The orchid flora of Cocos Island National Park ... - Epidendra
The orchid flora of Cocos Island National Park ... - Epidendra
The orchid flora of Cocos Island National Park ... - Epidendra

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
Botanical Journal <strong>of</strong> the Linnean Society, 2011, 166, 20–39. With 7 figures<br />
<strong>The</strong> <strong>orchid</strong> <strong>flora</strong> <strong>of</strong> <strong>Cocos</strong> <strong>Island</strong> <strong>National</strong> <strong>Park</strong>,<br />
Puntarenas, Costa Rica<br />
DIEGO BOGARÍN 1 *, JORGE WARNER 1 , MARTYN POWELL 2 and<br />
VINCENT SAVOLAINEN 2,3<br />
1 Jardín Botánico Lankester, Universidad de Costa Rica. P.O. Box 302-7050 Cartago, Costa Rica, A.C.<br />
2 Imperial College London, Silwood <strong>Park</strong>, Ascot, Berkshire, SL5 7PY, UK<br />
3 Royal Botanic Gardens, Kew, Richmond, Middlesex, TW9 3DS, UK<br />
Received 13 January 2011; revised 31 January 2011; accepted for publication 2 February 2011<br />
<strong>The</strong> <strong>orchid</strong> <strong>flora</strong> <strong>of</strong> <strong>Cocos</strong> <strong>Island</strong> <strong>National</strong> <strong>Park</strong>, Costa Rica is described based on field collections and herbarium<br />
sampling. <strong>Cocos</strong> <strong>Island</strong> is an oceanic island situated 550 km south-west <strong>of</strong> Puntarenas (the main Costa Rican port<br />
on the Pacific coast) and 680 km north-east <strong>of</strong> the Galápagos <strong>Island</strong>s. Five species <strong>of</strong> <strong>orchid</strong>s were recorded in this<br />
small area <strong>of</strong> 24 km 2 : Camaridium micranthum and Ornithidium adendrobium (both formerly included in<br />
Maxillaria) and three endemic species <strong>of</strong> Epidendrum (E. cocoense, E. insulanum and E. jimenezii). <strong>The</strong> species are<br />
described and analytical illustrations are provided for each. A key to the species for field identification based on<br />
morphology is presented. Biogeography, ecology, taxonomy, evolution and conservation status <strong>of</strong> the species are also<br />
discussed. © 2011 <strong>The</strong> Linnean Society <strong>of</strong> London, Botanical Journal <strong>of</strong> the Linnean Society, 2011, 166, 20–39.<br />
ADDITIONAL KEYWORDS: Camaridium – Epidendrum – floristics – Maxillaria – oceanic islands –<br />
Orchidaceae – Ornithidium – protected areas – taxonomy.<br />
INTRODUCTION<br />
Isla del Coco, also known as <strong>Cocos</strong> <strong>Island</strong>, is an<br />
oceanic island in the south-eastern tropical Pacific<br />
Ocean (5°31′39″N, 87°03′32″W), situated 550 km<br />
south-west <strong>of</strong> Puntarenas, the main port <strong>of</strong> Costa Rica<br />
on the Pacific coast (Fig. 1). <strong>The</strong> nearest continental<br />
point is Cabo Blanco on the Península de Nicoya,<br />
Costa Rica, 532 km from the island. Other oceanic<br />
islands relatively near <strong>Cocos</strong> <strong>Island</strong> are Isla Pinta <strong>of</strong><br />
the Galápagos archipelago (Ecuador) at 681 km, Isla<br />
Malpelo (Colombia) at 630 km, Isla Coiba (Panama)<br />
at 632 km and Clipperton <strong>Island</strong> (Île de Clipperton or<br />
Île de la Passion, France) at 2375 km.<br />
<strong>Cocos</strong> <strong>Island</strong> became part <strong>of</strong> Costa Rica in 1869,<br />
when President Jesús Jiménez decreed it as national<br />
territory. At that time, the island was uninhabited and<br />
had never been claimed by any other country. Its name<br />
was probably a mistranscription from the original map<br />
*Corresponding author. E-mail: diego.bogarin@ucr.ac.cr<br />
20<br />
by the French cartographer Nicolas Desliens in 1541,<br />
in which the island appeared (for first time) as ‘Y e<br />
Coques’ (Hogue & Miller, 1981; Trusty, 2004). In<br />
French, the word ‘coque’ means eggshell or nutshell<br />
and the name could have been a reference to the<br />
common fruit <strong>of</strong> the abundant endemic tree Sacoglottis<br />
holdridgei Cuatrec. (Humiriaceae).<br />
In 1978, under the government <strong>of</strong> President<br />
Rodrigo Carazo, the island was decreed a <strong>National</strong><br />
<strong>Park</strong> and a zone <strong>of</strong> absolute protection. Administratively,<br />
the area belongs to the Area de Conservación<br />
Marina Isla del Coco (ACMIC) <strong>of</strong> the Costa Rican<br />
Ministry <strong>of</strong> Environment, Energy and Telecommunications<br />
(MINAET) and its <strong>National</strong> System <strong>of</strong> Conservation<br />
Areas (SINAC). Later, in 1997 the island<br />
became a UNESCO-designated World Natural Heritage<br />
Site in recognition <strong>of</strong> its exceptional biodiversity.<br />
In 2002, the World Natural Heritage Site designation<br />
was extended to include an expanded marine zone<br />
<strong>of</strong> 1997 km 2 , thus protecting marine ecosystems,<br />
populations <strong>of</strong> sharks, rays, dolphins and other large<br />
marine species. In addition, in 1998 the island was<br />
© 2011 <strong>The</strong> Linnean Society <strong>of</strong> London, Botanical Journal <strong>of</strong> the Linnean Society, 2011, 166, 20–39
© 2011 <strong>The</strong> Linnean Society <strong>of</strong> London, Botanical Journal <strong>of</strong> the Linnean Society, 2011, 166, 20–39<br />
ORCHID FLORA OF COCOS ISLAND 21<br />
Figure 1. Map <strong>of</strong> <strong>Cocos</strong> <strong>Island</strong> showing collecting sites. Scale 1 : 25 000.
22 D. BOGARÍN ET AL.<br />
included in the list <strong>of</strong> Wetlands <strong>of</strong> International<br />
Importance under the RAMSAR Convention.<br />
TOPOGRAPHY<br />
<strong>The</strong> total land area is 24.85 km2 (c. 7.6 ¥ 4.4 km) with<br />
the following limits: to the north Punta Agujas, to the<br />
south Cabo Dampier, to the west Cabo Lionel and to<br />
the east Cabo Atrevido (Fig. 1). <strong>The</strong> topography is<br />
abrupt with high slopes and cliffs. <strong>The</strong> highest point<br />
is Cerro Iglesias, observable as a cloudy mountain<br />
rising 575 m above sea level. Other important hills<br />
are Cerro Pelón at 530 m and Cerro Jesús Jiménez at<br />
430 m <strong>of</strong> elevation. Some flat areas are observed in<br />
the region called Llanos Palo de Hierro at 300 m<br />
elevation (Fig. 1). <strong>The</strong> island has four bays: Iglesias,<br />
Weston, Chatham and Wafer. <strong>The</strong> later two are the<br />
most important for boats and crafts and have the<br />
main ranger stations. <strong>The</strong> most important rivers are:<br />
the Genio to the north-east that flows into Wafer Bay,<br />
the Chatham River that flows into Chatham Bay and<br />
the Iglesias to the south, which terminates in Iglesias<br />
Bay. <strong>The</strong> irregular topography produces many waterfalls<br />
because <strong>of</strong> the vertical cliffs and high water<br />
availability in the soil.<br />
GEOLOGY<br />
<strong>Cocos</strong> <strong>Island</strong> emerged from the ocean through volcanic<br />
activity <strong>of</strong> the submarine mountain range called<br />
the <strong>Cocos</strong> Ridge. <strong>The</strong> ridge was formed in the Miocene<br />
by tectonic movements <strong>of</strong> the <strong>Cocos</strong> and Nazca plates<br />
and volcanic action <strong>of</strong> the Galápagos hotspot. <strong>The</strong>se<br />
geological events occurred along the Pacific Ocean<br />
near South America. <strong>The</strong>n, the island moved to the<br />
north as a result <strong>of</strong> the action <strong>of</strong> the <strong>Cocos</strong> and Nazca<br />
plates. Geologists have estimated the age <strong>of</strong> the<br />
island to be 1.9–2.4 million years (Bellon, Sáenz &<br />
Tournon, 1984). <strong>The</strong> soils are composed <strong>of</strong> basaltic<br />
and pyroclastic alkaline rocks, the remains <strong>of</strong> ancient<br />
volcanic activity. However, the acidic condition is<br />
caused by the degradation <strong>of</strong> organic material under<br />
high humidity and the consequent action <strong>of</strong> bacteria<br />
and fungi (Trusty, 2004).<br />
CLIMATE<br />
<strong>The</strong> island is under the influence <strong>of</strong> the Intertropical<br />
Convergence Zone (ITCZ) and receives other climatic<br />
influences such as the El Niño-Southern Oscillation<br />
(ENSO). <strong>The</strong> climate <strong>of</strong> the island was discussed by<br />
Alfaro (2008) and Lizano (2008). A complex system <strong>of</strong><br />
marine currents originating in America and the Indo-<br />
Pacific region flows around the island, thus influencing<br />
its climate (Lizano, 2008). <strong>The</strong> island experiences<br />
high rainfall, which can reach 7000 mm per year.<br />
<strong>Cocos</strong> is the only island in the tropical eastern Pacific<br />
with a tropical humid forest. According to Tosi (1969),<br />
the life zone is the premontane belt forest, basal belt<br />
transition. <strong>The</strong> period <strong>of</strong> high rainfall is between May<br />
and October. <strong>The</strong>re is no true dry season but rainfall<br />
is slightly lower from November to March (Alfaro,<br />
2008). <strong>The</strong> average temperature is approximately<br />
27–28 °C. <strong>The</strong>se climatic factors allow the establishment<br />
<strong>of</strong> plants such as bromeliads, ferns and epiphytic<br />
<strong>orchid</strong>s.<br />
BIODIVERSITY<br />
<strong>The</strong> biodiversity <strong>of</strong> <strong>Cocos</strong> <strong>Island</strong> includes both marine<br />
and terrestrial ecosystems. Marine biodiversity is the<br />
main attraction for touristic and scientific activities.<br />
This place is an important breeding site for many<br />
species. <strong>The</strong> migration and reproduction <strong>of</strong> sharks is<br />
one <strong>of</strong> the most spectacular marine attractions for<br />
divers. It is possible to observe the rare and endangered<br />
whale shark (Rhincodon typus) and large populations<br />
<strong>of</strong> hammerhead sharks (Sphyrna lewinni), the<br />
