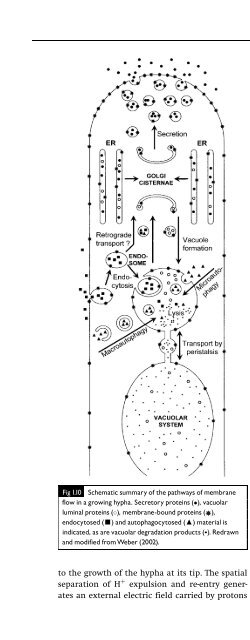
- Page 2:
This page intentionally left blank
- Page 8:
Introduction to Fungi JohnWebster U
- Page 12:
To Philip M. Booth
- Page 18: viii CONTENTS Chapter 6 Chytridiomy
- Page 22: x CONTENTS 15.4 Rhytismataceae 440
- Page 28: Preface to the first edition There
- Page 36: Preface to the third edition Major
- Page 40: Acknowledgements We are indebted to
- Page 46: 2 INTRODUCTION With photosynthetic
- Page 50: 4 INTRODUCTION Fig1.3 Transmission
- Page 54: 6 INTRODUCTION Fig1.5 Structural fo
- Page 58: 8 INTRODUCTION of mushroom-type fru
- Page 62: 10 INTRODUCTION Fig1.8 Diagrammatic
- Page 66: 12 INTRODUCTION Fig1.9 Tubular cont
- Page 72: HYPHAL AGGREGATES 15 leading to the
- Page 76: HYPHAL AGGREGATES 17 Fig1.14 Rhizom
- Page 80: HYPHAL AGGREGATES 19 Fig1.16 Develo
- Page 84: HYPHAL AGGREGATES 21 Germination of
- Page 88: SPORES OF FUNGI 23 over thousands o
- Page 92: SPORES OF FUNGI 25 Fig1.18 Sporangi
- Page 96: SPORES OF FUNGI 27 are unicellular,
- Page 100: SPORES OF FUNGI 29 Fig1.21 Sexual r
- Page 104: SPORES OF FUNGI 31 Fig1.23 Diagrams
- Page 108: TAXONOMY OF FUNGI 33 or the way in
- Page 112: TAXONOMY OF FUNGI 35 closely relate
- Page 116: TAXONOMY OF FUNGI 37 Fig1.26 Phylog
- Page 120:
TAXONOMY OF FUNGI 39 state is apoth
- Page 124:
DICTYOSTELIOMYCETES: DICTYOSTELID S
- Page 128:
DICTYOSTELIOMYCETES: DICTYOSTELID S
- Page 132:
PROTOSTELIOMYCETES: PROTOSTELID PLA
- Page 136:
MYXOMYCETES: TRUE (PLASMODIAL) SLIM
- Page 140:
MYXOMYCETES: TRUE (PLASMODIAL) SLIM
- Page 144:
MYXOMYCETES: TRUE (PLASMODIAL) SLIM
- Page 148:
MYXOMYCETES: TRUE (PLASMODIAL) SLIM
- Page 152:
PLASMODIOPHORALES 55 of unequal len
- Page 156:
PLASMODIOPHORALES 57 Fig 3.2 Club r
- Page 160:
PLASMODIOPHORALES 59 Fig 3.5 Plasmo
- Page 164:
PLASMODIOPHORALES 61 protein was de
- Page 168:
CONTROL OF DISEASES CAUSED BY PLASM
- Page 172:
HAPTOGLOSSA (HAPTOGLOSSALES) 65 alt
- Page 176:
4 Straminipila: minor fungal phyla
- Page 180:
THE STRAMINIPILOUS FLAGELLUM 69 Fig
- Page 184:
LABYRINTHULOMYCOTA 71 phylogenetica
- Page 188:
LABYRINTHULOMYCOTA 73 Fig 4.7 Ultra
- Page 192:
5 Straminipila: Oomycota 5.1 Introd
- Page 196:
INTRODUCTION 77 Fig 5.2 Schematic d
- Page 200:
SAPROLEGNIALES 79 prepared the way
- Page 204:
SAPROLEGNIALES 81 cell wall-softeni
- Page 208:
SAPROLEGNIALES 83 cleavage furrows
- Page 212:
SAPROLEGNIALES 85 Either way, attac
- Page 216:
SAPROLEGNIALES 87 studies on the na
- Page 220:
