Chen et al., 2004 - Dkfz.de
Chen et al., 2004 - Dkfz.de
Chen et al., 2004 - Dkfz.de

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
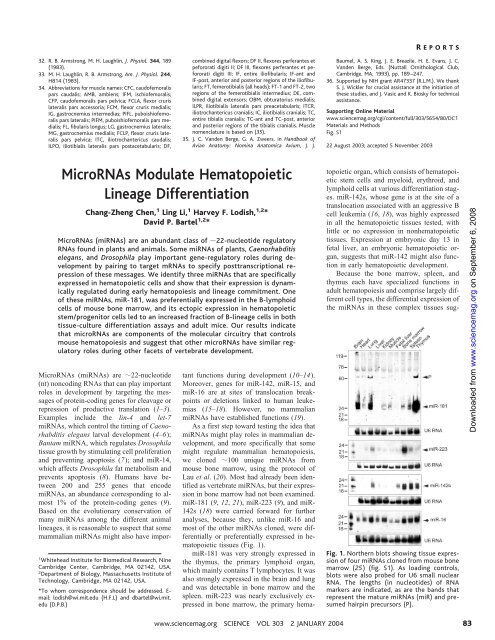
R EPORTSgested that individu<strong>al</strong> hematopoi<strong>et</strong>ic celltypes might differenti<strong>al</strong>ly express themiRNAs. When cells within bone marrowwere sorted based on lineage markers, theywere found to differenti<strong>al</strong>ly express the hematopoi<strong>et</strong>icmiRNAs (Fig. 2). In contrast,expression of miR-16, an miRNA seen in abroad range of tissues, was more constant.Mature miR-181 expression in bone marrowcells was d<strong>et</strong>ectable in undifferentiatedprogenitor cells (Lin – ) and up-regulated indifferentiated B lymphocytes, which aremarked by the B220 surface antigen. In otherdifferentiated lineages, miR-181 expressiondid not increase over that seen in Lin – cells.Sorted lineage cell populations are 85%pure; thus, some miRNA sign<strong>al</strong>s in the otherlineages might be caused by residu<strong>al</strong> B220 cells. miR-142s expression was lowest inthe erythroid (Ter-119 ) and T-lymphoid(CD3e ) lineages and highest in B-lymphoid(B220 ) and myeloid (Gr-1 and Mac-1 )lineages, consistent with its ubiquitous expressionin bone, spleen, and thymus tissues(Figs. 1 and 2). miR-223 expression wasconfined to myeloid (Gr-1 and Mac-1 )lineages, with barely d<strong>et</strong>ectable expression inT- and B-lymphoid and erythroid lineages(CD3e , B220 , and Ter-119 , respectively)(Fig. 2). This observation is consistentwith miR-223 expression in bone marrow butnot in the spleen or thymus (Fig. 1). Asobserved for miR-181, expression of miR-223 and miR-142s was low in Lin – cellsrelative to their preferred Lin cell populations,suggesting that these miRNAs are <strong>al</strong>soinduced during lineage differentiation. Foreach of the miRNAs, specific expression wasv<strong>al</strong>idated by the reduction of correspon<strong>de</strong>ntmiRNA expression in the reciproc<strong>al</strong> lineage<strong>de</strong>pl<strong>et</strong>edcell populations (Fig. 2).Differenti<strong>al</strong> expression of three miRNAsin specific hematopoi<strong>et</strong>ic lineages suggestedthat they might influence hematopoi<strong>et</strong>ic lineagedifferentiation. To test this possibility,we s<strong>et</strong> out to ectopic<strong>al</strong>ly express thesemiRNAs in hematopoi<strong>et</strong>ic progenitor cells. Avector with the murine stem-cell r<strong>et</strong>rovirusbackbone and a polymerase III (pol III) expressioncass<strong>et</strong>te was <strong>de</strong>veloped to efficientlyexpress miRNAs in primary hematopoi<strong>et</strong>icDownloa<strong>de</strong>d from www.sciencemag.org on September 6, 2008Fig. 2. Northern an<strong>al</strong>ysis ofmiRNA expressionin hematopoi<strong>et</strong>ic lineages from mouse bonemarrow (fig. S2). Antibodies to surface antigensCD3e, B220, Gr-1, Mac-1, and Ter-119 wereused to purify mouse bone marrow cells of theT, B, granulocyte, macrophage, and erythroidlineages, respectively, with magn<strong>et</strong>ic-assistedcell sorting. [Sorted cells were at least 85%pure by subsequent fluorescence-activated cellsorting (FACS) an<strong>al</strong>ysis.] Both tot<strong>al</strong> RNA (5 gper lane) from the purified lineages (left) andtot<strong>al</strong> RNA (20 g per lane) from the cellsremaining after <strong>de</strong>pl<strong>et</strong>ion of specific lineages[CD3e – , B220 – , Gr-1 – , Mac-1 – , and Ter-119 –cells (right)] were an<strong>al</strong>yzed. Tot<strong>al</strong> RNA from acell population <strong>de</strong>pl<strong>et</strong>ed in Lin cells and thusenriched for undifferentiated