marine emblem <strong>of</strong> the island. <strong>The</strong> history <strong>of</strong> marine<br />
research at <strong>Cocos</strong> <strong>Island</strong> was reviewed recently by<br />
Cortés (2008). Terrestrial biodiversity is also important.<br />
According to the most recent botanical treatment<br />
<strong>of</strong> the island by Trusty, Kesler & Haug Delgado<br />
(2006), there are 262 species <strong>of</strong> plants, <strong>of</strong> which 37<br />
(approximately 14%) are endemic. Among the terrestrial<br />
fauna, there are 382 species <strong>of</strong> insects, five<br />
species <strong>of</strong> reptiles, one scorpion and 12 species <strong>of</strong><br />
birds, including the endemic Coco finch, Pinaroloxias<br />
inornata. <strong>The</strong>re is a low elevation rainforest formed<br />
mainly by the endemic tree Sacoglottis holdridgei. At<br />
the highest points (i.e. Cerro Iglesias), there is a high<br />
elevation cloud forest. Other important plant communities<br />
are the riparian habitats, estuarine habitats,<br />
coastal cliff communities, shore vegetation, landslides<br />
and rocky islets that have sparse grassy vegetation<br />
and scattered trees <strong>of</strong> Clusia rosea Jacq. <strong>The</strong>se plant<br />
associations were defined and characterized by Trusty<br />
(2004). Detailed information on climate, geology,<br />
human impacts and introduced animals and plants<br />
can be found in Trusty (2004) and Trusty et al. (2006).<br />
THE HISTORY OF COCOS ISLAND ORCHIDS<br />
<strong>The</strong> first recorded visit to <strong>Cocos</strong> <strong>Island</strong> was in 1526,<br />
when the Spanish explorer Joan Cabeças reached the<br />
island. Since then, mariners and pirates have known<br />
the island and it became popular because <strong>of</strong> traditional<br />
stories <strong>of</strong> treasure supposedly buried there.<br />
According to the legend, two treasures have been<br />
hidden on <strong>Cocos</strong> <strong>Island</strong>: firstly, the pirate Bennett<br />
Graham (Benito Bonito) <strong>of</strong> the Relámpago, who looted<br />
several cities around South America, is reputed to<br />
© 2011 <strong>The</strong> Linnean Society <strong>of</strong> London, Botanical Journal <strong>of</strong> the Linnean Society, 2011, 166, 20–39
have kept his treasure in an unknown place on the<br />
island (Hogue & Miller, 1981; Trusty, 2004) and, secondly,<br />
the ‘treasure <strong>of</strong> Lima’ <strong>of</strong> Captain Thompson <strong>of</strong><br />
the Mary Dear may be on <strong>Cocos</strong>, including priceless<br />
gold and jewels from Peru. Following these tales,<br />
people looking for treasures made > 400 expeditions<br />
to the island and <strong>Cocos</strong> is therefore also well known<br />
as the ‘treasure island’.<br />
In spite <strong>of</strong> these hundreds <strong>of</strong> expeditions, plant<br />
collections only started in 1836 when the English<br />
collector George W. Barclay visited <strong>Cocos</strong> <strong>Island</strong> as<br />
part <strong>of</strong> an exploration <strong>of</strong> the Pacific coast aboard<br />
H.M.S. Sulphur, commanded by Captain Sir Edward<br />
Belcher (Bentham, 1844–1846). However, in their<br />
botanical treatment they did not report or describe<br />
any <strong>orchid</strong> species. After Barclay’s expedition, the<br />
marine zoologist and oceanographer Alexander<br />
Agassiz collected plants during the late 19 th century<br />
(1888 and 1891). Aboard the Albatross, he visited<br />
<strong>Cocos</strong> <strong>Island</strong> on his way to the Galápagos. He collected<br />
Epidendrum insulanum Schltr. (then undescribed),<br />
the first <strong>orchid</strong> specimen collected in the<br />
island (Agassiz s.n., GH).<br />
In 1898, the government <strong>of</strong> President Rafael Yglesias<br />
sponsored the first Costa Rican expedition. He<br />
assigned to the scientists Anastasio Alfaro and Henry<br />
Pittier the task <strong>of</strong> assessing the island in order for<br />
it to be reopened as a penal center (Pittier, 1898).<br />
<strong>The</strong>y went aboard the Poás and collected plants<br />
and animals. Four years later, on a second trip in<br />
1902, together with Paul Biolley, aboard the Turrialba,<br />
Pittier collected two specimens <strong>of</strong> Epidendrum<br />
L. One was the first <strong>orchid</strong> species described from the<br />
island, published by R. Schlechter as E. insulanum<br />
in Beihefte zum Botanischen Centralblatt in 1918.<br />
Unfortunately, the specimen studied by Schlechter<br />
was destroyed in the Dahlem-Berlin Herbarium<br />
during the Second World War and there is no indication<br />
that Schlechter received more than one specimen<br />
from Pittier. However, Pittier prepared at least five<br />
duplicates; two are kept at the herbarium <strong>of</strong> the<br />
Museo Nacional de Costa Rica (CR) and three at<br />
Oakes Ames Orchid Herbarium <strong>of</strong> Harvard University<br />
(AMES), GH-3581 (=AMES-70445), GH-3579<br />
(=AMES-70446) and the lectotype designated by<br />
Trusty & Blanco (2005) GH-3580 (=AMES-73449).<br />
<strong>The</strong>se specimens are available under Pittier n. 16350<br />
(Institute. Phys. Geogr. Cost.); the same number is<br />
cited by Schlechter (1918) in the protologue. A sheet<br />
with tracings <strong>of</strong> Schlechter’s drawing <strong>of</strong> the holotype<br />
is also kept at AMES (AMES-70447). <strong>The</strong> second<br />
species recorded by Pittier correspond to at least two<br />
plants <strong>of</strong> E. cocoense Hágsater (then undescribed)<br />
kept at AMES under Pittier n. 16351 (Institute. Phys.<br />
Geogr. Cost.). Hágsater (1999a) identified the specimens<br />
as E. cocoense. As Schlechter did not mention<br />
ORCHID FLORA OF COCOS ISLAND 23<br />
any <strong>of</strong> these specimens or describe another species, it<br />
is probable that Pittier did not send any more specimens<br />
to Berlin, but no information is available to<br />
clarify this situation.<br />
Robert E. Snodgrass and Edmund Heller in the<br />
Hopkins-Stanford Expedition made another expedition<br />
in 1899. Alban Stewart collected again for the<br />
California Academy <strong>of</strong> Sciences in 1905. <strong>The</strong>n, Henry<br />
Knute Svenson, who was the curator <strong>of</strong> Brooklyn<br />
Botanical Garden, New York, spent 3 days collecting<br />
plants in the island as the chief botanist on the<br />
Vincent Astor Expedition in April 1930 (Svenson,<br />
1935). Svenson found what is now the type specimen<br />
<strong>of</strong> E. cocoense growing by a stream flowing into Wafer<br />
Bay (H. K. Svenson 333, AMES-37853); the species<br />
was later described by Hágsater (1999a). Svenson<br />
prepared two duplicates; one is at AMES and the<br />
other at Chicago (F).<br />
Svenson was the first to collect specimens <strong>of</strong> Camaridium<br />
Lindl. (the specimens from the island were<br />
formerly included in Maxillaria Ruiz & Pav.). <strong>The</strong>y<br />
were misidentified under the names M. concavilabia<br />
Ames & Correll and Ornithidium aff. stenophyllum<br />
Schltr. In 1994, John T. Atwood identified the<br />
specimens as M. parvi<strong>flora</strong> (Poepp. & Endl.)<br />
Garay (=Camaridium micranthum M.A.Blanco, see<br />
Discussion).<br />
Two years after Svenson’s trip, the American botanist<br />
John Thomas Howell explored the island on <strong>The</strong><br />
Templeton Crocker Expedition sponsored by the California<br />
Academy <strong>of</strong> Sciences. He collected another<br />
specimen <strong>of</strong> E. cocoense erroneously identified as E.<br />
ramosum Jacq. var. imbricatum Ames, F.T.Hubb. &<br />
C.Schweinf. Later, President Cruise <strong>of</strong> the USA made<br />
other expeditions from 1935 to 1940.<br />
<strong>The</strong> Costa Rican botanist and ex-director <strong>of</strong> the<br />
Museo Nacional de Costa Rica, Juvenal Valerio<br />
Rodríguez, went to the island on two expeditions<br />
during 1939 and 1940. He collected C. micranthum<br />
(misidentified as Maxillaria variabilis Bateman ex<br />
Lindl) and another specimen <strong>of</strong> E. cocoense was collected<br />
in his second trip.<br />
Witold L. Klawe, a fishery researcher, collected<br />
plants as part <strong>of</strong> the Inter-American Tropical Tuna<br />
Commission aboard the Costa Rica Dome Cruise in<br />
1959. He collected E. cocoense, wrongly identified<br />
under the names E. paranaense Barb. Rodr. and E.<br />
santaclarense Ames.<br />
Hágsater (1999b) described the last species known<br />
from the island under the name E. jimenezii. <strong>The</strong><br />
species honours Alfonso Jiménez, ex-director <strong>of</strong> the<br />
Museo Nacional de Costa Rica, who collected the type<br />
specimen in 1965 (A. Jiménez 3178, CR). He stated<br />
that the plant had white flowers and it is apparently<br />
a common epiphyte along the Genio River, near its<br />
mouth into Wafer Bay.<br />
© 2011 <strong>The</strong> Linnean Society <strong>of</strong> London, Botanical Journal <strong>of</strong> the Linnean Society, 2011, 166, 20–39
24 D. BOGARÍN ET AL.<br />
In 1967, the botanists Ira L. Wiggins and Duncan<br />
M. Porter, who worked on the Flora <strong>of</strong> the Galápagos<br />
<strong>Island</strong>s, collected specimens <strong>of</strong> E. insulanum (Hágsater,<br />
2007). Luis D. Gómez went to the island in 1970<br />
and made another collection <strong>of</strong> E. cocoense from the<br />
forest <strong>of</strong> Wafer Bay.<br />
After many expeditions, the number <strong>of</strong> species <strong>of</strong><br />
the island remained unclear. In 1973, in a trip organized<br />
by the Smithsonian Tropical Research Institute,<br />
Robert L. Dressler collected an Epidendrum, identified<br />
by Hágsater as E. jimenezii (Dressler 4468, PMA-<br />
36761), but a duplicate kept at CR (Dressler 4468,<br />
CR) could be an E. insulanum as noted by Trusty &<br />
Blanco (2006). A mixed collection was probably made<br />
under the same number, demonstrating the difficulties<br />
in identifying those species without flowers as<br />