SAPROLEGNIALES 89 Fig 5.10 Achlya a
- Page 224:
SAPROLEGNIALES 91 there is also evi
- Page 228:
SAPROLEGNIALES 93 Fig 5.13 (a d) Di
- Page 232:
PYTHIALES 95 are keratinophilic, oc
- Page 236:
PYTHIALES 97 Fig 5.15 Pythium mycel
- Page 240:
PYTHIALES 99 Fig 5.17 Pythium middl
- Page 244:
PYTHIALES 101 Fig 5.19 Life cycle o
- Page 248:
PYTHIALES 103 roots severely rotted
- Page 252:
PYTHIALES 105 Fig 5.21 TEM images o
- Page 256:
PYTHIALES 107 Fig 5.23 Schematic dr
- Page 260:
PYTHIALES 109 Fig 5.25 Oogonial dev
- Page 264:
PYTHIALES 111 propagule of Phytopht
- Page 268:
PYTHIALES 113 Fig 5.27 Fungicides a
- Page 272:
PERONOSPORALES 115 followed by para
- Page 276:
PERONOSPORALES 117 Fig 5.28 Peronos
- Page 280:
PERONOSPORALES 119 P. parasitica an
- Page 284:
PERONOSPORALES 121 and in market ga
- Page 288:
PERONOSPORALES 123 Fig 5.32 Albugo
- Page 292:
SCLEROSPORACEAE 125 pass the winter
- Page 296:
6 Chytridiomycota 6.1 Introduction
- Page 300:
INTRODUCTION 129 In the inoperculat
- Page 304:
INTRODUCTION 131 Most zoospores are
- Page 308:
INTRODUCTION 133 Fig 6.5 Sexual rep
- Page 312:
CHYTRIDIALES 135 unlicensed workers
- Page 316:
CHYTRIDIALES 137 is golden brown in
- Page 320:
CHYTRIDIALES 139 in that both summe
- Page 324:
CHYTRIDIALES 141 the diatom Asterio
- Page 328:
CHYTRIDIALES 143 zoospore is uninuc
- Page 332:
SPIZELLOMYCETALES 145 Fig 6.13 Nowa
- Page 336:
SPIZELLOMYCETALES 147 Fig 6.15 Olpi
- Page 340:
SPIZELLOMYCETALES 149 zoospores (Fi
- Page 344:
NEOCALLIMASTIGALES (RUMEN FUNGI) 15
- Page 348:
BLASTOCLADIALES 153 and grooming (L
- Page 352:
BLASTOCLADIALES 155 The zoospore of
- Page 356:
BLASTOCLADIALES 157 Fig 6.20 (a h)
- Page 360:
BLASTOCLADIALES 159 membranes is in
- Page 364:
BLASTOCLADIALES 161 a thick-walled
- Page 368:
MONOBLEPHARIDALES 163 Fig 6.24 Summ
- Page 372:
7 Zygomycota 7.1 Introduction The p
- Page 376:
ZYGOMYCETES: MUCORALES 167 species
- Page 380:
ZYGOMYCETES: MUCORALES 169 mycelium
- Page 384:
ZYGOMYCETES: MUCORALES 171 Fig 7.6
- Page 388:
ZYGOMYCETES: MUCORALES 173 The spor
- Page 392:
ZYGOMYCETES: MUCORALES 175 but is c
- Page 396:
ZYGOMYCETES: MUCORALES 177 Fig 7.10
- Page 400:
ZYGOMYCETES: MUCORALES 179 collecte
- Page 404:
EXAMPLES OF MUCORALES 181 Fig 7.13
- Page 408:
EXAMPLES OF MUCORALES 183 Fig 7.15
- Page 412:
EXAMPLES OF MUCORALES 185 Fig 7.17
- Page 416:
EXAMPLES OF MUCORALES 187 Fig 7.19
- Page 420:
EXAMPLES OF MUCORALES 189 Fig 7.2 2
- Page 424:
EXAMPLES OF MUCORALES 191 Thamnidiu
