evaporación y espiración), riñón, heces y orina. Los riñones son<br />
esenciales en el control de la pérdida de agua y en el equilibrio del agua en el cuerpo. Así, la dará más hiperosmótica<br />
o más hipoosmótica según las necesidades. La excreción renal es independiente de otras sustancias.<br />
En el equilibrio hídrico interviene al angiotensina II, que estimula la reabsorción del sodio y agua en el túbulo<br />
contorneado proximal. Se produce en situaciones de descenso del volumen sanguíneo (hipovolemia). Se activa la<br />
liberación de renina y aumenta la angiotensina.<br />
La aldosterona actúa estimulando la reabsorción del sodio en la rama ascendente del asa de Henle, túbulo<br />
contorneado distal y túbulo colector. La aldosterona aumenta cuando han aumentado los niveles de potasio o<br />
angiotensina II previamente.<br />
El factor natriurético auricular estimula la excreción de sodio y agua (inhibe los canales de sodio). Como<br />
consecuencia de su liberación se inhibe la secreción de ADH. La urodilatina es bastante similar al anterior y se<br />
sintetiza y segrega por las células de túbulo contorneado distal y colector. Inhibe la reabsorción del agua y sodio a<br />
nivel del túbulo colector medular.<br />
La vasopresina (ADH) se sintetiza en la neurohipófisis. Cuando tenemos hipovolemia se segrega y aumenta la<br />
permeabilidad al agua del túbulo colector, aumentando la reabsorción de agua. Es independiente del sodio.<br />
Formación de la orina<br />
La orina que entra en el túbulo proximal es isoosmótica. Se adentra en la rama descendente del asa de Henle, que es<br />
muy poco permeable al agua y poco al sodio gracias al intersticio hiperosmótico, y cuando llega al giro del asa ha<br />
pasado a ser hiperosmótica. Comienza a ascender y se absorbe gran cantidad de sodio pasando a una orina<br />
hipoosmótica. En la zona más cortical se reabsorbe sodio y el agua se reabsorberá en presencia de vasopresina.<br />
Tanto en la porción cortical <strong>com</strong>o en la medular se sigue reabsorbiendo agua, gracias a lo que la orina sufre un<br />
proceso de concentración y se vuelve hiperosmótica.<br />
Este proceso es posible gracias a la presencia de dos sistemas paralelos en los que fluye fluido con direcciones<br />
opuestas (proceso multiplicador e intercambiador de corriente). Así tenemos dos tubos con fluidos en direcciones<br />
opuestas que permite que se intercambien sustancias y calor. Por una parte tenemos los tubos de la nefrona y los<br />
colectores, que van paralelos a los vasos rectos con la sangre fluyendo en dirección opuesta a la orina. Esto permite<br />
un intercambio de <strong>com</strong>puestos, fundamentalmente electrolitos y agua. El intersticio se vuelve hiperosmótico en la
Página180<br />
zona medular. Según profundizamos en la médula la concentración de sodio aumenta. Esto crea un gradiente en<br />
tanto que el agua es eliminada por los vasos rectos.<br />
La orina que se obtiene al final es hipertónica con pH 5.5. A ella se añaden <strong>com</strong>puestos finales <strong>com</strong>o urea, ácido<br />
úrico, amonio y creatinina en este proceso multiplicador.<br />
VÍAS URINARIAS<br />
Cálices menores y mayores<br />
Los cálices menores son cuatro y desembocan en un cáliz mayor. Tienen una mucosa con urotelio que se asienta<br />
sobre una lámina propia que muestra una zona superficial más densa y una zona profunda con conjuntivo más laxo.<br />
Además, la lámina propia es rica en fibras elásticas y tiene linfocitos y nódulos. Se asienta sobre una muscular con<br />
dos estratos, uno interno longitudinal y uno externo circular. Por último tendríamos la adventicia.<br />
URÉTER<br />
El uréter es una estructura par de unos 20-30cm y 5-7mm de diámetro que está incluido en la grasa retroperitoneal.<br />
Su porción proximal se dilata y forma la pelvis renal; la distal está dentro de la vejiga. En la nefrolitiasis aparecen<br />
cálculos renales por sales cálcicas, ácido úrico o acetato de magnesio-amonio por cristalización. Producen un dolor<br />
muy agudo e intenso al contraerse el uréter obstruido.<br />
Su epitelio es uroepitelio o epitelio de transición y se asienta sobre una lámina basal fibroelástica. Hay una zona de<br />
músculo liso que se estructura en 2-3 capas que son una longitudinal interna, otra circular externa y en el tercio<br />
inferior aparece una última más externa de carácter longitudinal. Por fuera, hay una capa adventicia.<br />
VEJIGA<br />
Es un receptáculo que aumenta o disminuye de tamaño (distensible) y con un tamaño de 14x10x7cm. Está en la<br />
pelvis por detrás de la sínfisis del pubis.<br />
La vejiga tiene una pared que cambia de forma y tamaño según el momento fisiológico en el que esté (esto permite<br />
