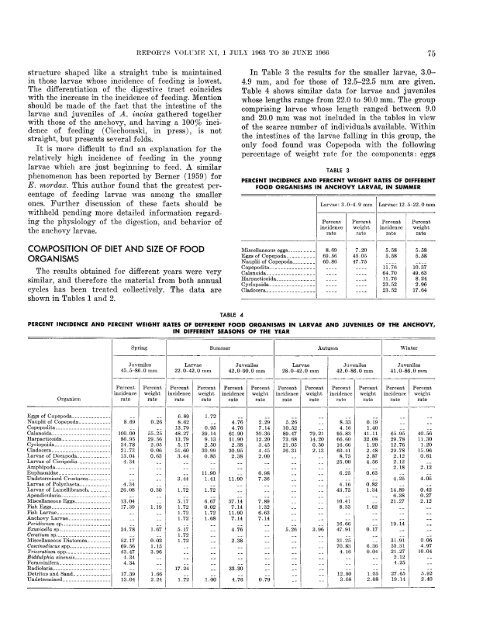
74 CAU,lE’ORSIS COOPERATIVE OCFLWIC FISHERIES ISYESTIGATIOSSof some species of marine fishes, including those ofEngradis nzordax, have an extensive layer of mucouscells over the surfaces of the back of the mouth. Morrisbelieves that “. . . the mucosa might serve as a mechanismfor collecting important quantities of dispersedorganic matter. . .”. Nothing concrete can besaid at this time concerning these assumptions.Other authors, such as Schumann (1965) take intoaccount the diurnal feeding habits of these species.The diurnal habits of the feeding of fishes and especiallyof that of their larvae and juveniles have beenpointed out by several authors : Schumanii (1965) forXardinops caerulca. Erccgovic (1962) for Clupea pilchardus,Duka (1961) for Engradis encrasiclLolus,etc. No matter what differences may exist among theobservations of these various authors, most of themagree that the larvae of these fishes feed exclusivelyduring daylight. They note that during the day,there are times at which the ingestion of food is moreintensive, but no ingestion of food has been observedto take place during the night.In the case of the Argentine anchovy, some assumptionscan be made in this connection, taking into accountall of the points of view mentioned and theauthor’s own observations. The hypothesis of the effectsof net selectivity upon the larvae during theircollection was rejected, since the larvae with the lowestincidence of feeding were gathered in great numbersby means of a large net of purse-seine type with whichthe loss of the larvae was highly improbable. Furthermore,the larvae collected were in very good conditionand did not appear to have starved. Plankton samplescollected simultaneously showed the presence ofnumerous organisms which could be used as food bythe larvae.Nothing can be said here on the diurnal feedinghabits of the anchovy because it was impossible toobtain samples during the night or during twilighthours. All of the material was collected during daylightand generally at about noon. Therefore, if it isassumed that the anchovy behaves in this respect likeother species of marine fishes which feed exclusivelyduring daylight, the fact of finding larvae witli theirintestinal tract devoid of food can not be accountedfor by the diurnal feeding habits.No studies have been made, up to the present time,which could cast light on the problem of the digestionof food by the larvae of E. anchoita. As a consequenceof this, the present analysis is based mainly on comparisonswith other related species, for which a betterunderstanding of these processes has already beenattained. Duka (1961) has reported that the speed ofdigestion in larvae of 6-7 mm in length of E. encrasiclzolusof the Black Sea, at a temperature of23” C., is from 2-2.5 hours. But, on the other hand,according to Schumann (1965) the speed of digestionfor nauplii of Artenzia salina by larrae of E. mordaxof the Pacific, off <strong>California</strong>, is much greater. Thisauthor has observed that “. . . an average of 25 secondsis required by larvae of 15 mm in length orlarger to pass an Artenaia nauplius from the mouth toapproximately one-half of the length of the digestivetract. Progress of a food particle (Artemia) throughthe remainder of the gut is much slower, with anaverage of 2 minutes required for food to reach theend of the gut and form a food plug.” It appearsthen, that a very short time interval is required foran ingested particle to disappear from the intestinaltract of the larvae of E. rnordax.