- Page 428:
EXAMPLES OF MUCORALES 193 distribut
- Page 432:
EXAMPLES OF MUCORALES 195 nodding s
- Page 436:
EXAMPLES OF MUCORALES 197 of the un
- Page 440:
EXAMPLES OF MUCORALES 199 Fig 7.31
- Page 444:
ZOOPAGALES 201 5 families and 20 ge
- Page 448:
ENTOMOPHTHORALES 203 An illustrated
- Page 452:
ENTOMOPHTHORALES 205 (2) Germinatio
- Page 456:
ENTOMOPHTHORALES 207 Basidiobolus m
- Page 460:
ENTOMOPHTHORALES 209 Fig 7.38 Basid
- Page 464:
ENTOMOPHTHORALES 211 layered thicke
- Page 468:
ENTOMOPHTHORALES 213 Fig 7.41 Eryni
- Page 472:
ENTOMOPHTHORALES 215 germination (W
- Page 476:
GLOMALES 217 become infected with E
- Page 480:
GLOMALES 219 Fig 7.45 Furia america
- Page 484:
GLOMALES 221 kind of mycorrhiza are
- Page 488:
TRICHOMYCETES 223 group have a worl
- Page 492:
TRICHOMYCETES 225 Simulium in the p
- Page 496:
VEGETATIVE STRUCTURES 227 Fig 8.1 (
- Page 500:
LIFE CYCLES OF ASCOMYCETES 229 Fig
- Page 504:
CONIDIUM PRODUCTION IN ASCOMYCETES
- Page 508:
CONIDIUM PRODUCTION IN ASCOMYCETES
- Page 512:
CONIDIUM PRODUCTION IN ASCOMYCETES
- Page 516:
DEVELOPMENT OF ASCI 237 These nucle
- Page 520:
DEVELOPMENT OF ASCI 239 appendages
- Page 524:
DEVELOPMENT OF ASCI 241 may converg
- Page 528:
DEVELOPMENT OF ASCI 243 Fig 8.14 As
- Page 532:
TYPESOFFRUITBODY 245 the spores may
- Page 536:
CLASSIFICATION 247 mutants and the
- Page 540:
CLASSIFICATION 249 An outline of cu
- Page 544:
TAPHRINALES 251 9.2 Taphrinales The
- Page 548:
SCHIZOSACCHAROMYCETALES 253 about 5
- Page 552:
SCHIZOSACCHAROMYCETALES 255 importa
- Page 556:
SCHIZOSACCHAROMYCETALES 257 Fig 9.5
- Page 560:
PNEUMOCYSTIS 259 formation of the i
- Page 564:
10 Hemiascomycetes 10.1 Introductio
- Page 568:
SACCHAROMYCES (SACCHAROMYCETACEAE)
- Page 572:
SACCHAROMYCES (SACCHAROMYCETACEAE)
- Page 576:
SACCHAROMYCES (SACCHAROMYCETACEAE)
- Page 580:
SACCHAROMYCES (SACCHAROMYCETACEAE)
- Page 584:
SACCHAROMYCES (SACCHAROMYCETACEAE)
- Page 588:
SACCHAROMYCES (SACCHAROMYCETACEAE)
- Page 592:
SACCHAROMYCES (SACCHAROMYCETACEAE)
- Page 596:
CANDIDA (ANAMORPHIC SACCHAROMYCETAL
- Page 600:
CANDIDA (ANAMORPHIC SACCHAROMYCETAL
- Page 604:
GALACTOMYCES (DIPODASCACEAE) 281 of
- Page 608:
SACCHAROMYCOPSIS (SACCHAROMYCOPSIDA
- Page 612:
11 Plectomycetes 11.1 Introduction
- Page 616:
ASCOSPHAERALES 287 ascocarps, and i
- Page 620:
ONYGENALES 289 infected larvae rath
- Page 624:
ONYGENALES 291 Onygenaceae as human
- Page 628:
ONYGENALES 293 disturb the soil. On
- Page 632:
ONYGENALES 295 Fig11.7 Friesian cat
- Page 636:
EUROTIALES 297 have gymnothecia wit
- Page 640:
EUROTIALES 299 penicilli, a further
- Page 644:
EUROTIALES 301 biseriate species As
- Page 648:
EUROTIALES 303 Cheese production in
- Page 652:
EUROTIALES 305 has similarities to
- Page 656:
EUROTIALES 307 are uncommon, with t
- Page 660:
EUROTIALES 309 Fig11.16 Eurotium re
- Page 664:
EUROTIALES 311 Fig11.18 Conidiophor
- Page 668:
EUROTIALES 313 Fig11.21 Elaphomyces
- Page 672:
12 Hymenoascomycetes: Pyrenomycetes
- Page 676:
SORDARIALES 317 The dark-coloured p
- Page 680:
SORDARIALES 319 vitamins by the sti
- Page 684:
SORDARIALES 321 Fig12.3 Schizotheci
- Page 688:
SORDARIALES 323 degenerates. Cultur
- Page 692:
SORDARIALES 325 Mating type factors
- Page 696:
SORDARIALES 327 kilns and bakeries
- Page 700:
SORDARIALES 329 Fig12.8 Neurospora
- Page 704:
SORDARIALES 331 heterokaryotic (A
- Page 708:
XYLARIALES 333 been placed in sever
- Page 712:
XYLARIALES 335 Daldinia concentrica
- Page 716:
HYPOCREALES 337 Fig12.13 serpens. T
- Page 720:
HYPOCREALES 339 mushroom mycelium.
- Page 724:
HYPOCREALES 341 Fig12.16 Conidiopho
- Page 728:
HYPOCREALES 343 Fig12.18 Canker cau
- Page 732:
HYPOCREALES 345 Fig12.21 Nectria ma
- Page 736:
HYPOCREALES 347 Fig12.22 Fusarium c
- Page 740:
CLAVICIPITALES 349 thick apical cap
- Page 744:
CLAVICIPITALES 351 Fig12.25 The hom
- Page 748:
CLAVICIPITALES 353 and dumb, and, b
- Page 752:
CLAVICIPITALES 355 alkaloid product
- Page 756:
CLAVICIPITALES 357 Fig12.29 Epichlo
- Page 760:
CLAVICIPITALES 359 Fig12.31 Endophy
- Page 764:
CLAVICIPITALES 361 artificially inf
- Page 768:
CLAVICIPITALES 363 conidia. The ass
- Page 772:
OPHIOSTOMATALES 365 Fig12.36 Ophios
- Page 776:
OPHIOSTOMATALES 367 Fig12.37 Asexua
- Page 780:
MICROASCALES 369 Fig12.38 Scopulari
- Page 784:
MICROASCALES 371 Fig12.40 Trichurus
- Page 788:
DIAPORTHALES 373 Aspects of pathoge
- Page 792:
DIAPORTHALES 375 enveloped by hypha
- Page 796:
MAGNAPORTHACEAE 377 cAMP levels wer
- Page 800:
MAGNAPORTHACEAE 379 Fig12.44 The in
- Page 804:
MAGNAPORTHACEAE 381 with a hydropho
- Page 808:
MAGNAPORTHACEAE 383 Fig12.47 (a) Co
- Page 812:
MAGNAPORTHACEAE 385 intracellular s
- Page 816:
GLOMERELLACEAE 387 Fig12.51 Colleto
- Page 820:
GLOMERELLACEAE 389 infection proces
- Page 824:
INTRODUCTION 391 Fig13.1 Summary of
- Page 828:
BLUMERIA GRAMINIS 393 of the Erysip
- Page 832:
BLUMERIA GRAMINIS 395 Fig13.3 An ex
- Page 836:
BLUMERIA GRAMINIS 397 leaf surface.