adaptarse a un mayor o menor volumen). Así, puede aparecer delgada y lisa o mucho más gruesa y con pliegues. Esta<br />
zona es distinta desde el punto de vista embriológico.<br />
En cambio, existe una zona llamada trígono, el espacio delimitado por los dos orificios ureterales y el uretral interno,<br />
cuya superficie es siempre lisa y tiene un grosor constante. Esto se explica porque la vejiga deriva de la cloaca, pero<br />
el trígono proviene de los conductos mesonéfricos.<br />
Tanto vejiga <strong>com</strong>o uréter tienen tres capas (es muy similar al tercio distal del uréter). La parte más distal de uréteres<br />
y vejiga están asociados a una serosa, el peritoneo. Las capas son cada vez más gruesas conforme vamos hacia distal.<br />
La mucosa está formada por urotelio asentado sobre una lámina propia y según la mucosa esté más o menos<br />
plegada puede ser más o menos lisa (muy lisa y aplanada en distensión).<br />
El urotelio es estratificado y tiene capas variables (2-3 hasta 7-8) según el momento en el que se observe la<br />
estructura y según la zona, aplanándose en distensión. Además, estas células tienen gran desarrollo de las zonulae<br />
occludens y son muy herméticas.
Página181<br />
Distinguimos:<br />
<br />
<br />
<br />
Células basales bastante altas cúbicas o cilíndricas<br />
Células intermedias poligonales<br />
Células superiores en paraguas o sombrilla: son más redondeadas,<br />
suelen ser binucleadas y se tiñen menos. Tienen gran desarrollo del<br />
citoesqueleto y se observa que tienen su superficie en cúpula. Su<br />
membrana apical presenta ciertas condensaciones en la membrana<br />
que al MET se aprecia que son placas rígidas y gruesas, más densas, en<br />
conexión con filamentos de actina; así permiten la distensibilidad de la<br />
vejiga. Encontramos gran cantidad de estructuras con forma de<br />
vesículas alargadas y una superficie muy irregular. Cuando no se necesita tanta superficie se retira<br />
membrana en forma de esas vesículas y las placas se invaginan y se forman vesículas fusiformes, dando un<br />
perfil celular muy irregular. En la distensión, las vesículas se fusionan con la célula para aumentar la<br />
superficie de la membrana y las células adquieren un perfil aplanado.<br />
La lámina propia tiene una zona superficial es más densa que la profunda, que es laxa. Hay abundantes linfocitos,<br />
nódulos y fibras elásticas.<br />
En el estrato muscular hay una capa longitudinal interna (espiral laxa) y una circular externa (espiral apretada). En la<br />
parte más distal de las dos estructuras hay un tercer estrato interno longitudinal. Los miocitos no se disponen<br />
independientes sino de forma helicoidal. Cuando se contrae todo el estrato muscular se produce la evacuación de la<br />
orina. La orina se transporta desde el seno renal por ondas peristálticas espontáneas que permiten el avance hacia la<br />
vejiga.<br />
Hay una serie de fibras musculares que se organizan de forma circular para evitar el reflujo de orina y que conforman<br />
el manguito o músculo detrusor de la vejiga; actúa a modo de esfínter.<br />
La vejiga se dispone de forma que al llenarse los orificios ureterales quedan <strong>com</strong>primidos, lo que evita un reflujo de<br />
la orina. Cuando la vejiga se vacía la contracción muscular también <strong>com</strong>prime los orificios ureterales, evitándose<br />
también el reflujo en la micción. Las infecciones de las vías urinarias son muy frecuentes por el reflujo de la orina<br />
hacia los uréteres.<br />
La capa muscular tiene una irrigación importante e inervación simpática y parasimpática. Si se extirpa la del uréter<br />
las ondas peristálticas se siguen produciendo. Las fibras simpáticas del plexo vesical e inervan la musculatura de los<br />
vasos, mientras que las parasimpáticas del plexo vesical inervan la musculatura de la pared vesical y llevan la orden<br />
del reflejo de la micción. En la micción se requiere un elemento aferente que va de la mano de fibras sensitivas. Las<br />
parasimpáticas motoras son la porción eferente del reflejo.<br />
URETRA<br />
La uretra es un tubo fibromuscular que permite la evacuación de la orina. La de la mujer es muy corta y la del varón<br />
es muy larga y permite también la salida del esperma. La uretra del varón se divide en tres porciones, la prostática,<br />
membranosa y esponjosa o peneana.<br />
La uretra prostática mide 3-4cm y recorre el interior de la próstata. A ella llegan las secreciones de los conductos<br />
eyaculadores y las secreciones de la propia próstata. Presenta una mucosa con urotelio y lámina propia con<br />
conjuntivo laxo muy vascularizado.