-2 rnme2aFIGURE 1. Development of the digestive tract of the anchovy larvae.p-pyloric appendices, d-ductus pneumaticus. The drawings correspondto the following lengths of fish: a-9.2 mm, b--15.0 mm, c-33.0 mm, d-38.0 mm, e 1 and e 2-50.0 mm.Figure 1 shows the development process of the digestivetract of E. anchoita.As is shown in the drawing, the digestive tract ofthe larvae, until they reach a length of at least 33 mm,is a short and completely straight tube, differentiatedonly in the anterior and posterior parts. In the larvaeof 33 mm length a sketch of the stomach and rudimentsof the pyloric appendices appears. At a lengthof 38 mm the stomach and pyloric appendices showthemselves in a rather developed state and the pigmentof the anterior part of the intestine begins toappear. In the juveniles of 50 mm, the part correspondingto the stomach is well developed and thewhole digestive tract resembles that of an adult individual.This type of digestive tract structure, in theshape of a straight tube, is typical of the engraulidsand clupeids. The very straightness and shortness ofthe digestive tract may favor a rapid digestion andexcretion. It can also be assumed that at the momentthe anchovy larvae are caught, an artificial accelerationof the excretion processes or the vomiting of theintestinal content could take place. By observing thedrawings (Figure 1) it can be seen that the intestinal
ateREPORTS T-OLTJIT", SI, 1 JULl 1963 TO 30 JUKE 1966 75structure shaped like a straight tube is maintainedin those larvae whose incidence of feeding is lowest.The differentiation of the digestive tract coincideswith the increase in the incidence of feeding. Mentionshould be made of the fact that the intestine of thelarvae and juveniles of A. incisa gathered togetherwith those of the anchovy, and having a 100% incidenceof feeding (Ciechomski, in press), is notstraight, but presents several folds.It is more difficult to find an explanation for therelatively high incidence of feeding in the younglarvae which are just beginning to feed. A similarphenomenon has been reported by Berner (1959) forE. mordax. This author found that the greatest percentageof feeding larvae was among the smallerones. Further discussion of these facts should bewithheld pending more detailed information regardingthe physiology of the digestion, and behavior ofthe anchovy larvae.PercentincidenceratePercentweightrate ~Percentincidence--PercentweightrateCOMPOSITION OF DIET AND SIZE OF FOODORGAN ISMSMiscellaneous eggs ._..__ 8.69Eggs of Copepoda .____.____ 69.56Nauplii of Copepoda _..__ - __60.86Copepodita- --- - -------------.The results obtained for different years were very Cdanoida .....................similar, and therefore the material from both annualHarpacticoida-------------- ----C yclopoida- ---- - - ---- - ----- - --cycles has been treated collectively. The data are Cladocera .....................____7.2045.0547.755.585.58....<strong>11</strong>.7664.70<strong>11</strong>.7623.52_.-- 23.525.585.58._..10.3749.638.242.9617.64PERCENT INCIDENCE AND PERCENT WEIGHT RATES OF DIFFERENT FOOD ORGANISMS IN LARVAE AND JUVENILES OF THE ANCHOVY,IN DIFFERENT SEASONS OF THE YEARI Spring SummerAutumn WinterOrganismJuveniles45.5-86.0 mmIPercentincidenceratePercentweightrateEggs of Copepoda .............. ..Nauplii of Copepoda ._.____._..8.69 0.26Copepodita.. ..................Calanoida- ......................100.00_.55.25Harpacticoida ................. 86.95Cyclopoida- ................... 34.7829.562.05Cladocera . 21.73 0.06Larvae of Decapoda .____.____..13.04 0.63Larvae of Cirripedia- _______._. . 4.34__Bmphipoda.-. .................Euphausidae ......................._..Undetermined Crustacea.