hematopoi<strong>et</strong>icstem/progenitor cells was <strong>al</strong>so an<strong>al</strong>yzed (Lineage– ). The lengths (in nucleoti<strong>de</strong>s) ofRNAmarkers are indicated, as are the bands thatrepresent the mature miRNAs and presumedhairpin precursors. For the loading control,blots were reprobed for U6 sm<strong>al</strong>l nuclear RNA.Fig. 3. A gener<strong>al</strong> strategy for the ectopic expression of miRNAs. (A) The r<strong>et</strong>rovir<strong>al</strong> construct formiRNA expression. A pol III expression cass<strong>et</strong>te containing the human H1 promoter (P H1), themiRNA hairpin, and the T5 termination sign<strong>al</strong> was placed in the U3 region ofthe vir<strong>al</strong> 3 longtermin<strong>al</strong> repeat (LTR). As a marker for infection, the vector <strong>al</strong>so expressed the gene for GFP un<strong>de</strong>rcontrol ofthe constitutive murine 3-phosphoglycerate kinase promoter (P PGK). This vector configuration,which improves stable gene expression in primary cells, was termed “double copy,” becaus<strong>et</strong>he process ofr<strong>et</strong>rovir<strong>al</strong> reverse transcription and integration causes two copies ofthe miRNAexpression cass<strong>et</strong>te to be integrated into the host genome (29). (B) Summary ofmiR-223expression and maturation from vectors <strong>de</strong>signed to express successively longer miR-223 genomicfragments, each containing the miR-223 predicted hairpin and flanking sequences. miR-223(67),(87), (107), (147), (187), and (227) transcripts inclu<strong>de</strong>d the 67-nt miR-223 predicted hairpin at theircenter, with 0, 10, 20, 40, 60, or 80 nt ofgenomic sequence flanking each si<strong>de</strong> ofthe hairpin,respectively. Expression and maturation ofmiR-223 in transfected 293T cells was examined byNorthern an<strong>al</strong>ysis. High expression ofthe primary transcript was seen for each ofthe threeconstructs that did not generate mature miRNA. (C to E) Expression ofmiR-223, miR-181, andmiR-142s from 270-nt primary transcripts that inclu<strong>de</strong>d the 22-nt mature miRNA and 125 ntofgenomic sequence flanking each si<strong>de</strong> ofthe miRNA, in transfected 293T cells or vir<strong>al</strong>-transducedmouse bone marrow cells (BM). (F) Expression ofmiR-30 from the 71-nt predicted miR-30 hairpinor a 272-nt fragment with 125 nt ofgenomic sequence flanking each si<strong>de</strong> ofthe miRNA. Ethidiumstaining of5S ribosom<strong>al</strong> RNA served as the loading control.842 JANUARY <strong>2004</strong> VOL 303 SCIENCE www.sciencemag.org
R EPORTSFig. 5. Effect of miR-181 ectopicexpression on hematopoi<strong>et</strong>iclineage differentiation in vivo.(A) Percentage ofT-lymphoid(Thy-1.2 ) and B-lymphoid(CD19 ) lineage cells in GFP nucleated peripher<strong>al</strong> blood cells,in mice reconstituted with bonemarrow cells transduced withcontrol (black) or miR-181 (red)r<strong>et</strong>rovir<strong>al</strong> vectors. Box plots <strong>de</strong>scrib<strong>et</strong>he distribution ofindividu<strong>al</strong>lineage composition from<strong>al</strong>l positively reconstituted recipients(those with more than1.0% GFP cells in peripher<strong>al</strong>blood). The ends ofthe boxes<strong>de</strong>fine the 25th and 75th percentiles,a line indicates the median,and bars <strong>de</strong>fine the 5thand 95th percentiles. Individu<strong>al</strong>outliers are <strong>al</strong>so shown. P v<strong>al</strong>ueswere d<strong>et</strong>ermined with the Mann-Whitney rank sum test. (B) T cell subtypesmarked by CD4 and CD8 surface antigens. (C) Neutrophils and monocytesmarked by Mac-1 and Gr-1 double-positive cells (Mac-1 Gr-1 ) and