Página182<br />
La uretra membranosa mide 1cm y atraviesa el diafragma urogenital de la pelvis alcanzando el periné. El músculo<br />
esquelético del diafragma urogenital forma el esfínter externo. Su mucosa tiene epitelio cilíndrico<br />
pseudoestratificado.<br />
La porción peneana mide 15cm y termina abriéndose a la zona del glande en la fosita navicular. Aquí llega la<br />
secreción de las glándulas de Cowper y de Littré. Tiene epitelio prismático estratificado o pseudoestratificado que en<br />
el glande se convierte en plano estratificado. Carece de musculatura y la mucosa se asienta directamente sobre el<br />
cuerpo esponjoso.<br />
La uretra femenina mide poco 3-4cm y se abre al vestíbulo vaginal justo detrás del clítoris. Perfora el diafragma<br />
urogenital de forma que hay un esfínter uretral externo. La mucosa forma pliegues longitudinales. Inicialmente está<br />
tapizada por un urotelio que se modifica hasta llegar a ser un epitelio plano estratificado. Se asienta sobre una<br />
lámina propia muy rica en elásticas con un plexo venoso muy desarrollado que recuerda al del cuerpo esponjoso de<br />
los varones. La capa muscular tiene un estrato interno longitudinal de músculo liso y uno externo longitudinal de<br />
miocitos esqueléticos. Ambas capas actúan <strong>com</strong>o esfínter, uno voluntario y el otro involuntario.<br />
Las glándulas uretrales están en la porción proximal de la uretra y son pequeñas. Hay unas glándulas homólogas a las<br />
prostáticas del varón, las parauretrales. Su secreción no se vierte a la uretra, sino que los conductos se abren al<br />
vestíbulo vaginal a ambos lados del orificio uretral externo. Tienen una secreción alcalina.
Página183<br />
ANEXO<br />
EJEMPLO EXAMEN SISTEMA CIRCULATORIO Y SISTEMA RESPIRATORIO<br />
Desarrollo:<br />
Test:<br />
1) Diferencias entre arteria femoral y vena femoral<br />
2) ¿Qué es y dónde se localiza el espacio de Reinke?<br />
El área de Kiesselbach la encontramos en:<br />
a) La cavidad orofaríngea<br />
b) La bifurcación bronquial carinal<br />
c) La laringe<br />
d) La tráquea<br />
e) Ninguna es correcta<br />
Respecto al sistema cardiocirculatorio, es cierto:<br />
a) La arteria umbilical presenta una gruesa lámina elástica interna<br />
b) La túnica íntima de los conductos linfáticos no contiene células mesoteliales<br />
c) Se consideran venas de mediano calibre a las venas de calibre inferior a 1cm de diámetro<br />
d) Son correctas b) y c)<br />
e) Todas son correctas<br />
Respecto a las arterias es correcto:<br />
a) Las células endoteliales participan en el control de la coagulación<br />
b) Las arterias que se originan en el cayado aórtico son las arterias musculares<br />
c) Las arteriolas presentan de 4 a 10 capas de miocitos lisos<br />
d) Son correctas a) y c)<br />
e) Todas son correctas<br />
Respecto al sistema respiratorio es verdadero:<br />
a) Las células neuroendocrinas pulmonares pertenecen al sistema APUD<br />
b) Las células neuroendocrinas suelen ser positivas a la IHQ de cromogranina<br />
c) Las células neuroendocrinas son aproximadamente el 30% del árbol bronquial<br />
d) Son correctas a) y b)<br />
e) Todas son correctas<br />
Respecto al corazón es cierto que:<br />
a) La superficie ventricular superior es rugosa para la inserción de las cuerdas tendinosas<br />
b) Las cuerdas tendinosas se nutren por difusión de la sangre de las cámaras ventriculares<br />
c) La parte membranosa del tabique interventricular ocupa el tercio inferior del mismo<br />
d) Son correctas a) y b)<br />
e) Todas son correctas
Página184<br />
Respecto al corazón es cierto que:<br />
a) Las valvas cardíacas son avasculares<br />
b) Las cuerdas tendinosas sólo tienen vascularización capilar<br />
c) Las proyecciones de miocardio y endocardio hacia la luz ventricular forma los músculos papilares<br />
d) Las arterias musculares tienen una capa media formada casi exclusivamente por músculo liso<br />
e) El endocardio se continúa con la túnica íntima de los vasos sanguíneos<br />
Respecto al sistema respiratorio es falso:<br />
a) Las células basales del epitelio bronquial no alcanzan la superficie luminal<br />