their vegetative features are highly variable. Another<br />
sample <strong>of</strong> E. cocoense was collected and a new record,<br />
Ornithidium adendrobium (Rchb.f.) M.A.Blanco &<br />
Ojeda (formerly known as Maxillaria adendrobium<br />
(Rchb.f.) Dressler) was found near Wafer Bay. This<br />
widespread species ranges in both continental and<br />
insular lands around the Neotropics. In 1865, Heinrich<br />
G. Reichenbach described it as Ponera adendrobium<br />
Rchb.f. based on a collection <strong>of</strong> Charles Wright<br />
in Cuba.<br />
In 1979, Robin Foster collected an interesting Epidendrum<br />
in the floodplain <strong>of</strong> Genio River. It seems to<br />
be an example <strong>of</strong> E. insulanum, but Hágsater cited<br />
it as E. jimenezii (Hágsater, 1999b). In 1980, Pablo<br />
Sánchez-Vindas collected E. cocoense and in 1981<br />
Jorge Gómez-Laurito collected E. jimenezii (wrongly<br />
identified as Epidendrum imbricatum Lam.) and<br />
another E. insulanum. He also collected E. cocoense,<br />
and Luis D. Gómez collected M. parvi<strong>flora</strong> in the<br />
same year. In 1989, R. Soto collected E. cocoense, O.<br />
adendrobium and other two specimens <strong>of</strong> C. micranthum.<br />
Other collections were made in 1994 by the<br />
researchers <strong>of</strong> Instituto Nacional de Biodiversidad<br />
(INBio). However, they reported the same five<br />
species. With the description <strong>of</strong> E. cocoense and<br />
E. jimenezii by Hágsater (1999b), the <strong>orchid</strong> <strong>flora</strong> <strong>of</strong><br />
<strong>Cocos</strong> <strong>Island</strong> is made up <strong>of</strong> three endemic species <strong>of</strong><br />
Epidendrum (E. cocoense, E. insulanum and E. jimenezii),<br />
one species <strong>of</strong> Camaridium, C. micranthum,<br />
and one species <strong>of</strong> Ornithidium, O. adendrobium<br />
(Dressler, 2003). <strong>The</strong> two former species are also<br />
found in continental parts <strong>of</strong> the Neotropics.<br />
Recently, Trusty and co-workers (Trusty, 2004;<br />
Trusty & Blanco, 2005, 2006; Trusty et al., 2006)<br />
finished their floristic treatment <strong>of</strong> the <strong>Cocos</strong> <strong>Island</strong><br />
<strong>National</strong> <strong>Park</strong>. <strong>The</strong>y reported the same five species<br />
for the island. However, they were unable to locate<br />
material <strong>of</strong> E. jimenezii, and consequently they could<br />
not distinguish this species from E. insulanum<br />
(Trusty & Blanco, 2006).<br />
<strong>The</strong> aim <strong>of</strong> this paper is to present detailed information<br />
for identifying the <strong>orchid</strong>s <strong>of</strong> <strong>Cocos</strong> <strong>Island</strong>, to<br />
provide illustrations <strong>of</strong> the five species based on living<br />
plants (two <strong>of</strong> them are illustrated for a first time)<br />
and to discuss the identity <strong>of</strong> the rare E. jimenezii.<br />
This work is part <strong>of</strong> a series <strong>of</strong> floristic studies<br />
intended to clarify the diversity and taxonomy <strong>of</strong><br />
Orchidaceae in Costa Rica (e.g. Pupulin, 2010) and its<br />
national parks and protected areas (Pupulin, 1998;<br />
Bogarín & Pupulin, 2007).<br />
MATERIAL AND METHODS<br />
This work was conducted mainly in the <strong>Cocos</strong> <strong>Island</strong><br />
<strong>National</strong> <strong>Park</strong>, Puntarenas, Costa Rica and the<br />
Lankester Botanical Garden (LBG), University <strong>of</strong><br />
Costa Rica. Living specimens were collected, cultivated<br />
and documented between 2006 and 2009. Data<br />
from all specimens cited have been recorded in a<br />
computerized database at LBG. <strong>The</strong>y are also available<br />
on the website at http://www.epidendra.org<br />
(Pupulin, 2007; Pupulin, 2009). Georeferences for<br />
specimens were obtained using a Garmin eTrex Vista<br />
GPS and maps. Ecological zones were estimated by<br />
using the Holdridge Life Zone System (Holdridge,<br />
1967, 1987) and the ecological map <strong>of</strong> Costa Rica<br />
(Tosi, 1969).<br />
Phenology data were recorded both in the field and<br />
in cultivated specimens or herbarium labels. Individual<br />
plants were photographed, illustrated and<br />
preserved as herbarium specimens and preserved<br />
specimens in formaldehyde : acetic acid : ethanol<br />
[FAA (53% ethanol, 37% water, 5% formaldehyde and<br />
5% glycerol)] (including flowers, portions <strong>of</strong> the stems<br />
or entire plants) for future reference. Newly collected<br />
herbarium specimens were deposited at CR, JBL and<br />
USJ herbaria. Whenever possible, the herbarium<br />
specimens were complemented with sketches, photographs<br />
and FAA material. <strong>The</strong> material preserved in<br />
FAA was deposited at JBL and indicated in the treatment<br />
as ‘JBL-spirit’. Herbarium and spirit material<br />
may consist <strong>of</strong> wild collected specimens or material<br />
collected entirely from cultivated plants.<br />
Sketches <strong>of</strong> specimens were made with a Leica<br />
MZ7.5 stereomicroscope with drawing tube and conserved<br />
in the reference collections at JBL. All taxa<br />
were illustrated by composite line drawings from<br />
living specimens. Illustrations include a typical plant<br />
habit, inflorescences or part <strong>of</strong> the inflorescences, the<br />
flower and a dissection <strong>of</strong> perianth. Plate composition<br />
was as consistent as possible to facilitate species<br />
comparison.<br />
Descriptions were prepared from both living specimens<br />
and herbarium material. Materials from the<br />
following herbaria were studied: AMES, BM, CAS,<br />
CR, INB, JBL, K, PMA and USJ.<br />
© 2011 <strong>The</strong> Linnean Society <strong>of</strong> London, Botanical Journal <strong>of</strong> the Linnean Society, 2011, 166, 20–39
KEY TO THE ORCHIDS OF THE COCOS ISLAND NATIONAL PARK<br />
LIST OF SPECIES<br />
1. CAMARIDIUM LINDL.<br />
CAMARIDIUM LINDL., Bot. Reg. 10: sub t. 844. 1824.<br />
Type species: Camaridium ochroleucum Lindl., Bot.<br />
Reg. 10: 844. 1824.<br />
PLANT epiphytic, caespitose or straggling plants,<br />
small to large in size, variable in habit. PSEUDOBULBS<br />
unifoliate, separated by an elongate rhizome, caespitose<br />
or lacking pseudobulbs with erect, monopodial<br />
leafy shoots. Rarely with dimorphic growth. LEAVES<br />
conduplicate, thin or coriaceous to subcoriaceous, conduplicate.<br />
INFLORESCENSES produced from a leafy<br />
new growth. From several to many. FLORAL BRACT<br />
longer than the ovary. FLOWERS small and inconspicuous<br />
to medium sized and showy. SEPALS AND<br />
PETALS subsimilar, spreading. LIP <strong>of</strong>ten shorter than<br />
the other perianth parts and <strong>flora</strong>l segments, lacking<br />
fibres, simple or three-lobed, articulated with or<br />
adnate to the column, usually provided at the base<br />
with a fleshy callus. COLUMN erect, terete. CAPSULES<br />
have apical dehiscence.<br />
Camaridium is a Neotropical genus with wide distribution<br />
and approximately 80 species. <strong>The</strong> highest<br />
diversity is found in Central America.<br />
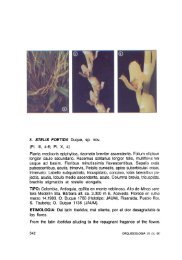
1. CAMARIDIUM MICRANTHUM M.A.BLANCO, Lankesteriana<br />
7: 520. 2007. Scaphyglottis parvi<strong>flora</strong> Poepp.<br />
& Endl., nov. gen. ac Sp. 1: 58, t. 97. 1835. Maxillaria<br />
parvi<strong>flora</strong> (Poepp. & Endl.) Garay, Bot. Mus. Leafl.<br />
21: 258. 1967. Non Camaridium parviflorum Fawc.,<br />
Symb. Antill. 1: 472. 1910. Type: PERU. Crescit ad<br />
arbores Peruviae in viciniis praedii Cuchero, floret<br />
Feb, Poeppig s.n. (holotype: W). (Figs 2, 3A, B).<br />
Description: PLANT epiphytic, pendent or suberect, to<br />
25 cm long. ROOTS flexuous, < 1.5 mm in diameter,<br />
white to greenish, with green or orange–yellowish<br />
tips. PSEUDOBULBS unifoliate, ovoid, pyriform, separated<br />
by an elongate rhizome, 2.5–6 ¥ 0.8–1.3 cm,<br />
internodes up to 3 cm long, covered by scarious,<br />
imbricate papyraceous sheaths. LEAVES apical on<br />
ORCHID FLORA OF COCOS ISLAND 25<br />
1. Plants with pseudobulbs, each with one apical leaf.................................................Camaridium micranthum<br />
1. Plant without pseudobulbs, the leaves distichously arranged.....................................................................2<br />
2. Inflorescense lateral, lip adnate to the column foot................................................Ornithidium adendrobium<br />
2. Inflorescence apical, lip adnate to the footless column..............................................................................3<br />
3. Inflorescence with one green–yellow flower < 1 cm in length, apex <strong>of</strong> column erose, lateral lobes <strong>of</strong> lip not covering<br />
the column apex .................................................................................................Epidendrum insulanum<br />
3. Inflorescence with two or more white to whitish flowers > 1 cm in length, column apex with two teeth in each side,<br />
lateral lobes <strong>of</strong> lip covering the column apex..........................................................................................4<br />
4. Leaves 1.5–2.7 cm wide, <strong>flora</strong>l bracts 1.3–1.6 cm wide, broadly ovate, larger than the scurfy ovary...Epidendrum<br />
cocoense<br />
4. Leaves 1.0–1.2 cm wide, <strong>flora</strong>l bracts 0.5–0.6 cm wide, tubular, as long as the glabrescent ovary......Epidendrum<br />
jimenezii<br />
each pseudobulb, petiolate, oblong–linear, acute,<br />
emarginate, unequally two-lobed at the apex, conduplicate,<br />
subcoriaceous, to approximately 10–20 ¥ 1.0–<br />
1.4 cm. INFLORESCENCE a single-flowered raceme,<br />
produced in clusters along the stem, at the base <strong>of</strong><br />
each pseudobulb, covered by several scarious, imbricate,<br />
acute, papyraceous sheaths. PEDICEL inconspicuous.<br />
FLORAL BRACTS triangular, ovate, scarious.<br />
OVARY cylindrical, < 4 mm long. FLOWERS small,<br />
