Seed surface morphology in some representatives of the Genus ...
Seed surface morphology in some representatives of the Genus ... Seed surface morphology in some representatives of the Genus ...
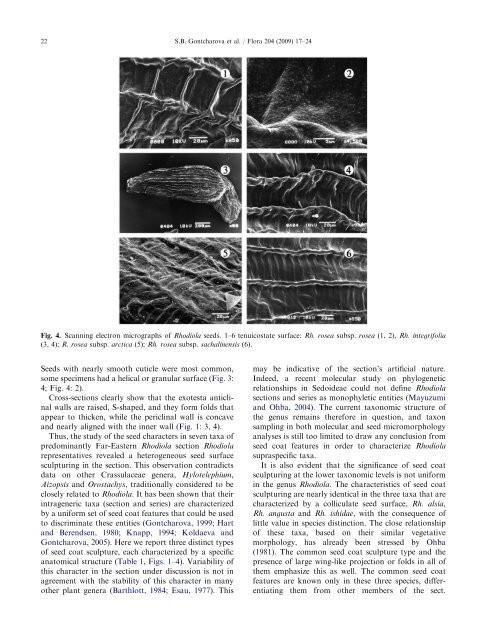
22 Seeds with nearly smooth cuticle were most common, some specimens had a helical or granular surface (Fig. 3: 4; Fig. 4: 2). Cross-sections clearly show that the exotesta anticlinal walls are raised, S-shaped, and they form folds that appear to thicken, while the periclinal wall is concave and nearly aligned with the inner wall (Fig. 1: 3, 4). Thus, the study of the seed characters in seven taxa of predominantly Far-Eastern Rhodiola section Rhodiola representatives revealed a heterogeneous seed surface sculpturing in the section. This observation contradicts data on other Crassulaceae genera, Hylotelephium, Aizopsis and Orostachys, traditionally considered to be closely related to Rhodiola. It has been shown that their intrageneric taxa (section and series) are characterized by a uniform set of seed coat features that could be used to discriminate these entities (Gontcharova, 1999; Hart and Berendsen, 1980; Knapp, 1994; Koldaeva and Gontcharova, 2005). Here we report three distinct types of seed coat sculpture, each characterized by a specific anatomical structure (Table 1, Figs. 1–4). Variability of this character in the section under discussion is not in agreement with the stability of this character in many other plant genera (Barthlott, 1984; Esau, 1977). This ARTICLE IN PRESS S.B. Gontcharova et al. / Flora 204 (2009) 17–24 Fig. 4. Scanning electron micrographs of Rhodiola seeds. 1–6 tenuicostate surface: Rh. rosea subsp. rosea (1, 2), Rh. integrifolia (3, 4); R. rosea subsp. arctica (5); Rh. rosea subsp. sachalinensis (6). may be indicative of the section’s artificial nature. Indeed, a recent molecular study on phylogenetic relationships in Sedoideae could not define Rhodiola sections and series as monophyletic entities (Mayuzumi and Ohba, 2004). The current taxonomic structure of the genus remains therefore in question, and taxon sampling in both molecular and seed micromorphology analyses is still too limited to draw any conclusion from seed coat features in order to characterize Rhodiola supraspecific taxa. It is also evident that the significance of seed coat sculpturing at the lower taxonomic levels is not uniform in the genus Rhodiola. The characteristics of seed coat sculpturing are nearly identical in the three taxa that are characterized by a colliculate seed surface, Rh. alsia, Rh. angusta and Rh. ishidae, with the consequence of little value in species distinction. The close relationship of these taxa, based on their similar vegetative morphology, has already been stressed by Ohba (1981). The common seed coat sculpture type and the presence of large wing-like projection or folds in all of them emphasize this as well. The common seed coat features are known only in these three species, differentiating them from other members of the sect.