b) Los neumocitos tipo I presentan uniones ocluyentes entre ellas<br />
c) Las paredes de los bronquiolos y sus ramas carecen de cartílago<br />
d) Las células en cepillo de la tráquea son el 20% de la población celular<br />
e) Las células mucosas son más escasas en los bronquiolos y ausentes en los terminales y respiratorios<br />
Respecto al sistema respiratorio, es cierto:<br />
a) Los pliegues ventriculares de la laringe presentan epitelio cúbico seudoestratificado<br />
b) Los conductos alveolares presentan pared propia con miocitos esqueléticos<br />
c) Las glándulas bronquiales submucosas son glándulas seromucosas<br />
d) Son correctas a) y c)<br />
e) Todas son correctas<br />
Señala la respuesta correcta:<br />
a) Los neumocitos tipo II presentan en su cara luminal microvellosidades<br />
b) Los sacos alveolares terminan en los conductos alveolares<br />
c) Los cuerpos multilaminares se localizan en los neumocitos tipo I<br />
d) Son correctas b) y c)<br />
e) Todas son correctas<br />
Respecto al capilar sinusoide es correcto que:<br />
a) Presentan una trayectoria tortuosa y diámetro amplio<br />
b) Sus células endoteliales forman una capa continua<br />
c) EL citoplasma de las células endoteliales muestran fenestraciones múltiples con diafragmas<br />
d) Son correctas a) y b)<br />
e) Todas son correctas<br />
EXÁMENES ORGANOGRAFÍA USP-CEU<br />
PREGUNTAS DESARROLLO<br />
Definición de ORGANOGRAFÍA MICROSCÓPICA <strong>HUMANA</strong><br />
Concepto de ÓRGANO<br />
Concepto de TEJIDO<br />
Organización básica de los ÓRGANOS MACIZOS<br />
Organización básica de los ÓRGANOS HUECOS<br />
Concepto de PARÉNQUIMA<br />
Concepto de ESTROMA<br />
¿Tienen los órganos huecos parénquima y estroma? Explica por qué SÍ o por qué NO.<br />
Estructura histológica de la piel. Tipos de células de la epidermis. Disposición, características de las mismas,<br />
similitudes y diferencias.<br />
Estructura histológica del folículo piloso.<br />
Tipos de glándulas cutáneas, características de las mismas, diferencias y similitudes.<br />
Estructura histológica de la uña.<br />
Estructura histológica de la hipófisis y células características de cada región.
Página185<br />
Estructura histológica de la glándula tiroides.<br />
Estructura histológica de la glándula paratiroides<br />
Estructura histológica de la glándula pineal<br />
Estructura histológica de la glándula suprarrenal<br />
Estructura histológica del ovario<br />
Estructura histológica de la trompa de Falopio<br />
Estructura histológica del útero<br />
PREGUNTAS TIPOS TEST<br />
La presencia de mitosis en la capa granulosa de la epidermis:<br />
a) Es prácticamente sinónimo de patología<br />
b) Es fisiológico<br />
c) No es habitual pero no es patológico<br />
d) Todas son correctas<br />
e) Ninguna es correcta<br />
Señala cual de estas afirmaciones acerca del lobulillo clásico hepático humano es verdadera:<br />
a) Los canalículos biliares vierten su contenido a un vaso central<br />
b) La sangre fluye en dirección centrípeta<br />
c) La triada portal está delimitado por el espacio de Mall<br />
d) Se encuentra delimitado por tabiques de tejido conjuntivo<br />
e) Existe una arteria central de gran calibre<br />
En la matriz extracelular de la Médula Ósea existe:<br />
a) Colágeno tipo I<br />
b) Colágeno tipo III<br />
c) Laminina<br />
d) Son correctas b) y c)<br />
e) Todas son correctas<br />
En la médula ósea,…<br />
a) El calibre de las vénulas es menor que el arteriolar<br />
b) La presión hidrostática de los capilares es elevada<br />
c) Los capilares son de tipo sinusoide<br />
d) Son correctas b) y c)<br />
e) Todas son correctas<br />
Con respecto a los hepatocitos, según el modelo de acino hepático, los localizados en la zona 1:<br />
a) Presentan mayor cantidad de enzimas implicadas en la síntesis y degradación de glucógeno<br />
b) Se encuentran más oxigenados<br />
c) Presentan mayor cantidad de enzimas implicadas en la síntesis de ácidos grasos<br />
d) a) y b) son ciertas<br />
e) Todas son ciertas<br />
Respecto a la médula ósea (MO) ...<br />
a) El tipo MO “gris” es habitual en recién nacidos<br />
b) El tipo MO “amarilla” es reversible a roja<br />