inconspicuous, white with a yellow dot in the lip<br />
callus, approximately 5 mm in length. DORSAL SEPAL<br />
oblong, acute, conduplicate, concave towards the<br />
apex, 5 ¥ 2 mm. LATERAL SEPALS narrowly ovate,<br />
acute, slightly conduplicate, concave, 4.5 ¥ 2.5 mm.<br />
PETALS narrowly oblong, acute, 3.8 ¥ 0.8 mm. LIP<br />
simple, spathulate, oblong, obtuse, attached to the<br />
column foot, conduplicate, callus laminate, 3.0 ¥<br />
1.5 mm. COLUMN short, terete, to 1 mm long, anther<br />
apical, stigma ventral, rectangular. POLLINIA four,<br />
ovoid, with a short stipe, viscidium elliptic. ANTHER<br />
CAP subquadrate–cucullate.<br />
Other vouchers examined: COSTA RICA. Puntarenas:<br />
Puntarenas, Isla del Coco, cumbre del Cerro Iglesias,<br />
575 m, 5°31′45.8N, 87°04′80.5″W, bosque pluvial premontano,<br />
transición a basal, epífitas en Sacoglottis<br />
holdridgei, 15 Abril 2006, D. Bogarín 2766, G. Blanco,<br />
G. Gigot, V. Savolainen & J. Warner (JBL-Spirit);<br />
Puntarenas: Puntarenas, Isla del Coco, camino entre<br />
Bahía Wafer y Bahía Chatham, orillas del sendero,<br />
210 m, 5°32′84.07″, N 87°03′0.18″W, bosque pluvial<br />
premontano, transición a basal, epífitas en Sacoglottis<br />
holdridgei, 14 abril 2006, D. Bogarín 2766, G. Blanco,<br />
G.Gigot, V. Savolainen & J. Warner (JBL-Spirit);<br />
Puntarenas: Puntarenas, Isla del Coco, camino al<br />
Cerro Iglesias, ascenso en el primer mirador, 275 m,<br />
5°32′14.09N, 87°03′36.8″W, bosque pluvial premontano,<br />
transición a basal, epífitas en Sacoglottis holdridgei,<br />
15 abril 2006, D. Bogarín 2746, G. Blanco,<br />
G.Gigot, V. Savolainen & J. Warner (JBL-Spirit). Pun-<br />
© 2011 <strong>The</strong> Linnean Society <strong>of</strong> London, Botanical Journal <strong>of</strong> the Linnean Society, 2011, 166, 20–39
26 D. BOGARÍN ET AL.<br />
Figure 2. Camaridium micranthum M.A.Blanco. A, habit. B, flower. C, dissected perianth. D, column and lip, lateral<br />
view. E, column, lateral and front views. F, pollinarium anther cap. Drawn by D. Bogarín from Bogarín 2746 (JBL-Spirit).<br />
© 2011 <strong>The</strong> Linnean Society <strong>of</strong> London, Botanical Journal <strong>of</strong> the Linnean Society, 2011, 166, 20–39
ORCHID FLORA OF COCOS ISLAND 27<br />
Figure 3. Floral morphology <strong>of</strong>: (A, B) Camaridium micranthum; (C) Epidendrum cocoense; (D) Epidendrum insulanum;<br />
(E) Epidendrum jimenezii; (F) Ornithidium adendrobium.<br />
© 2011 <strong>The</strong> Linnean Society <strong>of</strong> London, Botanical Journal <strong>of</strong> the Linnean Society, 2011, 166, 20–39
28 D. BOGARÍN ET AL.<br />
tarenas: Isla del Coco. Bahía Chatham, 5°32′82″N,<br />
82°02′17″W, c. 200 m, 12 marzo 1994, G. Dauphin<br />
1175 (CR, USJ); J. Gómez 18043 (CR); Quesada 1081<br />
(INB); A. Rojas 3659 (CR, INB); 6–8 junio 1989, R.<br />
Soto s.n. (USJ-49289 and 49290); along a brook<br />
flowing into Wafer Bay, April 18, 1930, H. K. Svenson<br />
334 (GH); <strong>Cocos</strong> <strong>Island</strong>. Closed canopy forest, 5°31′N,<br />
87°03′W, 26 January 2002, J. Trusty 104, 156, 244,<br />
482, and 504 (CR); Isla del Coco, diciembre 1939,<br />
J. Valerio 1086 and 1106 (CR).<br />
Distribution: this species ranges from South Florida,<br />
Antilles and Mexico to Honduras and from Colombia<br />
to Brazil. It seems oddly absent from Nicaragua to<br />
Panama. In Costa Rica it is known only from <strong>Cocos</strong><br />
<strong>Island</strong>.<br />
Etymology: From the Greek micranthum, ‘small<br />
flower’, in allusion to the small and inconspicuous<br />
flowers <strong>of</strong> this species.<br />
Phenology: September to April<br />
Habitat and ecology: a common epiphyte in premontane<br />
rainforest, basal belt transition. It is found from<br />
sea level to the top <strong>of</strong> Cerro Iglesias at 585 m. <strong>The</strong>y<br />
flower throughout the year but mostly between<br />
November to April Pollination is possibly carried out<br />
by ants (Singer, 2003). <strong>The</strong>re are no bees in the island<br />
but there are several species <strong>of</strong> ants. Apparently, the<br />
plants are not self-pollinated. Some plants in cultivation<br />
at Lankester Botanical Garden did not produce<br />
fruits.<br />
Discussion: This is the only species on the island<br />
having pseudobulbs with one apical leaf and the<br />
pseudobulbs are produced in chains. It can be recognized<br />
by the inconspicuous flowers in clusters at the<br />
base <strong>of</strong> the pseudobulbs, the partially connate lateral<br />
sepals, the nearly absent mentum and the cuneate<br />
lip. Poeppig and Endlicher described this species in<br />
Nova Genera ac Species Plantarum under the name<br />
Scaphyglottis parvi<strong>flora</strong> in 1835. <strong>The</strong> name was based<br />
on a plant collected in Cuchero, Peru. Garay transferred<br />
this species to Maxillaria (Garay, 1967).<br />
Molecular studies and nomenclatural changes in<br />
Maxillariinae Benth. & Hook.f. (Blanco et al., 2007;<br />
Whitten et al., 2009) revealed that this species should<br />
be placed under the resurrected Camaridium. Asthe<br />
name Camaridium parviflorum Fawc. was previously<br />
applied to a different species (a synonym <strong>of</strong> O. adendrobium,<br />
a species also occurring in the island), thus<br />
preventing its use in Camaridium, a new name, C.<br />
micranthum, was proposed by Blanco et al. (2007).<br />
Conservation status: This species is not rare and has<br />
a wide distribution range throughout Neotropics.<br />
In the island it is a common epiphyte. This species<br />
should not be regarded as endangered and may well<br />
fit the IUCN Least Concern (LC) category for widespread<br />
and abundant taxa.<br />
2. EPIDENDRUM L.<br />
EPIDENDRUM L., Sp. Pl. ed. 2: 1347. 1763., nom. cons.<br />
Type species: Epidendrum nocturnum Jacq., type.<br />
cons., Enum. Pl. Carib. 29. Jan–December 1760.<br />
PLANT epiphytic or rarely terrestrial herbs to subshrubs,<br />
small to large, variable in size and habit,<br />
caespitose, creeping, erect to pendent, occasionally<br />
with pseudobulbs. LEAVES distichously arranged<br />
along the stem, occasionally one apical leaf or several<br />
apical leaves distributed throughout the stem or<br />
aggregate at the apex <strong>of</strong> the stem or pseudobulb,<br />
conduplicate, subcoriaceous to fleshy, not petiolate.<br />
INFLORESCENCE apical, rarely lateral, a one- to manyflowered<br />
raceme, spike, panicle or umbel. FLOWERS<br />
resupinate or not, small to large and showy. SEPALS<br />
AND PETALS subsimilar, free, usually spreading; the<br />
lip simple or lobed, mostly united to the ventral<br />
portion <strong>of</strong> the column, usually with fleshy calli at the<br />
base. COLUMN mostly totally fused with the lip, but<br />
sometimes totally free or half fused with the lip just<br />
to the apex, <strong>of</strong>ten provided with a hooded clinandrium.<br />
POLLINIA two, four or rarely eight, waxy.<br />
Epidendrum is one <strong>of</strong> the largest genera <strong>of</strong> Neotropical<br />
<strong>orchid</strong>s with more than 1500 species. <strong>The</strong> genus<br />
is widely distributed throughout the Neotropics, from<br />
North Carolina to northern Argentina including the<br />
Antilles.<br />
1. EPIDENDRUM COCOENSE HÁGSATER, Icon. Orchid.<br />
(Mexico) 3: pl. 325. 1999. Type: COSTA RICA, PUN-<br />
TARENAS, <strong>Cocos</strong> <strong>Island</strong>, along brook flowing into<br />
Wafer Bay, on trees, 18 April 1930, H.K. Svenson<br />
333 (holotype: AMES; isotypes: AMES, BKL, F).<br />
(Figs 3C, 4).<br />
Description: PLANT a large epiphyte, pendent, with<br />
terete, leafy stem up to 2.5 m long. Roots thick, up<br />
to 1–2 mm in diameter, white to greenish, with green<br />
tips, produced basally. STEM terete, branching, canelike,<br />
with lateral, secondary stems shorter than the<br />
main stems, each stems ending in apical inflorescences.<br />
LEAF SHEATHS tubular, enclosing the stem,<br />
laterally flattened, to 3 cm long. LEAVES many (to 30),<br />
distichous, distributed throughout the stem, laterally<br />
twisted, lanceolate, elliptic–oblong, acute, with a<br />
bilobed apex, conduplicate, subcoriaceous, articulate<br />
with the sheath involving the stem, to approximately<br />
5–14 ¥ 1.5–2.7 cm. INFLORESCENCE apical, racemose,<br />
distichous, two- to four-flowered, shorter than the<br />
leaves, flowering once, to 6 cm long, covered by<br />
© 2011 <strong>The</strong> Linnean Society <strong>of</strong> London, Botanical Journal <strong>of</strong> the Linnean Society, 2011, 166, 20–39
ORCHID FLORA OF COCOS ISLAND 29<br />
Figure 4. Epidendrum cocoense Hágsater. A, habit. B, segment <strong>of</strong> the inflorescence. C, flower. D, dissected perianth. E,<br />
column and lip, lateral view. F, column, lateral and front views. Drawn by D. Bogarín from Bogarín 2767 (JBL-Spirit).<br />
© 2011 <strong>The</strong> Linnean Society <strong>of</strong> London, Botanical Journal <strong>of</strong> the Linnean Society, 2011, 166, 20–39
30 D. BOGARÍN ET AL.<br />
imbricate sheaths, foliaceous, becoming papyraceous<br />
with age. PEDICEL inconspicuous, < 0.5 mm long,<br />
<strong>flora</strong>l bracts widely ovate, longer than the ovary,<br />
scarious, conduplicate, acute to subacute, 2.5–<br />
3.3 ¥ 1.5–2.0 cm. OVARY cylindrical, to 2 ¥ 3 mm,<br />
scurfy. FLOWERS white–cream, approximately<br />
1.7 cm in length. DORSAL SEPAL subequal to the<br />
lateral sepals, rectangular–ovate to elliptic, acute,<br />
12 ¥ 5 mm. LATERAL SEPALS ovate, elliptic, acute,<br />
slightly mucronate, 10 ¥ 5 mm. PETALS rectangular–<br />
oblong, subacute, margins revolute, 10 ¥ 2.5 mm. LIP<br />
triangular to slightly cordate, with basal rounded<br />
lobes covering the column apex, subacute, callus<br />
basal, rectangular with erose apex touching the<br />
column apex and a central keel running to the lip<br />
apex, adnate to the column, 1.0 ¥ 0.7 cm. COLUMN<br />
short, straight, slightly alate, with sinuose thick<br />
wings, widely dilated towards the apical half in<br />
lateral position, with an minutely acute, triangular<br />