- Page 840:
BLUMERIA GRAMINIS 399 Fig13.5 Inter
- Page 844:
ERYSIPHE 401 that B. graminis survi
- Page 848:
ERYSIPHE 403 Fig13.9 Summer shoot o
- Page 852:
PHYLLACTINIA AND LEVEILLULA 405 Fig
- Page 856:
PHYLLACTINIA AND LEVEILLULA 407 Fig
- Page 860:
CONTROL OF POWDERY MILDEW DISEASES
- Page 864:
CONTROL OF POWDERY MILDEW DISEASES
- Page 868:
Plate1 Slimemoulds(Myxomycota).(a)W
- Page 872:
Plate 3 Chytridiomycota (a c) and Z
- Page 876:
Plate 5 Pyrenomycetes. (a) Daldinia
- Page 880:
Plate 7 Inoperculate Discomycetes (
- Page 884:
Plate 9 Fruit bodies of Homobasidio
- Page 888:
Plate11 Gasteromycetes (a f) and He
- Page 892:
CONTROL OF POWDERY MILDEW DISEASES
- Page 896:
PYRONEMA (PYRONEMATACEAE) 415 Table
- Page 900:
ALEURIA (PYRONEMATACEAE) 417 simult
- Page 904:
ASCOBOLUS (ASCOBOLACEAE) 419 Aleuri
- Page 908:
ASCOBOLUS (ASCOBOLACEAE) 421 Fig14.
- Page 912:
TUBER (TUBERACEAE) 423 of neighbour
- Page 916:
TUBER (TUBERACEAE) 425 Fig14.7 Tube
- Page 920:
MORCHELLA (MORCHELLACEAE) 427 surro
- Page 924:
15 Hymenoascomycetes: Helotiales (i
- Page 928:
SCLEROTINIACEAE 431 Fig15.1 Sclerot
- Page 932:
SCLEROTINIACEAE 433 Fig15.3 Monilin
- Page 936:
SCLEROTINIACEAE 435 Fig15.4 Non-vol
- Page 940:
SCLEROTINIACEAE 437 Fig15.6 Life cy
- Page 944:
DERMATEACEAE 439 Further, B. cinere
- Page 948:
RHYTISMATACEAE 441 Fig15.8 Infectio
- Page 952:
OTHER REPRESENTATIVES OF THE HELOTI
- Page 956:
OTHER REPRESENTATIVES OF THE HELOTI
- Page 960:
GENERAL ASPECTS OF LICHEN BIOLOGY 4
- Page 964:
Table 16.1. Summary of the most imp
- Page 968:
GENERAL ASPECTS OF LICHEN BIOLOGY 4
- Page 972:
GENERAL ASPECTS OF LICHEN BIOLOGY 4
- Page 976:
LECANORALES 455 listed in Table 16.
- Page 980:
LECANORALES 457 Fig16.8 Cladonia py
- Page 984:
17 Loculoascomycetes 17.1 Introduct
- Page 988:
PLEOSPORALES 461 Pseudoparaphyses a
- Page 992:
PLEOSPORALES 463 Fig17.3 Leptosphae
- Page 996:
PLEOSPORALES 465 Fig17.5 Ascochyta
- Page 1000:
PLEOSPORALES 467 Fig17.8 Epicoccum
- Page 1004:
PLEOSPORALES 469 used (Ellis, 1971b
- Page 1008:
PLEOSPORALES 471 Fig17.11 Alternari
- Page 1012:
PLEOSPORALES 473 Fig17.12 Helmintho
- Page 1016:
PLEOSPORALES 475 Fig17.14 Curvulari
- Page 1020:
PLEOSPORALES 477 1999). Victorin bi
- Page 1024:
PLEOSPORALES 479 Fig17.17 Venturia
- Page 1028:
DOTHIDEALES 481 Table 17.3. Some an
- Page 1032:
DOTHIDEALES 483 Fig17.20 Wheat leaf
- Page 1036:
DOTHIDEALES 485 Fig17.22 Cercospori
- Page 1040:
18 Basidiomycota 18.1 Introduction
- Page 1044:
DEVELOPMENT OF BASIDIA 489 Fig18.2
- Page 1048:
BASIDIOSPORE DEVELOPMENT 491 Fig18.