c) El tipo MO “roja” puede pasar a amarilla o gris según las circunstancias<br />
d) Son correctas b) y c)<br />
e) Todas son correctas
Página186<br />
Respecto a los conductos genitales masculinos…<br />
a) Los túbulos seminíferos drenan a la rete testis mediante los tubos rectos<br />
b) La rete testis drena al epidídimo mediante los conductillos eferentes<br />
c) El epitelio del epidídimo es pseudoestratificado, al igual que el del conducto deferente<br />
d) Son correctas a) y b)<br />
e) Todas son correctas<br />
Señala cual de las siguientes afirmaciones es falsa<br />
a) El los espacios de Disse del hígado se genera la linfa primitiva<br />
b) El parénquima hepático está inervado por el sistema nervioso vegetativo<br />
c) Los primeros capilares linfáticos se localizan en el espacio porta<br />
d) La membrana del polo biliar del hepatocito es lisa<br />
e) En los hepatocitos de ancianos a menudo aparecen acúmulos de lipofuxina<br />
Con respecto a las células folículo-estrelladas de la hipófisis:<br />
a) Son cromófobas<br />
b) Aparecen en la pars distalis de la adenohipófisis<br />
c) Son abundantes en la pars intermedia donde forman folículos<br />
d) a) y b) son verdaderas<br />
e) Todas son verdaderas<br />
De los siguientes conductos ¿cuál está tapizado por un epitelio cúbico con microvellosidades o borde en cepillo?<br />
a) El túbulo contorneado proximal<br />
b) El túbulo contorneado distal<br />
c) El túbulo colector cortical<br />
d) El túbulo colector medular<br />
e) La porción ascendente gruesa<br />
¿Cual de las siguientes afirmaciones con respecto al páncreas es falsa?:<br />
a) En la parte exocrina, la porción secretora <strong>com</strong>ienza en los conductos intercalares<br />
b) Los conductos interlobulillares puede presentar células caliciformes<br />
c) Los enterocitos activan las proenzimas del jugo pancreático a través de enterocinasas localizadas en su glicocalix<br />
d) En la porción exocrina, las células tienen escaso RER y ribosomas pero existe abundante REL<br />
e) La secreción exocrina del páncreas está sometida a control nervioso y endocrino<br />
El endometrio:<br />
a) Presenta glándulas alveolares<br />
b) Presenta células cilíndricas estereociliadas<br />
c) Presenta glándulas endocrinas<br />
d) Son correctas a) y c)<br />
e) Ninguna es correcta<br />
Las células parafoliculares del tiroides:<br />
a) Se localizan en el espacio de Disse<br />
b) Contactan directamente con la luz del folículo<br />
c) Forman una capa continua<br />
d) También se llaman células claras aunque tienen gránulos densos en su citoplasma basal<br />
e) Todas son verdaderas<br />
En los islotes de Langerhans:<br />
a) Las células alfa secretan insulina y son pequeñas y centrales<br />
b) Las células beta secretan glucagón y son grandes y acidófilas<br />
c) Las células delta son las más abundantes en el islote<br />
d) a) y b) son verdaderas<br />
e) Ninguna es verdadera
Página187<br />
¿Dónde se encuentran las células laci?<br />
a) Testículo<br />
b) Epidídimo<br />
c) Glándula de Cowper<br />
d) Vesícula seminal<br />
e) Ninguna es correcta<br />
En el lóbulo anterior de la hipófisis:<br />
a) Pituicito es el nombre genérico para las células secretoras de hormonas<br />
b) Se observa la aparición de cuerpos de Herring<br />
c) Aparecen múltiples axones del tracto diencéfalo-hipofisario<br />
d) Todas son falsas<br />
e) Todas son verdaderas<br />
Las células matriciales se localizan en:<br />
a) Las glándulas sebáceas<br />
b) Las glándulas sudoríparas apoecrinas<br />
c) La piel lampiña<br />
d) El bulbo piloso<br />
e) Ninguna es correcta<br />
En el hombre…<br />
a) En el túbulo colector renal existen células principales e intercalares<br />
b) La pelvis renal está tapizada por urotelio<br />
c) Los cuerpos amiláceos se observan en la próstata<br />
d) Son correctas b) y c)<br />
e) Todas son correctas<br />
¿Cuáles de las siguientes células en la adenohipófisis se teñirían de rojo con H/E?<br />
a) Tirotropas, somatotropas y corticotropas<br />
b) Lactotropas y somatotropas<br />
c) Tirotropas, gonadotropas y corticotropas<br />
d) Gonadotropas y somatotropas<br />
e) Gonadotropas y corticotropas<br />