Rhodiola. Therefore, we believe that these species can be grouped into one sect. Algida (Boriss.) S. B. Gontch. (Gontcharova, 2006). In contrast, the morphological characters are more diverse in seeds with a tenuicostate surface type and their patterns clearly differentiate taxa having seeds of this type (e.g. Rh. rosea and its subspecies). Discrimination of these subspecies based on vegetative morphology alone is somewhat problematic, and some specialists reject them as synonyms of the type variety (Ohba, 1981, 2005). However, the data presented here put such a treatment in question. The broad variability of the seed surface sculpture revealed in Rh. rosea subsp. rosea contradicts a generally accepted notion that morphological features of the seed coats show little plasticity and thus the variation among individuals most likely reflects genetic differences (Barthlott, 1981; Buss et al., 2001). Seed coat feature diversity in one taxon (e.g. Rh. rosea subsp. rosea) suggests either ambiguous identification of some specimens or the artificial nature of the taxon. We were unable to establish firm relationships between seed coat morphology and specimen origin. However, plants from Europe have a laticostate seed surface, while those from Asia are observed to have two types of seed coats (laticostate and tenuicostate), with the tenuicostate type the more common. This pattern tentatively suggests that at least some populations of this widely distributed species may, in fact, represent a cryptic subspecies or even taxa of a higher rank otherwise weakly differentiated morphologically from Rh. rosea subsp. rosea. In conclusion, seed morphology and anatomical characters of the Far-Eastern representatives of Rhodiola sect. Rhodiola show considerable variation that may reflect non-monophyly of the section, suggested by molecular phylogeny data. At the species level, the seed coat characters correspond to the features of gross morphology and they identify at least three morphotypes in the section. Diversity of seed coat sculptures in widely distributed Rh. rosea subsp. roseapossibly suggests the presence of cryptic species and requires further studies. Acknowledgement We would like to thank the collectors and curators of the TNS and TI herbaria for allowing us to use their plant material. This study was supported in part by the grant from FEB RAS (06-III-A-06-143). Appendix A. Specimens examined Rhodiola alsia TNS 103785; Rh. angusta TNS 67793; ARTICLE IN PRESS S.B. Gontcharova et al. / Flora 204 (2009) 17–24 23 Rh. arctica obtained from the Polar-Alpine Botanical Garden-Institute of the KSC RAS; Rh. integrifolia Kamchatka, Bistrynskiy Distr., 5 km from village Esso 850 m alt. 29.08.1998, V. V. Yakubov; Kamchatka, Bystrinsky Distr., Sredinniy Ridge, 4 km east from village Esso, 1000 m alt. 01.09.1998, V. V. Yakubov; Kamchatka, Bystrinsky Distr., Sredinniy Ridge, 800 m alt. 30.08.2000, V. V. Yakubov; Central Kamchatka, south slopes of vulcano Tolbachik, 22.08.2000. V. V. Yakubov; TNS 237779; Rh. ishidae Japan, Honshu, Hayachin, 27.09.98. A. A. Gontcharov; TNS 228917; TNS 303356; TNS 12448; TNS 228917; TI 11547; Rh. rosea ssp. rosea Hokkaido, Shiretoko, Mt. Rausu, 02.09.98, A.A. Gontcharov; Kamchatka, Bystrinsky Distr., foots of vulcano Anaun, 820 m alt. 11.09. 1998.V. V. Yakubov; Central Kamchatka, south slopes of vulcano Ostri Tolbachik, 26.08.2000. V. V. Yakubov; Japan, Honshu, Mt. Hayachine, 27.09.98. A. A. Gontcharov, TNS 107450 (USA); TNS 209962 (Switzerland), TNS 198310 (Scotland), TNS 246341 (Japan, Honshu), TNS 603276 (Japan, Hokkaido), TNS 107450 (USA), TI 90800 (Japan, Honshu), TI 908005 (Japan, Honshu), TI 867092 (Japan, Hokkaido), TI 758154 (Japan, Honshu), TI 5511 (Japan, Hokkaido), TI 177 (Japan, Hokkaido, Isl. Rishiri, 09.05. 1978), TI 909004 (Japan, Honshu), TI 758154 (Japan, Honshu), TNS 107450 (USA), TNS 29776; TNS 209962 (Switzerland, Geneva); Rh. rosea ssp. sachalinensis Sakhalin, Isl. Moneron, 10.09.91, M. N. Abankina, S. B. Gontcharova; Sakhalin, Aniva Bay, near village Yuznoe, 28.08.2001, S. B. Gontcharova. References Attar, F., Keshvari, A., Ghahreman, A., Zarre, S., Aghabeigi, F., 2007. Micromorphological studies on Verbascum (Scrophulariaceae) in Iran with emphasis on seed surface, capsule ornamentation and trichomes. Flora 202, 169–175. Barthlott, W., 1981. Epidermal and seed surface characters of plants: systematic applicability and some evolutionary aspects. Nord. J. Bot. 1, 345–354. Barthlott, W., 1984. Microstructural features of seed surfaces. In: Heywood, V.H., Moore, D.M. (Eds.), Current Concepts in Plant Taxonomy. Academic Press, London, pp. 95–105. Buss, C.C., Lammers, T.G., Wise, R.R., 2001. Seed coat morphology and its systematic implications in Cyanea and other genera of Lobelioideae (Campanulaceae). Am. J. Bot. 88, 1301–1308. Clausen, R., 1984. Sedum (Crassulaceae) of the Mexican Cordilleran Plateau. Gent. Herb. 12, 8–36. Corner, E.J.H., 1976. The Seeds of the Dicotyledons. Cambridge University Press, Cambridge. Esau, K., 1977. Anatomy of Seed Plants, second ed. Wiley, New York.