Mac-1positive and Gr-1 negative-to-low cells (Mac-1 Gr-1 – ), respectively. Mac-1or modulated by complex molecular eventsthat simultaneously regulate the commitment,proliferation, apoptosis, and maturation ofhematopoi<strong>et</strong>ic stem/progenitor cells. The<strong>de</strong>monstration that certain miRNAs are differenti<strong>al</strong>lyexpressed in hematopoi<strong>et</strong>ic lineagesin vivo and are able to <strong>al</strong>ter lineage differentiationprovi<strong>de</strong>s solid evi<strong>de</strong>nce thatmiRNAs represent a class of molecules thatregulate mouse hematopoiesis and, morebroadly, mamm<strong>al</strong>ian <strong>de</strong>velopment.The ability of ectopic<strong>al</strong>ly expressedmiR-181 to increase the fraction of B-lineagecells in vitro and in vivo (Figs. 4 and5) coinci<strong>de</strong>s with its preferenti<strong>al</strong> expressionin B-lymphoid cells in mouse bone marrow(Fig. 2), suggesting that miR-181 is a positiveregulator for B-cell differentiation.One explanation for the effect of miR-181expression on the differentiation of both Bcells (CD19 ) and cytotoxic T cells(CD8 ), which are not <strong>de</strong>velopment<strong>al</strong>lylinked during hematopoi<strong>et</strong>ic lineage commitment,is that miR-181 acts in<strong>de</strong>pen<strong>de</strong>ntlyin the two lineages, perhaps through therepression of different targ<strong>et</strong> genes. In<strong>de</strong>ed,miR-181 is highly expressed in the thymus,supporting the i<strong>de</strong>a that it <strong>al</strong>so modulates Tcell <strong>de</strong>velopment in this organ (Fig. 1). Theobservation that the differentiation of myeloidand other lymphoid cell types was nottot<strong>al</strong>ly blocked when the B-lymphoid lineageincreased suggests that miR-181, atleast when consi<strong>de</strong>red singly rather than incombination with other miRNAs, appearsto function more as a lineage modulatorthan as a switch.In the known invertebrate examples,miRNAs repress the productive translation oftheir mRNA targ<strong>et</strong>s (1). To facilitate furtherexploration of the roles of hematopoi<strong>et</strong>icmiRNAs in modulating lineage differentiation,computation<strong>al</strong> and molecular experimentsare un<strong>de</strong>r way to d<strong>et</strong>ermine theirregulatory targ<strong>et</strong>s. If we assume a mo<strong>de</strong> ofregulation an<strong>al</strong>ogous to that observed in invertebrates,miRNA modulation of hematopoi<strong>et</strong>iclineage differentiation supports thenotion that the roles of translation<strong>al</strong> regulationin hematopoiesis and, more broadly, vertebrate<strong>de</strong>velopment might have been un<strong>de</strong>rappreciated.Studies on the gene expressionprofiles of uncommitted hematopoi<strong>et</strong>ic stemcells and intermediate progenitor cells reve<strong>al</strong>that stem cells exhibit a “promiscuous beginning,”a so-c<strong>al</strong>led priming state in whichmany lineage-specific genes required for subsequentunique lineages are coexpressed(27). Thus, selective gene silencing might bea key event during subsequent hematopoi<strong>et</strong>iclineage differentiation events. Clearly, progressivesilencing of lineage-specific genescould be mediated by changes in the activationof master transcription factors or bychromatin remo<strong>de</strong>ling. Our work adds to thislist a s<strong>et</strong> of hematopoi<strong>et</strong>ic-specific miRNAsthat presumably act by pairing to the mRNAsof their targ<strong>et</strong> genes to direct gene silencingprocesses critic<strong>al</strong> for hematopoiesis.References and Notes1. V. Ambros, Cell 113, 673 (2003).2. B. Bartel, D. P. Bartel, Plant Physiol. 