Con respecto a los folículos tiroideos:<br />
a) Presentan un epitelio alto cuando la glándula está activa<br />
b) Cuando el epitelio está activado presenta menos microvellosidades<br />
c) Su epitelio es siempre cúbico<br />
d) Cuando la glándula está en reposo las células son cilíndricas altas<br />
e) a) y b) son ciertas<br />
Dentro de las glándulas paratiroideas:<br />
a) las células principales claras tienen muchas mitocondrias<br />
b) Las células principales oscuras son PASc)<br />
Las células oxífilas son células en plena actividad secretora<br />
d) Las células principales claras y oscuras son dos tipos celulares independientes<br />
e) Todas son verdaderas
Página188<br />
¿Cual de las siguientes afirmaciones acerca de la glándula suprarrenal es falsa?:<br />
a) La corteza es de origen mesodérmico<br />
b) Vasos y fibras nerviosas entran a través de tabiques conjuntivos<br />
c) En el estroma hay ausencia de fibras reticulares<br />
d) La médula tiene un doble aporte sanguíneo arterial y venoso<br />
e) Sólo la secreción de la médula tiene control nervioso<br />
En la médula de la glándula suprarrenal:<br />
a) Las células secretoras de adrenalina son las menos frecuentes<br />
b) Las células que secretan noradrenalina tienen gránulos con material irregular<br />
c) Sus células son autofluorescentes<br />
d) Todas son verdaderas<br />
e) b) y c) son verdaderas<br />
En la glándula suprarrenal, la hormona hipofisaria ACTH es necesaria para que se mantenga:<br />
f) La zona fascicular<br />
g) La zona reticular<br />
h) La zona glomerulosa<br />
i) a) y b) son ciertas<br />
j) Todas son ciertas<br />
La eleidina es una proteína presente en:<br />
a) Corteza adrenal<br />
b) Piel gruesa<br />
c) Glándulas de Moll<br />
d) Cérvix uterino<br />
e) Ninguna es correcta<br />
En el estudio de la vesícula seminal existe un detalle importante para diferenciarla de la próstata, ¿cuál?<br />
a) La presencia de microvellosidades<br />
b) La presencia de pigmento<br />
c) La presencia de vacuolas subnucleares<br />
d) La presencia de estratificación nuclear<br />
e) Ninguna es correcta<br />
Con respecto a los pinealocitos:<br />
a) Sus prolongaciones se extienden sobre los vasos de los tabiques<br />
b) Además de hormonas peptídicas contienen proteinas transportadoras o neurofisinas<br />
c) Se localizan preferentemente hacia la periferia de la glándula<br />
d) Todas son verdaderas<br />
e) a) y b) son verdaderas<br />
El colédoco:<br />
a) Se localiza en el hígado en la triada portal<br />
b) Se localiza en el lateral del lobulillo hepático<br />
c) Presenta la misma estructura básica de la vesícula biliar<br />
d) Presenta un epitelio cúbico<br />
e) No presenta miocitos alrededor<br />
La mácula densa se localiza en:<br />
a) El asa de Henle<br />
b) El túbulo contorneado distal<br />
c) El túbulo contorneado proximal<br />
d) El conducto colector<br />
e) Ninguna es correcta
Página189<br />
La fase de reposo del folículo piloso se denomina:<br />
a) Anageno<br />
b) Catageno<br />
c) Telogeno<br />
d) Paquigeno<br />
e) Introgeno<br />
La capa más externa de la cápsula testicular es:<br />
a) La túnica íntima<br />
b) La túnica vascular<br />
c) La túnica albugínea<br />
d) La túnica vaginal<br />
e) Ninguna es correcta<br />
Las glándulas bulbouretrales también se denominan:<br />
a) Glándulas de Tyson<br />
b) Glándulas de Bartholin<br />
c) Glándulas de Moll<br />
d) Glándulas de Littre<br />
e) Ninguna es correcta<br />
Señala la relación correcta:<br />
a) Glándula sudorípara apocrina – Feromonas<br />
b) Glándula sudorípara ecrina – Acrosiringio<br />
c) Glándula sudorípara apoecrina – Axila<br />
d) Todas son correctas<br />
e) Son correctas a) y b)<br />
El epitelio que tapiza la porción externa de los senos galactóforos es de tipo:<br />
a) Plano estratificado no queratinizado<br />
b) Plano estratificado queratinizado<br />
c) Cilíndrico estratificado<br />
d) Plano simple<br />
e) Ninguna es correcta<br />
Las glándulas de Montgomery se localizan en:<br />
a) El cérvix uterino<br />
b) La vagina<br />
c) La areola mamaria<br />
d) El vestíbulo vulvar<br />
e) Ninguna es correcta<br />