tooth on each side, 5 ¥ 4 mm, anther apical, stigma<br />
ventral. CLINANDRIUM reduced, erose, sinuose.<br />
NECTARY prominent, penetrating the ovary. POLLINIA<br />
four, ovoid in pairs, with caudicles. ANTHER CAP cucullate,<br />
four-celled. CAPSULE pyriform, 3.0 ¥ 1.5 cm.<br />
Other vouchers examined: COSTA RICA. Puntarenas:<br />
Puntarenas, Isla del Coco, camino al Cerro Iglesias,<br />
cerca de Llanos Palo de Hierro, 310 m, 5°31′55.3N,<br />
87°03′47.8″W, bosque pluvial premontano, transición<br />
a basal, epífitas en Sacoglottis holdridgei, 15 abril<br />
2006, D. Bogarín 2750, G. Blanco, G. Gigot, V. Savolainen<br />
& J. Warner (JBL-Spirit). Puntarenas: Puntarenas,<br />
Isla del Coco, orillas del Río Genio, camino a<br />
la represa, 30 m, 5°32′33.08″N, 87°03′10.3″W, bosque<br />
pluvial premontano, transición a basal, epífita en<br />
Sacoglottis holdridgei, 15 abril 2006, D. Bogarín<br />
2732, G. Blanco, G.Gigot, V. Savolainen & J. Warner<br />
(JBL-Spirit). Puntarenas: Puntarenas, Isla del Coco,<br />
orillas del Río Genio, cerca de la casa de máquinas<br />
de la planta hidroeléctrica, 15 m, 5°32′60.08″N,<br />
87°03′41.3″W, bosque pluvial premontano, transición<br />
a basal, epífitas, 15 abril 2006, D. Bogarín 2767, G.<br />
Blanco, G. Gigot, V. Savolainen & J. Warner (JBL-<br />
Spirit). Puntarenas: Puntarenas, Isla del Coco, orillas<br />
del Río Genio, 50 m a la izquierda de la Casa de<br />
Máquinas, 30 m, 5°32′31.4″N, 87°03′16.7″W, bosque<br />
pluvial premontano, transición a basal, 18 abril 2007,<br />
D. Bogarín 3704, G.Gigot & J.D. Zuñiga (JBL-Spirit).<br />
Wafer Bay, 14 August 1973, R.L. Dressler 4471 (CR).<br />
Isla del Coco, bosque de Wafer, 20 m, febrero 1970,<br />
L.D. Gómez 3270 (CR). Isla del Coco, Bahía wafer, c.<br />
nivel del mar, 31 Julio 1981, J. Gómez-Laurito 6914<br />
(CR). Wafer Bay, June 28, 1932, J.T. Howell 10190<br />
(CAS). <strong>Cocos</strong> <strong>Island</strong>, December 5, 1959, W. Klawe<br />
1519 (US). Lépiz 339 (INB). Sendero Cerro Iglesias,<br />
29 June 1997, A. Rojas 3691 (CR, INB). Bahía Wafer,<br />
21 enero 1980, P. Sánchez-Vindas 10 (CR). Isla del<br />
Coco, 6–8 junio 1989, R. Soto s.n. (USJ). <strong>Cocos</strong> <strong>Island</strong>,<br />
along brook flowing into Wafer Bay, on trees, 18 April<br />
1930, H. K. Svenson 333 (AMES, BKL, F). Isla del<br />
Coco, 5°32′N, 87°03′W, November 2002, J. Trusty 101,<br />
167, 176, 184, 194, 195, 363, 387, 461, 483, 494, 496,<br />
501, 510, 511, 512, 520, 544, 548, 560 and 561 (CR,<br />
FTG). Isla del Coco, marzo 1940, J. Valerio 1108 (CR).<br />
Distribution: known only from <strong>Cocos</strong> <strong>Island</strong> in Costa<br />
Rica. Most plants are found between sea level and<br />
250 m elevation, but some can reach 575 m elevation<br />
on Cerro Iglesias.<br />
Etymology: from the Spanish word ‘coco’, coconut the<br />
name <strong>of</strong> the island to which this species is endemic.<br />
Habitat and ecology: A common endemic epiphyte in<br />
premontane rainforest, basal belt transition. Plants<br />
are easily found on the common <strong>Cocos</strong> <strong>Island</strong> endemic<br />
Sacoglottis holdridgei. <strong>The</strong>y grow hanging, mostly<br />
(but not strictly) on the main tree trunks at 2 m or<br />
less above the ground in shade understorey conditions.<br />
However, other individuals can be found in<br />
exposed humid conditions around the Genio River,<br />
Wafer and Chatham Bays. <strong>The</strong>y are recorded across<br />
the island, but in some areas with scattered Clusia<br />
rosea trees and rocky cliffs such as Punta Presidio<br />
there were no plants.<br />
Phenology: Flowers are produced from December to<br />
August.<br />
Discussion: Among the <strong>orchid</strong>s <strong>of</strong> the island, E.<br />
cocoense can be distinguished by the large pendent<br />
plants up to 2.5 m long. It can be distinguished from<br />
E. insulanum and E. jimenezii by the larger main<br />
stems, its conspicuous broad <strong>flora</strong>l bracts to 1.3–<br />
2.0 cm wide (vs. less than 0.5–0.6 cm wide in E.<br />
insulanum and E. jimenezii) and the scurfy ovary<br />
5 mm in diameter (vs. a glabrescent ovary < 2mmin<br />
diameter). <strong>The</strong> leaves are wider and longer, up to<br />
2.7 cm wide and 12.5 cm long (vs. < 1.5 cm wide and<br />
8 cm long in the other Epidendrum spp.). Also, it<br />
differs from E. insulanum in the inflorescence having<br />
three or four white flowers, rarely two (vs. one yellow<br />
flower), the column with two apical teeth in each side<br />
(vs. erose column apex) and the lateral lobes <strong>of</strong> lip<br />
covering the column apex in lateral position (vs.<br />
lateral lobes <strong>of</strong> lip not covering the column apex). It<br />
differs from E. jimenezii by the less conspicuous teeth<br />
at the apex <strong>of</strong> the column, the subacute lip apex and<br />
the wider <strong>flora</strong>l bracts completely covering the ovary.<br />
© 2011 <strong>The</strong> Linnean Society <strong>of</strong> London, Botanical Journal <strong>of</strong> the Linnean Society, 2011, 166, 20–39
Conservation status: <strong>The</strong> first IUCN Red List Assessment<br />
<strong>of</strong> E. cocoense by Trusty (2004) estimated an<br />
extent <strong>of</strong> occurrence (EOO) <strong>of</strong> 22.3 km 2 . Based on<br />
fieldwork collections during a trip in 2006, the EOO<br />
was calculated as 4.5 km 2 and the area <strong>of</strong> occupancy<br />
(AOO) in 7 km 2 . However, as E. cocoense is a common<br />
epiphyte in the island, its distribution in the whole<br />
territory is expected, with the exception <strong>of</strong> few disturbed<br />
areas and cliffs along the coast. If threats are<br />
identified in the future, then a rating <strong>of</strong> Vulnerable<br />
(VU) under criterion D2 may be appropriate. In 2004,<br />
Trusty suggested classifying this species either as<br />
Critically Endangered (CR) or as VU because <strong>of</strong> criterion<br />
B and D. <strong>The</strong> current conservation status by<br />
IUCN is Near Threatened (NT).<br />
2. EPIDENDRUM INSULANUM SCHLTR., Beih. Bot.<br />
Centralbl., Abt. 36(2): 404. 1918. Type: Costa Rica.<br />
Cordon littoral à Wafer Bay, <strong>Cocos</strong> <strong>Island</strong> (Pacific<br />
Ocean), January 1902, H. Pittier (n. 16350 Herb.<br />
Nacion. Costa Rica). Lectotype GH-3580 (=AMES-<br />
73449), designated by Trusty & Blanco (Epidendrum<br />
27: 12–13. 2005) Isosyntypes B (destroyed, drawing in<br />
AMES, AMES-70447), CR (two sheets), AMES [two<br />
sheets, GH-3581 (=AMES-70445), GH-3579 (=AMES-<br />
70446). (Figs 3D, 5).<br />
Description: PLANT epiphytic, pendent, erect or<br />
suberect, with terete, leafy stem to 30 cm long. Roots<br />
thin, < 1.5 mm in diameter, white to greenish, with<br />
green tips, produced basally. STEM terete, branching,<br />
cane-like, with lateral, secondary stems shorter than<br />
the main stems, each stems ending in apical inflorescences.<br />
LEAF SHEATHS tubular, enclosing the stem,<br />
laterally flattened, to 1 cm long. LEAVES many (to 18),<br />
distichous, distributed throughout the stem, laterally<br />
twisted, lanceolate, elliptic–oblong, acute, with a<br />
bilobed apex, conduplicate, coriaceous, articulate with<br />
the sheath involving the stem, greenish to yellowish,<br />
to c. 5–6 ¥ 2.5–1.3 cm. INFLORESCENCE apical, racemose,<br />
distichous, one-flowered, shorter than the<br />
leaves, flowering once, to 5 cm long, covered by<br />
imbricate sheaths, foliaceous, papyraceous with age;<br />
pedicel inconspicuous, < 0.5 mm long. FLORAL BRACTS<br />
tubular, slightly larger than the ovary, scarious, acute<br />
to subacute, to 6 mm long. OVARY cylindric, to 5 cm<br />
long. FLOWERS yellowish to lemon, approximately<br />
6 mm in length. DORSAL SEPAL subequal to the lateral<br />
sepals, ovate to elliptic, subacute, 7.5 ¥ 2.3 mm.<br />
LATERAL SEPALS ovate to elliptic, acute, 7.5 ¥ 2.5 mm.<br />
PETALS anguste–oblong to lanceolate, acute, margins<br />
slightly revolute, 7.0 ¥ 1.2 mm. LIP subcordate, with<br />
short basal rounded lobes not covering the column<br />
apex and produced just after the apex <strong>of</strong> the column,<br />
acute, callus basal, triangular with touching the<br />
column apex and a central keel running to the lip<br />
apex, adnate to the column, 6.0 ¥ 2.5 mm. COLUMN<br />
ORCHID FLORA OF COCOS ISLAND 31<br />
very short, straight, almost tubular, without teeth,<br />
5.0 ¥ 3.3 mm, anther apical, stigma ventral. CLINAN-<br />
DRIUM reduced, erose, sinuose. NECTARY prominent,<br />
penetrating the ovary. POLLINIA four, ovoid in pairs,<br />
with caudicles. ANTHER CAP cucullate, four-celled.<br />
Other vouchers examined: COSTA RICA. Puntarenas:<br />
Puntarenas, Isla del Coco, Llanos Palo de Hierro,<br />
310 m, 5°31′55.3″N, 87°03′47.8″W, bosque pluvial premontano,<br />
transición a basal, epífitas en Sacoglottis<br />
holdridgei, 16 abril 2006, D. Bogarín 2772, G. Blanco,<br />
G.Gigot, V. Savolainen & J. Warner (JBL-Spirit). Puntarenas:<br />
Puntarenas, Isla del Coco, camino al Cerro<br />
Iglesias, cerca de Llanos Palo de Hierro, 310 m,<br />
5°31′55.3″N, 87°03′47.8″W, bosque pluvial premontano,<br />
transición a basal, epífitas en Sacoglottis holdridgei,<br />
15 abril 2006, D. Bogarín 2762, G. Blanco,<br />
G.Gigot, V. Savolainen & J. Warner (JBL-Spirit). Puntarenas:<br />
Puntarenas, Isla del Coco, orillas del Río<br />
Genio, 50 m a la izquierda de la Casa de Máquinas,<br />
30 m, 5°32′31.4″N, 87°03′16.7″W, bosque pluvial premontano,<br />
transición a basal, 18 abril 2007, D. Bogarín<br />
3707, G. Gigot & J.D. Zuñiga (JBL-Spirit). Same<br />
locality: D. Bogarín 3705, 3708, 3702, 3706. (JBL-<br />
Spirit). <strong>Cocos</strong> <strong>Island</strong>, 28 march 1891, A. Agassiz s.n.<br />
(GH) near Wafer Bay, 14 august 1973, R.L. Dressler<br />
4468 (CR, PMA). 9–11 April 1979, R.B Foster 4117 (F,<br />
US). Isla del Coco, 31 July 1981, J. Gómez-Laurito<br />
6924 (CR). Río Genio, Estación Wafer, 1–10 m, 14<br />
abril 1994, E. Lépiz 358 (INB). Cerro Iglesias, 200–<br />
250 m, 22 Junio 1997, A. Rojas 3660 (F, INB). Wafer<br />
Bay, 18 April 1930, H. Svenson 335 (AMES, BKL, F,<br />