- ._. -. .. _.Larvae of Polychaeta.- ._______4.34 ._26.08_.13.0417.39Larvae of Lamellibranch. .......A4pendicularia.. ................Miscellaneous Eggs. ............Fish Eggs .Fish Larvae ...................Anchovy Larvae. ..............Peridinium sp .Ezuuiaella sp .Ceratium sp .Miscellaneous Diatomea.. ......Coscinodiscus spp.- .............Triceratium spp .Biddulphia sinenszs.. ...........Foraminifera- .................Radiolaria .Detritus and Sand .............Undetermined ................._.._..34.7852.i;69.5643.474.344.34._17.3913.04__0.30....1.19._.._.1.67._0.021.153.96..__..1.662.24Larvae22.0-42.0 mmPercentncidencerate6.898.6213.7948.2713.795.1751.603.44.._.__3.441.G..5.171.721.721.72_.5.171.721.72.____-._17.24..1.72Percentweightrate1.72__0.9539.149.132.5030.990.85.._.<strong>11</strong>.901.41--1.72__6.670.021.721.68___.._.______---__1.00Juveniles42.0-90.0 mmPercentncidencarate4.764.7661.90<strong>11</strong>.902.3830.952.38____..<strong>11</strong>.90_.__57.G7.14<strong>11</strong>.907.144.76..2.38_____-33.30..4.76____Percentweightrate__2.297.1430.3612.203.454.452.00___.6.987.36__.___7.891.326.637.14___...._.____..__...0.79Larvae28.0-42.0 mmPercentncidenceratePercentweightrate__....79.2<strong>11</strong>4.200.502.13..._..__._._._._....__._..3.96__.....___.___....Juveniles42.0-86.0 mm__-__Percentncidencerate._8.334.1695.8366.6016.6660.418.7525.00..6.25..4.1643.75_ _10.418.33.__.16.6647.91..31.2570.834.16._._..12.503.08Percentweightrate__0.191.4041.<strong>11</strong>32.081.202.482.874.56._0.63..0.821.34_-..1.62__.___0.17___.6.360.04__.._.1.052.08Juveniles41.0-86.0 mmPercentincidencerate_ _._..65.9529.7812.7629.782.122.122.18._4.25._14.896.3821.27_..___19.14__..31.9155.3121.272.124.25._27.6519.14Percentweightrate.-__._40.56<strong>11</strong>.301.2015.960.61-_2.12..4.05_-0.420.272.12____----._0.064.9710.04.___._5.922.40
- Page 1 and 2:
STATE OF CALIFORNIAMARINE RESEARCH
- Page 3 and 4:
STATE OF CALIFORNIADEPARTMENT OF FI
- Page 5 and 6:
RONALD REAGAXGovcriwr of the Slate
- Page 7 and 8:
PART 1REVIEW OF ACTIVITIESJuly 1,19
- Page 9 and 10:
REPORTS YOLUAIE SI, 1 JULY 1963 TO
- Page 11 and 12:
REPORTS VOLUNE XI, 1 JULY 1963 TO 3
- Page 13 and 14:
REPORTS YOLUME SI, 1 JULY 1963 TO 3
- Page 15 and 16:
REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 17 and 18:
REPORTS vor,uiwIi; SI, 1 JULY 1063
- Page 19 and 20:
REPORTS TOT2T71\IF: SI, 1 JUJIT 196
- Page 21 and 22:
IiEPOHTS TOLUME XI, 1 JULY 1963 TO
- Page 23 and 24:
REVIEW OF THE PELAGIC WET FISHERIES
- Page 25 and 26: KEI'OiiTH TOLUJIE SI, 1 JULY 1963 T
- Page 27 and 28: REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 29 and 30: PART IISYMPOSIUM ON ANCHOVIES, GENU
- Page 31 and 32: OCEANIC ENVIRONMENTS OF THE GENUS E
- Page 33 and 34: KEI'OKTS VOLIXI.: SI, 1 JULY 1963 T
- Page 35 and 36: REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 37 and 38: REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 39 and 40: REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 41 and 42: ~~~REPORTS VOLUME XI, 1 JULY 1963 T
- Page 43 and 44: REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 45 and 46: REPORTS VOLUME SI, 1 JULY 1963 TO 3
- Page 47 and 48: 130'E 132' 134' 136' 138' 140' 142-
- Page 49 and 50: REPORTS VOLUSIE SI, 1 JULY 1963 TO
- Page 51 and 52: REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 53 and 54: REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 55 and 56: REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 57 and 58: KI3I’ORTH VOLUJIE XI, 1 JULY 1963
- Page 59 and 60: REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 61 and 62: REPORTS VOLUME SI, 1 JULY 1963 TO 3