Las glándulas tubulares ramificadas de Littre se localizan en:<br />
a) La próstata<br />
b) El uréter<br />
c) El riñón<br />
d) La uretra<br />
e) Son correctas b) y d)
Página190<br />
En la mucosa endometrial, señala lo incorrecto:<br />
a) En la fase proliferativa existen vacuolas subnucleares en el epitelio glandular.<br />
b) En la fase secretora existen numerosos vasos en espiral en el estroma<br />
c) La fase estrogénica es la primera fase del ciclo<br />
d) En la fase secretora existen abundantes mitosis<br />
e) En la fase secretora hay mayor influencia de progesterona<br />
La transición entre un epitelio cilíndrico simple de secreción mucosa a otro de tipo plano estratificado no<br />
queratinizado se produce en:<br />
a) Glande del pene<br />
b) Próstata<br />
c) Cérvix uterino<br />
d) Pelvis renal- pielocalicial<br />
e) Rete testis<br />
Entre las siguientes, ¿cuál no es una capa de la vagina?<br />
a) La mucosa<br />
b) La submucosa<br />
c) La muscular<br />
d) La adventicia<br />
e) Todas son capas de la vagina<br />
Respecto a la Glándula mamaria, señala la respuesta correcta:<br />
a) Se considera una glándula sebácea modificada<br />
b) Entre dos lobulillos existe tejido conjuntivo laxo<br />
c) Existe músculo liso y esquelético en la areola<br />
d) Son correctas b) y c)<br />
e) Ninguna es correcta<br />
El ovario está tapizado externamente por un epitelio denominado:<br />
a) Epitelio germinal<br />
b) Epitelio sustentacular<br />
c) Epitelio interfásico<br />
d) Epitelio transicional<br />
e) Epitelio folicular<br />
La capa parietal de la cápsula de Bowman lo forma:<br />
a) Un epitelio cilíndrico ciliado<br />
b) Un epitelio cúbico estratificado<br />
c) Un epitelio cilíndrico pseudoestratificado<br />
d) Un epitelio cúbico con estereocilios<br />
e) Ninguna es correcta<br />
De las siguientes células, ¿Cuáles son neuronas bipolares?:<br />
a) Las células olfatorias<br />
b) Las células ciliadas de las máculas<br />
c) Las neuronas gigantes del ganglio raquídeo<br />
d) Las células piramidales de la corteza cerebral<br />
e) Todas son falsas
Página191<br />
Las glándulas de Skene también se denominan…<br />
a) Glándulas bulbouretrales<br />
b) Glándulas parauretrales<br />
c) Glándulas bulbares menores<br />
d) Glándulas vestibulares mayores<br />
e) Ninguna es correcta<br />
Las pirámides renales terminan en los cálices en puntos denominados:<br />
a) Puntos orificiales<br />
b) Membrana de fusión<br />
c) Lámina cribosa<br />
d) Puntos de anclaje<br />
e) Ninguna es correcta<br />
El <strong>com</strong>ponente mayoritario del lóbulo posterior de la hipófisis:<br />
a) Son los pituicitos<br />
b) Son axones del tracto diencéfalo-hipofisario<br />
c) Son los cuerpos neuronales secretores de hormonas<br />
d) Son las células de Schwann<br />
e) Son los oligodendrocitos<br />
Señala cual de estas afirmaciones es falsa:<br />
a) El esófago de Barret consiste en una metaplasia del epitelio<br />
b) En la artritis reumatoide la membrana sinovial se engrosa y se denomina pannus<br />
c) Una de las características histológicas de la pancreatitis es la fibrosis<br />
d) La diabetes mellitus se debe al mal funcionamiento de las células beta del pancreas<br />
e) En la cirrosis biliar primaria el primer cambio que se produce es la fibrosis<br />
Señala la respuesta correcta<br />
a) La diabetes mellitus puede dar lugar a alteraciones vasculares a nivel de la retina<br />
b) Las células madre nunca pierden su pluripotencialidad<br />
c) En el envejecimiento la disminución de la estatura sólo se debe al encorvamiento de la columna<br />
d) En la cirrosis biliar primaria no existe respuesta inflamatoria<br />
e) La membrana sinovial está formada por fibras, es acelular<br />
Señala cual de estas afirmaciones acerca del lobulillo clásico hepático humano es verdadera:<br />
f) Los canalículos biliares vierten su contenido a un vaso central<br />
g) La sangre fluye en dirección centrípeta<br />
h) La triada portal está delimitado por el espacio de Mall<br />
i) Se encuentra delimitado por tabiques de tejido conjuntivo<br />
j) Existe una arteria central de gran calibre<br />
Con respecto a los hepatocitos, según el modelo de acino hepático, los localizados en la zona 1:<br />