K, LE, S, UC). Isla del Coco, 5°31′N, 87°03′W,<br />
J. Trusty 53, 158, 218 and 500 (CR, USJ).<br />
Distribution: known only from <strong>Cocos</strong> <strong>Island</strong>, Costa<br />
Rica.<br />
Etymology: From the Latin insula, ‘island’, in allusion<br />
to <strong>Cocos</strong> <strong>Island</strong>, to which this species is endemic.<br />
Phenology: Apparently they flower throughout the<br />
year. However, plants collected and cultivated for this<br />
study flowered from March to July.<br />
Habitat and ecology: A common endemic epiphyte<br />
in premontane rainforest, basal belt transition. <strong>The</strong><br />
habitat is mostly the same as for E. cocoense and<br />
sometimes they grow intermixed. <strong>The</strong>y are pendent or<br />
suberect, growing on exposed or shady conditions in<br />
both shrubs and trees. Most <strong>of</strong> the plants are found<br />
between sea level and 300 m <strong>of</strong> elevation; however,<br />
they can reach 575 m on Cerro Iglesias. Pollinators<br />
are unknown. <strong>The</strong> three Epidendrum spp. have<br />
similar <strong>flora</strong>l architecture. As in E. cocoense and E.<br />
jimenezii, plants could be pollinated by Lepidoptera.<br />
© 2011 <strong>The</strong> Linnean Society <strong>of</strong> London, Botanical Journal <strong>of</strong> the Linnean Society, 2011, 166, 20–39
32 D. BOGARÍN ET AL.<br />
Figure 5. Epidendrum insulanum Schltr. A, habit. B, segment <strong>of</strong> the inflorescence. C, flower. D, dissected perianth. E,<br />
column and lip, lateral view. F, column, front view. Drawn by D. Bogarín from Bogarín 2772 (JBL-Spirit).<br />
© 2011 <strong>The</strong> Linnean Society <strong>of</strong> London, Botanical Journal <strong>of</strong> the Linnean Society, 2011, 166, 20–39
Discussion: Epidendrum insulanum is easily distinguished<br />
by the inflorescences having only one, green–<br />
yellow flower (vs. whitish to white, and two or four in<br />
E. jimenezii and E. cocoense, respectively). <strong>The</strong> apex<br />
<strong>of</strong> column is erose, lacking the two peaks present in<br />
the other species. <strong>The</strong> lateral lobes <strong>of</strong> lip do not cover<br />
the apex <strong>of</strong> column in lateral view. <strong>The</strong> lip is smaller,<br />
up to 6 mm long. Sepals and petals are smaller,<br />
< 1 cm in length. Vegetatively, plants are smaller than<br />
those <strong>of</strong> E. cocoense, reaching up to 30 cm long with<br />
leaves < 1.5 cm wide. Some larger plants can be confused<br />
with E. jimenezii because <strong>of</strong> the similar size.<br />
Conservation status: In 2004, Trusty suggested classifying<br />
this species either as Critically Endangered<br />
CR or as VU because <strong>of</strong> criterion B and D. Based on<br />
herbarium specimens and field collections, she estimated<br />
an EOO <strong>of</strong> 18.2 km 2 . Based on field collections<br />
performed in 2006, the EOO was calculated as 3 km 2<br />
and the AOO as 7 km 2 . Although restricted in range<br />
and variation in EOO and AOO values, based on field<br />
observations it is highly probable that plants <strong>of</strong> this<br />
species are found on the whole island. <strong>The</strong>y are not<br />
restricted to a particular place. <strong>The</strong> current conservation<br />
status by IUCN is Near Threatened (NT).<br />
3. EPIDENDRUM JIMENEZII HÁGSATER, Icon. Orchid.<br />
(Mexico) 3: pl. 341. 1999. Type: Costa Rica. <strong>Cocos</strong><br />
<strong>Island</strong>, Wafer Bay, Arroyo del Genio, 13 April 1965, A.<br />
Jiménez M. 3178 (holotype: F isotypes: CR, MO, SEL,<br />
U) (Figs 3E, 6).<br />
Description: PLANT epiphytic, pendent, with terete,<br />
leafy stem to 35 cm long. ROOTS thin, < 1.5 mm in<br />
diameter, white to greenish, with green tips, produced<br />
basally. STEM terete, branching, cane-like, with<br />
lateral, secondary stems shorter than the main stems,<br />
each stem ending in apical inflorescences. LEAF<br />
SHEATHS tubular, enclosing the stem, laterally flattened,<br />
to 2 cm long. LEAVES many (to 20), distichous,<br />
distributed throughout the stem, laterally twisted,<br />
lanceolate, elliptic–oblong, acute, with a bilobed apex,<br />
conduplicate, coriaceous to subcoriaceous, articulate<br />
with the sheath involving the stem, to approximately<br />
6–8 ¥ 1.0–1.3 cm. INFLORESCENCE apical, racemose,<br />
distichous, two-flowered, shorter than the leaves,<br />
flowering once, to 3.1 cm long, covered by imbricate<br />
sheaths, foliaceous, becoming papyraceous with<br />
age. PEDICEL inconspicuous, < 0.5 mm long. FLORAL<br />
BRACTS tubular, as long as the ovary, scarious, acute<br />
to subacute, 0.5–0.6 cm long. OVARY cylindric, to<br />
1.5 cm long. FLOWERS whitish, about 1.3 cm in length.<br />
DORSAL SEPAL subequal to the lateral sepals, ovate to<br />
elliptic, acute, 9.5 ¥ 3.0 mm. LATERAL SEPALS ovate to<br />
elliptic, acute, 9.5 ¥ 3.0 mm. PETALS rectangular–<br />
oblong to lanceolate, acute to subacute, margins revolute,<br />
8 ¥ 2 mm. LIP cordate, with basal rounded lobes<br />
ORCHID FLORA OF COCOS ISLAND 33<br />
covering the column apex, acute, callus basal, rectangular<br />
with erose apex touching the column apex and<br />
a central keel running to the lip apex, adnate to the<br />
column, 7 ¥ 4 mm. COLUMN short, straight, slightly<br />
alate, with sinuose wings, widely dilated towards<br />
the apical half in lateral position, with an acute,<br />
triangular tooth on each side, 5.0 ¥ 2.2 mm, anther<br />
apical, stigma ventral. CLINANDRIUM reduced,<br />
sinuose. NECTARY prominent, penetrating the ovary.<br />
POLLINIA four, ovoid in pairs, with caudicles. ANTHER<br />
CAP cucullate, four-celled.<br />
Other vouchers examined: COSTA RICA. Puntarenas:<br />
Puntarenas, Isla del Coco, orillas del Río Genio,<br />
camino a la represa, 30 m, 5°32′33.08N, 87°03′10.3″W,<br />
bosque pluvial premontano, transición a basal,<br />
epífita, 13 abril 2006, D. Bogarín 2735, G. Blanco,<br />
G.Gigot, V. Savolainen & J. Warner (JBL-Spirit).<br />
<strong>Cocos</strong> <strong>Island</strong>, Wafer Bay, Arroyo del Genio, 13 April<br />
1965, A. Jiménez M. 3178 (CR); Wafer Bay, R.L.<br />
Dressler 4468 (FLAS); <strong>Cocos</strong> <strong>Island</strong>, R. Foster 4114<br />
(US).<br />
Distribution: Known only from <strong>Cocos</strong> <strong>Island</strong>, Costa<br />
Rica. It is restricted to the Genio River and Wafer Bay<br />
areas.<br />
Eponymy: Named in honour <strong>of</strong> Ing. Alfonso Jiménez<br />
Muñoz, ex-pr<strong>of</strong>esor <strong>of</strong> Agronomy and Biology at Universidad<br />
Nacional de Costa Rica and ex-director <strong>of</strong><br />
the Museo Nacional de Costa Rica, who collected the<br />
type specimen.<br />
Phenology: December to January.<br />
Habitat and ecology: a very rare endemic epiphytic<br />
<strong>orchid</strong> in premontane rainforest, basal belt transition.<br />
Only five specimens are known from the island and<br />
their ecology is poorly understood. As noted for the<br />
other Epidendrum spp., the <strong>flora</strong>l architecture is<br />
similar and their pollinators could share the same<br />
characteristics. <strong>The</strong> only plant found in this study<br />
was growing along the path to the Genio River waterfall,<br />
in shady humid conditions.<br />
Discussion: Plants <strong>of</strong> this species are distinguished by<br />
the inflorescence producing two whitish flowers, the<br />
cordate, acute lip and the column with two acute<br />
teeth at apex. Compared with E. cocoense, it can be<br />
distinguished by its smaller habit, up to 35 cm long,<br />
the leaves 1.0–1.2 wide, the less conspicuous tubular<br />
<strong>flora</strong>l bracts 0.5–0.6 cm wide and as long as the glabrescent<br />
ovary. Vegetatively it is similar to some<br />
larger plants <strong>of</strong> E. jimenezii. Large plants <strong>of</strong> E. insulanum<br />
are <strong>of</strong>ten indistinguishable from those <strong>of</strong><br />
E. jimenezii. However, plants <strong>of</strong> E. jimenezii can be<br />
© 2011 <strong>The</strong> Linnean Society <strong>of</strong> London, Botanical Journal <strong>of</strong> the Linnean Society, 2011, 166, 20–39
34 D. BOGARÍN ET AL.<br />
Figure 6. Epidendrum jimenezii Hágsater. A, habit. B, segment <strong>of</strong> the inflorescence. C, flower. D, dissected perianth. E,<br />
column and lip, lateral view. F, column, lateral and front views. Drawn by D. Bogarín from Bogarín 2735 (JBL-Spirit).<br />
© 2011 <strong>The</strong> Linnean Society <strong>of</strong> London, Botanical Journal <strong>of</strong> the Linnean Society, 2011, 166, 20–39
distinguished by the inflorescences having two<br />
whitish flowers, > 1 cm in length, the column apex<br />
with two teeth in each side and the lateral lobes <strong>of</strong> lip<br />
covering the column apex.<br />
<strong>The</strong> <strong>flora</strong>l features <strong>of</strong> the three Epidendrum spp.<br />
are similar. <strong>The</strong> only specimen collected for this study<br />
seems to be intermediate between E. cocoense and<br />
E. insulanum. Basically, the lip shape is similar to<br />
E. cocoense, with the same callus structure, but<br />
having rounded basal lobes and acute apex as in<br />
E. insulanum. <strong>The</strong> sepals and petals are as long as<br />
those <strong>of</strong> E. cocoense, but the shape is more similar to<br />
E. insulanum. Plant size is intermediate between the<br />
two species. <strong>The</strong> column shape also shares intermediate<br />
characteristics. <strong>The</strong> column is oblong as in<br />
E. insulanum but with two teeth at apex as in<br />
E. cocoense.<br />
Trusty (2006) was unable to distinguish this species<br />
from E. insulanum. <strong>The</strong>y suggested the possibility<br />
that this species could be a hybrid between E.<br />
cocoense and E. insulanum and observations <strong>of</strong> <strong>flora</strong>l<br />
morphology support their hypothesis. However, more<br />
evidence is needed to complement the morphological<br />
data, especially at population genetics and reproductive<br />
biology levels.<br />