- Page 1 and 2: Flora 204 (2009) 17-24 Seed surface
- Page 3 and 4: Table 1. Seed morphological charact
- Page 5: Transverse and longitudinal section
22<br />
<strong>Seed</strong>s with nearly smooth cuticle were most common,<br />
<strong>some</strong> specimens had a helical or granular <strong>surface</strong> (Fig. 3:<br />
4; Fig. 4: 2).<br />
Cross-sections clearly show that <strong>the</strong> exotesta anticl<strong>in</strong>al<br />
walls are raised, S-shaped, and <strong>the</strong>y form folds that<br />
appear to thicken, while <strong>the</strong> pericl<strong>in</strong>al wall is concave<br />
and nearly aligned with <strong>the</strong> <strong>in</strong>ner wall (Fig. 1: 3, 4).<br />
Thus, <strong>the</strong> study <strong>of</strong> <strong>the</strong> seed characters <strong>in</strong> seven taxa <strong>of</strong><br />
predom<strong>in</strong>antly Far-Eastern Rhodiola section Rhodiola<br />
<strong>representatives</strong> revealed a heterogeneous seed <strong>surface</strong><br />
sculptur<strong>in</strong>g <strong>in</strong> <strong>the</strong> section. This observation contradicts<br />
data on o<strong>the</strong>r Crassulaceae genera, Hylotelephium,<br />
Aizopsis and Orostachys, traditionally considered to be<br />
closely related to Rhodiola. It has been shown that <strong>the</strong>ir<br />
<strong>in</strong>trageneric taxa (section and series) are characterized<br />
by a uniform set <strong>of</strong> seed coat features that could be used<br />
to discrim<strong>in</strong>ate <strong>the</strong>se entities (Gontcharova, 1999; Hart<br />
and Berendsen, 1980; Knapp, 1994; Koldaeva and<br />
Gontcharova, 2005). Here we report three dist<strong>in</strong>ct types<br />
<strong>of</strong> seed coat sculpture, each characterized by a specific<br />
anatomical structure (Table 1, Figs. 1–4). Variability <strong>of</strong><br />
this character <strong>in</strong> <strong>the</strong> section under discussion is not <strong>in</strong><br />
agreement with <strong>the</strong> stability <strong>of</strong> this character <strong>in</strong> many<br />
o<strong>the</strong>r plant genera (Barthlott, 1984; Esau, 1977). This<br />
ARTICLE IN PRESS<br />
S.B. Gontcharova et al. / Flora 204 (2009) 17–24<br />
Fig. 4. Scann<strong>in</strong>g electron micrographs <strong>of</strong> Rhodiola seeds. 1–6 tenuicostate <strong>surface</strong>: Rh. rosea subsp. rosea (1, 2), Rh. <strong>in</strong>tegrifolia<br />
(3, 4); R. rosea subsp. arctica (5); Rh. rosea subsp. sachal<strong>in</strong>ensis (6).<br />
may be <strong>in</strong>dicative <strong>of</strong> <strong>the</strong> section’s artificial nature.<br />
Indeed, a recent molecular study on phylogenetic<br />
relationships <strong>in</strong> Sedoideae could not def<strong>in</strong>e Rhodiola<br />
sections and series as monophyletic entities (Mayuzumi<br />
and Ohba, 2004). The current taxonomic structure <strong>of</strong><br />
<strong>the</strong> genus rema<strong>in</strong>s <strong>the</strong>refore <strong>in</strong> question, and taxon<br />
sampl<strong>in</strong>g <strong>in</strong> both molecular and seed micro<strong>morphology</strong><br />
analyses is still too limited to draw any conclusion from<br />
seed coat features <strong>in</strong> order to characterize Rhodiola<br />
supraspecific taxa.<br />
It is also evident that <strong>the</strong> significance <strong>of</strong> seed coat<br />
sculptur<strong>in</strong>g at <strong>the</strong> lower taxonomic levels is not uniform<br />
<strong>in</strong> <strong>the</strong> genus Rhodiola. The characteristics <strong>of</strong> seed coat<br />
sculptur<strong>in</strong>g are nearly identical <strong>in</strong> <strong>the</strong> three taxa that are<br />
characterized by a colliculate seed <strong>surface</strong>, Rh. alsia,<br />
Rh. angusta and Rh. ishidae, with <strong>the</strong> consequence <strong>of</strong><br />
little value <strong>in</strong> species dist<strong>in</strong>ction. The close relationship<br />
<strong>of</strong> <strong>the</strong>se taxa, based on <strong>the</strong>ir similar vegetative<br />
<strong>morphology</strong>, has already been stressed by Ohba<br />
(1981). The common seed coat sculpture type and <strong>the</strong><br />
presence <strong>of</strong> large w<strong>in</strong>g-like projection or folds <strong>in</strong> all <strong>of</strong><br />
<strong>the</strong>m emphasize this as well. The common seed coat<br />
features are known only <strong>in</strong> <strong>the</strong>se three species, differentiat<strong>in</strong>g<br />
<strong>the</strong>m from o<strong>the</strong>r members <strong>of</strong> <strong>the</strong> sect.