f) Presentan mayor cantidad de enzimas implicadas en la síntesis y degradación de glucógeno<br />
g) Se encuentran más oxigenados<br />
h) Presentan mayor cantidad de enzimas implicadas en la síntesis de ácidos grasos<br />
i) a) y b) son ciertas<br />
j) Todas son ciertas
Página192<br />
Entre las venas de mediano calibre, algunas muestran un elevado espesor de capa media. ¿Cuál de las siguientes<br />
venas no corresponde a este grupo?<br />
a) Las venas de un útero grávido<br />
b) Las venas meníngeas<br />
c) La vena femoral<br />
d) La vena poplítea<br />
e) La vena umbilical<br />
Con respecto a la pars tuberalis de la hipófisis:<br />
a) No presenta células endocrinas<br />
b) La vascularización es abundante<br />
c) Recibe inervación adrenérgica del diencéfalo<br />
d) Controla los cambios de coloración por la producción de MSH<br />
e) Todas son falsas<br />
El <strong>com</strong>ponente mayoritario del lóbulo posterior de la hipófisis:<br />
f) Son los pituicitos<br />
g) Son axones del tracto diencéfalo-hipofisario<br />
h) Son los cuerpos neuronales secretores de hormonas<br />
i) Son las células de Schwann<br />
j) Son los oligodendrocitos<br />
Las células parafoliculares del tiroides:<br />
f) Se localizan en el espacio de Disse<br />
g) Contactan directamente con la luz del folículo<br />
h) Forman una capa continua<br />
i) También se llaman células claras aunque tienen gránulos densos en su citoplasma basal<br />
j) Todas son verdaderas<br />
Con respecto a los folículos tiroideos:<br />
f) Presentan un epitelio alto cuando la glándula está activa<br />
g) Cuando el epitelio está activado presenta menos microvellosidades<br />
h) Su epitelio es siempre cúbico<br />
i) Cuando la glándula está en reposo las células son cilíndricas altas<br />
j) a) y b) son ciertas<br />
Dentro de las glándulas paratiroideas:<br />
f) las células principales claras tienen muchas mitocondrias<br />
g) Las células principales oscuras son PASh)<br />
Las células oxífilas son células en plena actividad secretora<br />
i) Las células principales claras y oscuras son dos tipos celulares independientes<br />
j) Todas son verdaderas<br />
¿Cual de las siguientes afirmaciones acerca de la glándula suprarrenal es falsa?:<br />
f) La corteza es de origen mesodérmico<br />
g) Vasos y fibras nerviosas entran a través de tabiques conjuntivos<br />
h) En el estroma hay ausencia de fibras reticulares<br />
i) La médula tiene un doble aporte sanguíneo arterial y venoso<br />
j) Sólo la secreción de la médula tiene control nervioso
Página193<br />
En la médula de la glándula suprarrenal:<br />
f) Las células secretoras de adrenalina son las menos frecuentes<br />
g) Las células que secretan noradrenalina tienen gránulos con material irregular<br />
h) Sus células son autofluorescentes<br />
i) Todas son verdaderas<br />
j) b) y c) son verdaderas<br />
En la glándula suprarrenal, la hormona hipofisaria ACTH es necesaria para que se mantenga:<br />
k) La zona fascicular<br />
l) La zona reticular<br />
m) La zona glomerulosa<br />
n) a) y b) son ciertas<br />
o) Todas son ciertas<br />
¿Qué dos orgánulos son imprescindibles para la síntesis de ésteres en la corteza suprarrenal?:<br />
a) RER y REL<br />
b) REL y A. Golgi<br />
c) REL y mitocondrias<br />
d) RER y A. Golgi<br />
e) RER y mitocondrias<br />
Señala cuál de estas afirmaciones no es correcta:<br />
f) Existe una barrera que separa las células principales de los paraganglios de la sangre<br />
g) En la glándula pineal existen astrocitos<br />
h) Las células epitelioides de la médula se originan a partir de la corteza fetal que acaba desapareciendo<br />
i) Las células de las glándulas paratiroideas tienen gránulos densos a los electrones<br />
j) En la glándula pineal del adulto puede aparecer arena cerebral<br />
Con respecto a los pinealocitos:<br />
f) Sus prolongaciones se extienden sobre los vasos de los tabiques<br />
g) Además de hormonas peptídicas contienen proteinas transportadoras o neurofisinas<br />
h) Se localizan preferentemente hacia la periferia de la glándula<br />
i) Todas son verdaderas<br />
j) a) y b) son verdaderas