Conservation status: Trusty (2004) suggested classifying<br />
this species as CR because <strong>of</strong> criterion B and D.<br />
<strong>The</strong> estimation <strong>of</strong> the EOO is < 1km 2 . This species is<br />
very rare and restricted to the Wafer Bay areas<br />
around the Genio River where it is known from less<br />
than five collections. This species has not been evaluated<br />
yet under IUCN criteria; however, it should be<br />
classified as CR as suggested by Trusty (2004).<br />
<strong>The</strong> endemic Epidendrum species: <strong>The</strong> three Epidendrum<br />
spp. are similar in their vegetative morphology.<br />
All the species have a pendent branching habit. <strong>The</strong><br />
inflorescence is terminal and new branches are produced<br />
beneath the apex after they have flowered. <strong>The</strong><br />
new branches flower again. Flowers are superficially<br />
similar. <strong>The</strong> lip is cordate or subcordate with the<br />
callus always Y-shaped. <strong>The</strong> column has a reduced<br />
clinandrium and shows little variation between<br />
species. <strong>The</strong> perianth segments show some variation<br />
in size and their shape varies from oblong to<br />
ovate–elliptic.<br />
Relationships with other members <strong>of</strong> the genus<br />
Epidendrum: According to Hágsater (2007), they are<br />
members <strong>of</strong> the Epidendrum ramosum Jacq. group,<br />
which is characterized by the monopodial branching<br />
stems, the spike-like distichous inflorescence and the<br />
lip with a single callus and the flexicaule subgroup,<br />
ORCHID FLORA OF COCOS ISLAND 35<br />
which has a straggling habit, the main stem not very<br />
evident and the dorsal keel present in the sepals,<br />
which is generally prominent.<br />
Epidendrum insulanum is closely related to E.<br />
flexicaule Schltr., E. stevensii Hágsater, E. modestiflorum<br />
Schltr and E. veraguasense Hágsater (Hágsater,<br />
2007), but they can be distinguished: E. flexicaule has<br />
longer leaves (up to 8.2 cm long) and two or three<br />
larger flowers, E. stevensii has two or three smaller<br />
flowers, E. modestiflorum shorter wider leaves<br />
(5 ¥ 15 cm) and two or three flowers and E. veraguasense<br />
has two or three white, larger flowers. Epidendrum<br />
jimenezii is perhaps closely allied to E.<br />
insulanum and E. cocense. Being sympatric and<br />
having similar plant size, E. jimenezii and E. insulanum<br />
are sometimes confused.<br />
Epidendrum cocoense is vegetatively similar to E.<br />
rafael-lucasii Hágsater, E. santaclarense Ames and E.<br />
acunae Dressler. <strong>The</strong>y are found in Central America<br />
and Panama, with the exception <strong>of</strong> the Costa Rican<br />
endemic E. rafael-lucasii. According to Hágsater<br />
(1999a), E. rafael-lucasii has larger yellow flowers (vs.<br />
white), sepals 16–20 mm long (vs. 12 mm in E.<br />
cocoense) and the column is blunt at the apex (vs.<br />
with two teeth at each side <strong>of</strong> apex). Epidendrum<br />
santaclarense has deep green flowers (vs. white) and<br />
E. acunae has shorter stems and very short flowering<br />
stems.<br />
Pollination <strong>of</strong> endemic species <strong>of</strong> Epidendrum: <strong>The</strong><br />
pollination biology is unknown. Epidendrum spp.<br />
have a deep tube inside the ovary, sometimes with<br />
free nectar. <strong>The</strong> lip and column made up a simple<br />
horizontal tube, with the free part <strong>of</strong> the lip used as<br />
an appendage for pollinator attraction. Some species<br />
<strong>of</strong> Lepidoptera, mainly moths, are important pollinators<br />
<strong>of</strong> members <strong>of</strong> subtribe Laeliinae Benth., including<br />
Epidendrum (van der Pijl & Dodson, 1966). <strong>The</strong><br />
white to whitish flowers <strong>of</strong> E. cocoense produce a<br />
perceptible sweet smell and they could attract some<br />
species <strong>of</strong> moths or butterflies. Nearly 100 species<br />
<strong>of</strong> Lepidoptera are recorded from <strong>Cocos</strong> <strong>Island</strong> and<br />
moths are the largest group (Brown, Donahue &<br />
Miller, 1991; Brown & Miller, 1999). As there are no<br />
bees in the island, it is highly probable that the<br />
Epidendrum spp. are pollinated by Lepidoptera. Selfpollination<br />
was not observed and plants in cultivation<br />
flowered without producing fruits.<br />
3. ORNITHIDIUM SALISB. EX R.BR.<br />
ORNITHIDIUM SALISB. EX R.BR., Hort. Kew. ed. 2, 5:<br />
210. 1813. Type species: Epidendrum coccineum Jacq.,<br />
Enum. Syst. Pl. 29. 1760. = Ornithidium coccineum<br />
(Jacq.) Salisb., Trans. Hort. Soc. London 1: 293. 1812.<br />
EPIPHYTIC, sympodial (caespitose to rhizomatous)<br />
or monopodial plants; rarely with dimorphic growth.<br />
© 2011 <strong>The</strong> Linnean Society <strong>of</strong> London, Botanical Journal <strong>of</strong> the Linnean Society, 2011, 166, 20–39
36 D. BOGARÍN ET AL.<br />
<strong>The</strong> thick roots with an orangish coloration. STEMS<br />
cane-like with distichous leaves or with pseudobulbs<br />
bearing basal and apical leaves. <strong>The</strong> stems and leaves<br />
<strong>of</strong> most species have an olive green coloration. LEAVES<br />
conduplicate, coriaceous to subcoriaceous. INFLORES-<br />
CENCES usually fascicled, pedicel and ovary longer<br />
than the <strong>flora</strong>l bract. FLOWERS usually small, fleshy,<br />
campanulate or subglobose, the perianth lacks fibres;<br />
many species have yellow, orange, red, tan, green or<br />
pink flowers. SEPALS AND PETALS subsimilar, spreading.<br />
LIP simple or three- or four-lobed, articulated<br />
with or adnate to the column, usually provided at the<br />
base with a fleshy callus and <strong>of</strong>ten producing nectar<br />
at the base <strong>of</strong> the labellum. COLUMN erect, terete.<br />
CAPSULES have apical dehiscence.<br />
Ornithidium is a widely distributed genus throughout<br />
the Neotropics with approximately 60 species.<br />
1. ORNITHIDIUM ADENDROBIUM (RCHB.F.)<br />
M.A.BLANCO &OJEDA, Lankesteriana 7: 532. 2007.<br />
Ponera adendrobium Rchb.f., Flora 48: 278. 1865.<br />
Pleuranthium adendrobium (Rchb.f.) Benth. &<br />
Hook.f. ex B.D.Jacks., Index Kew. 2: 562. 1894. Neourbania<br />
adendrobium (Rchb.f.) Fawc. & Rendle, J.<br />
Bot. 47: 125 1909. Type: CUBA. 1860–1864, C. H.<br />
Wright 1697 (holotype: W; isosintypes: AMES, BM, K,<br />
MO). Camaridium parviflorum Fawc. in I. Urban,<br />
Symb. Antill. 1: 472. 1900. (Figs 3F, 7).<br />
Description: PLANT epiphytic, erect or suberect, with<br />
terete, leafy stem, to 60 cm long. ROOTS flexuous,<br />
less < 1.5 mm in diameter, orangish to whitish. STEM<br />
without pseudobulbs, terete, leafy, covered by<br />
addressed leaf sheaths, scarious at base, foliaceous<br />
towards the apex, producing secondary branches,<br />
with adventitious roots produced at bases <strong>of</strong> leaf<br />
nodes. LEAVES many (to 25), distichous, narrowly<br />
oblong to anguste–elliptic, acute, conduplicate, fleshy,<br />
flexible, articulate with the sheath involving the<br />
stem, to c. 11.0 ¥ 1.6 cm. INFLORESCENCE a singleflowered<br />
raceme usually larger than the leaves, produced<br />
along the stem, in the axils <strong>of</strong> the leaves, c.<br />
2 cm long. PEDICEL inconspicuous, < 0.5 mm long.<br />
FLORAL BRACTS ovate, foliaceous. OVARY cylindric,<br />
< 4 mm long, glabrous. FLOWERS small, inconspicuous,<br />
c. 6 mm in length, yellowish to cream–yellow<br />
or greenish. DORSAL SEPAL subequal to the lateral<br />
sepals, oblong–elliptic, acute or slightly acuminate,<br />
6 ¥ 2 mm. LATERAL SEPALS oblong–elliptic, acute,<br />
subfalcate, 6 ¥ 2 mm. PETALS ovate–lanceolate, acute,<br />
parallel to the column, 5.0 ¥ 1.7 mm. LIP ovate, lobed,<br />
sinuose, retuse, margin erose, callous laminar in the<br />
centre, with two parallel keels running to the apex,<br />
attached to the column foot. COLUMN short, terete,<br />
slightly arcuate, to 3 mm long; stigma ventral, widely<br />
obovate, anther terminal. POLLINIA two. ANTHER CAP,<br />
cucullate. CAPSULE pyriform to 1.7 cm long.<br />
Other vouchers examined: COSTA RICA. Puntarenas:<br />
Puntarenas, Isla del Coco, camino al Cerro Iglesias,<br />
cerca de Llanos Palo de Hierro, 310 m, 5°31′55.3″N,<br />
87°03′47.8″W, bosque pluvial premontano, transición<br />
a basal, epífitas en Sacoglottis holdridgei, 15 abril<br />
2006, D. Bogarín 2755, G. Blanco, G. Gigot, V. Savolainen<br />
& J. Warner (JBL-Spirit). Same locality, D.<br />
Bogarín 2759 (JBL-Spirit). Puntarenas, Isla del Coco,<br />
December 2003, J. Trusty 495 (JBL-Spirit). Same<br />
locality: J. Trusty 52 (JBL-Spirit). Isla del Coco: Wafer<br />
Bay, 14 august 1973, R.L. Dressler 4470 (CR). Isla del<br />
Coco, J. Trusty 52, 100, 157, 495, 515, 556, 564 (CR).<br />
december 2003, J. Trusty 52 and 495 (JBL-spirit). Isla<br />
del Coco, región de los Llanos y subiendo a Cerro<br />
Pelón, 5°32′15″N 87°03′29″W, 200–250 m, 29 junio<br />
1997, A. Rojas 3692 (INB). 6–8 junio 1989, R. Soto<br />
s.n. (USJ-49286).<br />
Distribution: This is a widespread species ranging<br />
from Mexico and the Antilles throughout Central and<br />
South America.<br />
Etymology: Made up by the Latin prefix ‘a’, ‘away<br />
from’ and the name <strong>of</strong> the genus Dendrobium. According<br />
to Reichenbach (1865), the specimen he studied<br />
‘has all characteristics <strong>of</strong> Dendrobium, therefore the<br />
name’.<br />
Phenology: <strong>The</strong> plants have been recorded in flower<br />
throughout the year, mostly between September and<br />
April<br />
Habitat and ecology: Plants are frequent in the<br />
island. <strong>The</strong>y are mostly found from 250 to 585 m <strong>of</strong><br />
elevation, up to the top <strong>of</strong> Cerro Iglesias. It is rarely<br />
found at sea level. Some plants are probably selfpollinated<br />
because they form capsules quickly and<br />
regularly in almost all flowers. In the field, small ants<br />
were observed around the flowers but without pollinia.<br />
Capsules are seen throughout the year.<br />