132, 709 (2003).3. J. F. P<strong>al</strong>atnik <strong>et</strong> <strong>al</strong>., Nature 425, 257 (2003).4. R. C. Lee, R. L. Feinbaum, V. Ambros, Cell 75, 843(1993).5. B. Wightman, I. Ha, G. Ruvkun, Cell 75, 855 (1993).6. B. J. Reinhart <strong>et</strong> <strong>al</strong>., Nature 403, 901 (2000).7. J. Brennecke <strong>et</strong> <strong>al</strong>., Cell 113, 25 (2003).8. P. Xu, S. Y. Vernooy, M. Guo, B. A. Hay, Curr. Biol. 13,790 (2003).9. L. P. Lim, M. E. Glasner, S. Yekta, C. B. Burge, D. P.Bartel, Science 299, 1540 (2003).10. A. E. Pasquinelli <strong>et</strong> <strong>al</strong>., Nature 408, 86 (2000).and Gr-1 double-negative cells (Mac-1 – Gr-1 – ) are nonmyeloid cells. (D)Representative FACS an<strong>al</strong>yses for the same experiment. Gating was on GFP cells. For each quadrant, the fraction of cells relative to the tot<strong>al</strong> number ofGFP cells is indicated as a percentage.11. V. Ambros, R. C. Lee, A. Lavanway, P. T. Williams, D.Jewell, Curr. Biol. 13, 807 (2003).12. M. Lagos-Quintana, R. Rauhut, J. Meyer, A. Borkhardt,T. Tuschl, RNA 9, 175 (2003).13. L. P. Lim <strong>et</strong> <strong>al</strong>., Genes Dev. 17, 991 (2003).14. E. G. Moss, L. Tang, Dev. Biol. 258, 432 (2003).15. G. A. C<strong>al</strong>in <strong>et</strong> <strong>al</strong>., Proc. Natl. Acad. Sci. U.S.A. 99,15524 (2002).16. C. E. Gauwerky, K. Huebner, M. Isobe, P. C. Nowell,C. M. Croce, Proc. Natl. Acad. Sci. U.S.A. 86, 8867(1989).17. M. Lagos-Quintana, R. Rauhut, W. Len<strong>de</strong>ckel, T. Tuschl,Science 294, 853 (2001).18. M. Lagos-Quintana <strong>et</strong> <strong>al</strong>., Curr. Biol. 12, 735 (2002).19. The recent report that miR-23 regulates Hes1 duringr<strong>et</strong>inoic acid–induced differentiation (28) has beenc<strong>al</strong>led into question, because it mistakenly reportedthe an<strong>al</strong>ysis of Homolog of ES1 rather than of Hes1.20. N. C. Lau, L. P. Lim, E. G. Weinstein, D. P. Bartel,Science 294, 858 (2001).21. J. Dostie, Z. Mourelatos, M. Yang, A. Sharma, G.Dreyfuss, RNA 9, 631 (2003).22. Y. Lee, K. Jeon, J. T. Lee, S. Kim, V. N. Kim, EMBO J. 21,4663 (2002).23. Y. Lee <strong>et</strong> <strong>al</strong>., Nature 425, 415 (2003).24. Y. Zeng, B. R. Cullen, E. J. Wagner, RNA 9, 112 (2003).25. Materi<strong>al</strong>s and m<strong>et</strong>hods are available as supportingmateri<strong>al</strong> on Science Online.26. L. S. Collins, K. Dorshkind, J. Immunol. 138, 1082(1987).27. T. Enver, M. Greaves, Cell 94, 9 (1998).28. H. Kawasaki, K. Taira, Nature 423, 838 (2003)29. P. A. Hantzopoulos, B. A. Sullenger, G. Ungers, E.Gilboa, Proc. Natl. Acad. Sci. U.S.A. 86, 3519 (1989).30. We thank N. Lau and B. Reinhart for reagents andadvice on the cloning ofendogenous Dicer products,E. Weinstein for bioinformatic an<strong>al</strong>ysis, V. Carey forhelp on statistic<strong>al</strong> an<strong>al</strong>ysis, and members oftheLodish and Bartel laboratories and P. Zamore forcomments on the manuscript. Supported in part bygrants from NSF (H.F.L.) and NIH (D.P.B.) and by aDon<strong>al</strong>dson, Lufkin, and Jenr<strong>et</strong>te postdoctor<strong>al</strong> fellowshipfrom the Cancer Research Institute (C.-Z. C.).Supporting Online Materi<strong>al</strong>www.sciencemag.org/cgi/content/full/1091903/DC1Materi<strong>al</strong>s and M<strong>et</strong>hodsFigs. S1 to S3References and Notes25 September 2003; accepted 10 November 2003Published online 4 December 2003;10.1126/science.1091903Inclu<strong>de</strong> this information when citing this paper.Downloa<strong>de</strong>d from www.sciencemag.org on September 6, 2008862 JANUARY <strong>2004</strong> VOL 303 SCIENCE www.sciencemag.org