- Page 63 and 64: REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 65 and 66: REPORTS VOLUIIE SI. 1 JULY 1963 TO
- Page 67 and 68: REPORTS T’OLUME XI, 1 JULY 1963 T
- Page 69 and 70: INFLUENCE OF SOME ENVIRONMENTAL FAC
- Page 71 and 72: REPORTS Y‘OLUJIE XI, 1 JULY 1963
- Page 73 and 74: REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 75: ~table shows the values calculated
- Page 79 and 80: REPORTS POLGNE SI, 1 JU LY 1963 TO
- Page 81 and 82: REPORTS VOLUXE XI, 1 JULY 1963 TO 3
- Page 83 and 84: REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 85 and 86: _________--_____REPORTS VOLUJIE SI,
- Page 87 and 88: REPORTS TOLTIJIE SI, 1 JULY 1963 TO
- Page 89 and 90: -20/5-0 10-f 5-w 25-0 20-= 15-: 10-
- Page 91 and 92: REPORTS VOLUNE SI. 1 JULY 19G3 TO 3
- Page 93 and 94: REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 95 and 96: REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 97 and 98: REPORTS VOLURlE SI, 1 JULY 1963 TO
- Page 99 and 100: REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 101 and 102: REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 103 and 104: REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 105 and 106: REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 107 and 108: THE PREDATION OF GUANO BIRDS ON THE
- Page 109 and 110: iod and after 8 AM during the rest
- Page 111 and 112: REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 113 and 114: REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 115 and 116: FISH E RYThe California anchovy fis
- Page 117 and 118: ~~ ::REPORTS VOLUME XI, 1 JULY 1963
- Page 119: CO-OCCURRENCES OF SARDINE AND ANCHO
- Page 122 and 123: 120 CALIFORNIA COOPERATIVE OCEANIC
- Page 124 and 125: CALIFORNIA COOPERATIVE OCEANlC FISH
- Page 126 and 127:
124 CALIFORNIA COOPERATIVE OCEANIC
- Page 128 and 129:
126CALIFORNIB COOPERATIVE OCEANIC F
- Page 130 and 131:
128 CALIFORNIA COOPERATIVE OCEANIC
- Page 132 and 133:
~~ ~130 CALIFORNIA COOPERATIVE OCEA
- Page 134 and 135:
132 CALIF0RhTTIA COOPERATIVE OCEANI
- Page 136 and 137:
134 CALIFORNIA COOPERATIVE OCEANIC
- Page 138 and 139:
THE ACCUMULATION OF FISH DEBRIS IN
- Page 140 and 141:
138 CALIFORNIA COOPERATIVE OCEANIC
- Page 143 and 144:
PART IllSCIENTIFIC CONTRIBUTIONS
- Page 145 and 146:
REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 147 and 148:
REPORTS VOLUME SI, 1 JULY 1963 TO 3
- Page 149 and 150:
REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 151 and 152:
REPORTS VOLU&IE XI, 1 JULY 1963 TO
- Page 153 and 154:
REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 155 and 156:
REPORTS VOLUME SI, 1 JULY 1963 TO 3
- Page 157 and 158:
SUMMARY OF THERMAL CONDITIONS AND P
- Page 159 and 160:
SEASONAL VARIATION OF TEMPERATURE A
- Page 161 and 162:
REPORTS VOLUME SI, 1 JULY 1963 TO 3
- Page 163 and 164:
including the semiannual harmonic?
- Page 165 and 166:
REPORTS VOLUME XI, 1 JULY 1983 TO 3
- Page 167 and 168:
REPORTS VOiLUME XI, 1 JULY 1983 TO
- Page 169:
REPORTS VOLUME XI, 1 JULY 1963 TO 3
- Page 172 and 173:
170 CALIFORNIA COOPERATIVE OCWIC FI
- Page 174 and 175:
172 CALIFORNIA COOPERATIVE OCBANIC
- Page 176 and 177:
174 CALIFORNIA COOPERATIVE OCEANIC
- Page 178 and 179:
176 CALIFORNIA COOPERATIVE OCEANIC
- Page 180 and 181:
178 CALIFORNIA COOPERATIVE OCEANIC
- Page 182 and 183:
180 CALIFORNIA COOPERATIVE OCEANIC
- Page 184 and 185:
182 CALIFORNIA COOPERATIVE OCEANIC
- Page 186 and 187:
184CALIFORNIA COOPERATIVE OCEANIC F
- Page 188 and 189:
186 CALIFORNIA COOPERATIVE OCEANIC
- Page 190:
CONTENTSI. Review of Activities Pag