Discussion: Plants have distichously arranged leaves<br />
without pseudobulbs. Compared with Epidendrum<br />
spp., it is easily recognized by the lateral singleflowered<br />
inflorescences and the footed column. It is<br />
distinguished from C. micranthum by its larger habit,<br />
without pseudobulbs and the stem with distichously<br />
arranged leaves. <strong>The</strong> small flowers do not have the<br />
lip adnate to the column foot. This species was previously<br />
treated in Maxillaria; however, nomenclatural<br />
changes in Maxillariinae suggest its inclusion in<br />
Ornithidium (Blanco et al., 2007; Whitten et al.,<br />
2009).<br />
© 2011 <strong>The</strong> Linnean Society <strong>of</strong> London, Botanical Journal <strong>of</strong> the Linnean Society, 2011, 166, 20–39
ORCHID FLORA OF COCOS ISLAND 37<br />
Figure 7. Ornithidium adendrobium (Rchb.f.) M.A.Blanco & Ojeda. A, habit. B, flowers. C, dissected perianth. D, column<br />
and lip, lateral view. E, column, lateral and front view. Drawn by D. Bogarín from Bogarín 2755 (JBL-Spirit).<br />
© 2011 <strong>The</strong> Linnean Society <strong>of</strong> London, Botanical Journal <strong>of</strong> the Linnean Society, 2011, 166, 20–39
38 D. BOGARÍN ET AL.<br />
Conservation status: This species has a wide distribution<br />
range throughout the Neotropics. It is common<br />
in the island. It should not be regarded as endangered<br />
and may well fit the LC category for widespread and<br />
abundant taxa.<br />
ACKNOWLEDGEMENTS<br />
<strong>The</strong> present paper is part <strong>of</strong> the Project 814-BO-245,<br />
‘La <strong>flora</strong> de orquídeas del Parque Nacional Isla del<br />
Coco’ sponsored by the Vice Presidency <strong>of</strong> Research,<br />
University <strong>of</strong> Costa Rica and ‘Conservation and Monitoring<br />
<strong>of</strong> Meso-American Orchids’ supported by the<br />
Darwin Initiative, Department for Environment Food<br />
and Rural Affairs (DEFRA), UK. We thank the Costa<br />
Rican Ministry <strong>of</strong> Environment and Energy (MINAE),<br />
the <strong>National</strong> System <strong>of</strong> Conservation Areas (SINAC)<br />
throughout Área de Conservación Marina Isla del<br />
Coco (ACMIC) for issuing the Resolution Permit no.<br />
002-06 and the Scientific Passport no. 1281, under<br />
which the material was collected and studied, Mario<br />
Blanco and Robert L. Dressler for useful discussions<br />
and digital images <strong>of</strong> critical specimens, the curators<br />
<strong>of</strong> AMES, BM, CAS, CR, INB, JBL, K, PMA and USJ,<br />
the crew <strong>of</strong> Okeanos for providing transportation and<br />
facilities to the island, the park guards and personnel<br />
<strong>of</strong> <strong>Cocos</strong> <strong>Island</strong> <strong>National</strong> <strong>Park</strong>, especially Isaac Chinchilla,<br />
German Haugh and Lourdes Vargas for granting<br />
access to the island and help with fieldwork<br />
logistics, Guillermo Blanco who shared his knowledge<br />
and most <strong>of</strong> the fieldwork and Guillaume Gigot and<br />
Jose Daniel Zúñiga for helping during the fieldwork<br />
activities.<br />
REFERENCES<br />
Alfaro EJ. 2008. Ciclo diario y anual de variables troposféricas<br />
y oceánicas en la Isla del Coco, Costa Rica. Revista de<br />
Biología Tropical 56: 19–29.<br />
Bellon H, Sáenz R, Tournon J. 1984. K-Ar radiometric ages<br />
<strong>of</strong> lavas from <strong>Cocos</strong> <strong>Island</strong> (Eastern Pacific). Marine Geology<br />
54: M17–M23.<br />
Bentham G. 1844–1846. <strong>The</strong> botany <strong>of</strong> the voyage <strong>of</strong> H.M.S.<br />
Sulphur, under command <strong>of</strong> Captain Sir Edward Belcher,<br />
during the years 1836–42. Published under the authority <strong>of</strong><br />
the Lords Commissioners <strong>of</strong> the Admiralty. London, UK. Vol.<br />
12.<br />
Blanco MA, Carnevali G, Whitten WM, Singer RB,<br />
Koehler S, Williams NH, Ojeda I, Neubig KM, Endara<br />
L. 2007. Generic realignments in Maxillariinae (Orchidaceae).<br />
Lankesteriana 7: 515–537.<br />
Bogarín D, Pupulin F. 2007. Las orquídeas del Parque<br />
Nacional Barra Honda, Guanacaste, Costa Rica. Lankesteriana<br />
7: 446–449.<br />
Brown JW, Miller SE. 1999. A new species <strong>of</strong> Coelostathma<br />
Clemens (Lepidoptera: Tortricidae) from <strong>Cocos</strong> <strong>Island</strong>, Costa<br />
Rica, with comments on the phylogenetic significance <strong>of</strong><br />
abdominal dorsal pits in Sparganothini. Proceedings <strong>of</strong> the<br />
Entomological Society <strong>of</strong> Washington 101: 701–707.<br />
Brown JW, Donahue JP, Miller SE. 1991. Two new species<br />
<strong>of</strong> geometrid moths (Lepidoptera: Geometridae: Ennominae)<br />
from <strong>Cocos</strong> <strong>Island</strong>, Costa Rica. Natural History Museum<br />
<strong>of</strong> Los Angeles County, Contributions in Science 423: 11–18.<br />
Cortés J. 2008. Historia de la investigación marina de la Isla<br />
del Coco, Costa Rica. Revista de Biología Tropical 56: 1–18.<br />
Dressler RL. 2003. Orchidaceae. In: Hammel BE, Grayum<br />
MH, Herrera C, Zamora N, eds. Manual de plantas de Costa<br />
Rica. Vol. 3. Monocotiledóneas (Orchidaceae–Zingiberaceae).<br />
Monographs in Systematic Botany from the Missouri Botanical<br />
Garden. St. Louis, MO: Missouri Botanical Garden<br />
Press, 93:1–595.<br />
Garay LA. 1967. Studies in American <strong>orchid</strong>s VI. Botanical<br />
Museum Leaflets 21: 249–264.<br />
Hágsater E. 1999a. Epidendrum cocoënse Hágsater. Sub. pl.<br />
325. In: Hágsater E, Sánchez L, García-Cruz J, eds. Icones<br />
Orchidacearum (Mexico) 3. <strong>The</strong> genus Epidendrum: part 2.<br />
A second century <strong>of</strong> new species in Epidendrum. Mexico:<br />
Herbario AMO, 301–400.<br />
Hágsater E. 1999b. Epidendrum jimenezii Hágsater. Sub. pl.<br />
341. In: Hágsater E, Sánchez L, García-Cruz J, eds. Icones<br />
Orchidacearum (Mexico) 3. <strong>The</strong> genus Epidendrum: part 2.<br />
A second century <strong>of</strong> new species in Epidendrum. Mexico:<br />
Herbario AMO, 301–400.<br />
Hágsater E. 2007. Epidendrum insulanum Hágsater. Sub.<br />
948. In: Hágsater E, Sánchez L, eds. Icones Orchidacearum<br />
(Mexico) 9. <strong>The</strong> genus Epidendrum: part 6. Species new and<br />
old in Epidendrum. Mexico: Herbario AMO, 901–1000.<br />
Hogue CL, Miller SE. 1981. Entom<strong>of</strong>auna <strong>of</strong> <strong>Cocos</strong> <strong>Island</strong>,<br />
Costa Rica. Atoll Research Bulletin 250: 1–29.<br />
Holdridge LR. 1967. Life zone ecology. San José: Tropical<br />
Science Center.<br />
Holdridge LR. 1987. Ecología basada en zonas de vida, 3rd<br />
edn. San José: IICA.<br />
Lizano OG. 2008. Dinámica de aguas alrededor de la Isla del<br />
Coco, Costa Rica. Revista de Biología Tropical 56: 31–48.<br />
van der Pijl L, Dodson CH. 1966. Orchid flowers: their<br />
pollination and evolution. Coral Gables: University <strong>of</strong><br />
Miami Press.<br />
Pittier HF. 1898. Apuntamientos preliminares sobre la Isla<br />
del <strong>Cocos</strong>, posesión costarricense en el Océano Pacifico.<br />
Revista del Colegio Superior de Señoritas 2: 2–11.<br />
Pupulin F. 1998. Orchid florula <strong>of</strong> Parque Nacional Manuel<br />
Antonio, Quepos, Costa Rica. Revista de Biología Tropical<br />
46: 961–10317.<br />
Pupulin F. 2007. <strong>Epidendra</strong>, the botanical databases <strong>of</strong><br />
Jardín Botánico Lankester at the University <strong>of</strong> Costa Rica.<br />
Lankesteriana 7: 178–180.<br />
Pupulin F. 2009. <strong>Epidendra</strong>, la base de datos en línea del<br />
Jardín Botánico Lankester de la Universidad de Costa Rica.<br />
Orquideología XXVI: 89–94.<br />
Pupulin F. 2010. Orchidaceae werckleanae: typification <strong>of</strong><br />
Costa Rican <strong>orchid</strong> species described from collections by K.<br />
Wercklé. Botanical Journal <strong>of</strong> the Linnean Society 163:<br />
111–154.<br />
Reichenbach HG. 1865. Vorstudien zu einer Orchidographie<br />
© 2011 <strong>The</strong> Linnean Society <strong>of</strong> London, Botanical Journal <strong>of</strong> the Linnean Society, 2011, 166, 20–39
der Antillen, besonders britischen Antheils. Neuheiten und<br />
eingehende Critiken bisheriger Literatur I. Neuheiten aus<br />
Cuba von Wright. Flora 48: 273–280.<br />
Schlechter R. 1918. Kritische Aufzählung der bisher aus<br />
Zentralamerika bekanntgeworden Orchideen. Beihefte zum<br />
Botanischen Centralblatt, Kassel 36: 404.<br />
Singer RA. 2003. Orchid pollination: recent developments<br />
from Brazil. Lankesteriana 7: 111–114.<br />
Svenson HK. 1935. Plants <strong>of</strong> the Astor Expedition, 1930<br />
(Galápagos and <strong>Cocos</strong> <strong>Island</strong>s). American Journal <strong>of</strong> Botany<br />
22: 208–277.<br />
Tosi J. 1969. Mapa ecológico según la clasificación de zonas de<br />
vida del mundo de L. R. Holdridge. San José: Instituto<br />
Geográfico Nacional.<br />
Trusty J. 2004. Plant biogeography and conservation on a<br />
ORCHID FLORA OF COCOS ISLAND 39<br />
tropical island: Isla del Coco, Costa Rica. D. Phil. <strong>The</strong>sis,<br />
Florida International University.<br />
Trusty J, Blanco MA. 2005. Las orquídeas de la Isla del<br />
Coco. Epidendrum 27: 10–15.<br />
Trusty J, Blanco MA. 2006. Orchids <strong>of</strong> Isla del Coco: five<br />
species native to an island in Costa Rica. Orchids 75:<br />
826–831.<br />
Trusty J, Kesler HC, Haug Delgado G. 2006. Vascular <strong>flora</strong><br />
<strong>of</strong> Isla del Coco, Costa Rica. Proceedings <strong>of</strong> the California<br />
Academy Sciences 57: 247–355.<br />
Whitten WM, Blanco MA, Williams NH, Koehler S,<br />
Carnevali G, Singer RB, Endara L, Neubig KM. 2009.<br />
Molecular phylogenetics <strong>of</strong> Maxillaria and related genera<br />
(Orchidaceae: Cymbidiae) based on combined molecular<br />
data sets. American Journal <strong>of</strong> Botany 94: 1860–1889.<br />
© 2011 <strong>The</strong> Linnean Society <strong>of</strong> London, Botanical Journal <strong>of</strong> the Linnean Society, 2011, 166, 20–39