- Page 1052:
THE MECHANISM OF BASIDIOSPORE DISCH
- Page 1056:
NUMBERS OF BASIDIOSPORES 495 Fig18.
- Page 1060:
497 BASIDIOSPORE GERMINATION AND HY
- Page 1064:
BASIDIOSPORE GERMINATION AND HYPHAL
- Page 1068:
ASEXUAL REPRODUCTION 501 Fig18.12 S
- Page 1072:
ASEXUAL REPRODUCTION 503 Fig18.14 A
- Page 1076:
ASEXUAL REPRODUCTION 505 Fig18.16 S
- Page 1080:
MATING SYSTEMS IN BASIDIOMYCETES 50
- Page 1084:
MATING SYSTEMS IN BASIDIOMYCETES 50
- Page 1088:
RELATIONSHIPS 511 colonized by this
- Page 1092:
CLASSIFICATION 513 These taxonomic
- Page 1096:
INTRODUCTION 515 Fig19.1 Examples o
- Page 1100:
STRUCTURE AND MORPHOGENESIS OF BASI
- Page 1104:
STRUCTURE AND MORPHOGENESIS OF BASI
- Page 1108:
STRUCTURE AND MORPHOGENESIS OF BASI
- Page 1112:
STRUCTURE AND MORPHOGENESIS OF BASI
- Page 1116:
IMPORTANCE OF HOMOBASIDIOMYCETES 52
- Page 1120:
IMPORTANCE OF HOMOBASIDIOMYCETES 52
- Page 1124:
IMPORTANCE OF HOMOBASIDIOMYCETES 52
- Page 1128:
IMPORTANCE OF HOMOBASIDIOMYCETES 53
- Page 1132:
EUAGARICS CLADE 533 Fig19.14 Basidi
- Page 1136:
EUAGARICS CLADE 535 primordium has
- Page 1140:
EUAGARICS CLADE 537 psychromorbidus
- Page 1144:
EUAGARICS CLADE 539 Fig19.15 Second
- Page 1148:
EUAGARICS CLADE 541 (see Figs. 19.1
- Page 1152:
EUAGARICS CLADE 543 Fig19.16 Basidi
- Page 1156:
EUAGARICS CLADE 545 flesh of the fr
- Page 1160:
EUAGARICS CLADE 547 Fig19.18 Basidi
- Page 1164:
EUAGARICS CLADE 549 Fig19.19 Gravit
- Page 1168:
EUAGARICS CLADE 551 Control of witc
- Page 1172:
EUAGARICS CLADE 553 Fig19.20 Basidi
- Page 1176:
BOLETOID CLADE 555 fungus (P. namek
- Page 1180:
BOLETOID CLADE 557 consumption. The
- Page 1184:
BOLETOID CLADE 559 such timbers tha
- Page 1188:
POLYPOROID CLADE 561 Fig19.23 Basid
- Page 1192:
POLYPOROID CLADE 563 Fig19.24 Trame
- Page 1196:
POLYPOROID CLADE 565 distinct compo
- Page 1200:
RUSSULOID CLADE 567 Fig19.26 Basidi
- Page 1204:
RUSSULOID CLADE 569 predominantly b
- Page 1208:
RUSSULOID CLADE 571 Fig19.28 The bo
- Page 1212:
HYMENOCHAETOID CLADE 573 one of a g
- Page 1216:
GOMPHOID PHALLOID CLADE 575 bodies
- Page 1220:
20 Homobasidiomycetes: gasteromycet
- Page 1224:
EVOLUTION AND PHYLOGENY OF GASTEROM
- Page 1228:
GASTEROMYCETES IN THE EUAGARICS CLA
- Page 1232:
GASTEROMYCETES IN THE EUAGARICS CLA
- Page 1236:
GASTEROMYCETES IN THE BOLETOID CLAD
- Page 1240:
GASTEROMYCETES IN THE BOLETOID CLAD
- Page 1244:
GASTEROMYCETES IN THE GOMPHOID PHAL
- Page 1248:
GASTEROMYCETES IN THE GOMPHOID PHAL
- Page 1252:
21 Heterobasidiomycetes 21.1 Introd
- Page 1256:
CERATOBASIDIALES 595 affected (Sneh
- Page 1260:
CERATOBASIDIALES 597 Fig 21.2 Rhizo
- Page 1264:
DACRYMYCETALES 599 Fig 21.4 Dacrymy
- Page 1268:
AURICULARIALES 601 of basidiospores
- Page 1272:
AURICULARIALES 603 Fig 21.7 Life cy
- Page 1276:
TREMELLALES 605 conjugation of comp
- Page 1280:
TREMELLALES 607 Fig 21.12 Life cycl
- Page 1284:
22 Urediniomycetes: Uredinales (rus
- Page 1288:
UREDINALES: THE RUST FUNGI 611 Fig
- Page 1292:
UREDINALES: THE RUST FUNGI 613 Homo
- Page 1296:
UREDINALES: THE RUST FUNGI 615 Fig
- Page 1300:
UREDINALES: THE RUST FUNGI 617 Fig
- Page 1304:
UREDINALES: THE RUST FUNGI 619 form
- Page 1308:
PUCCINIA GRAMINIS, THE CAUSE OF BLA
- Page 1312:
PUCCINIA GRAMINIS, THE CAUSE OF BLA
- Page 1316:
PUCCINIA GRAMINIS, THE CAUSE OF BLA
- Page 1320:
OTHER CEREAL RUSTS 627 race charact
- Page 1324:
PUCCINIA AND UROMYCES 629 Fig 22.13
- Page 1328:
OTHER MEMBERS OF THE PUCCINIACEAE 6
- Page 1332:
OTHER MEMBERS OF THE PUCCINIACEAE 6
- Page 1336:
MELAMPSORACEAE 635 Fig 22.15 Sectio
- Page 1340:
THE ‘TRUE’ SMUT FUNGI (USTILAGI
- Page 1344:
THE ‘TRUE’ SMUT FUNGI (USTILAGI
- Page 1348:
THE ‘TRUE’ SMUT FUNGI (USTILAGI
- Page 1352:
THE ‘TRUE’ SMUT FUNGI (USTILAGI
- Page 1356:
THE ‘TRUE’ SMUT FUNGI (USTILAGI
- Page 1360:
THE ‘TRUE’ SMUT FUNGI (USTILAGI
- Page 1364:
THE ‘TRUE’ SMUT FUNGI (USTILAGI
- Page 1368:
THE ‘TRUE’ SMUT FUNGI (USTILAGI
- Page 1372:
MICROBOTRYALES (UREDINIOMYCETES) 65
- Page 1376:
EXOBASIDIALES (USTILAGINOMYCETES) 6
- Page 1380:
EXOBASIDIALES (USTILAGINOMYCETES) 6
- Page 1384:
INTRODUCTION 659 Fig 24.1 Basidiomy
- Page 1388:
HETEROBASIDIOMYCETE YEASTS 661 Tabl
- Page 1392:
HETEROBASIDIOMYCETE YEASTS 663 Fig
- Page 1396:
HETEROBASIDIOMYCETE YEASTS 665 wide
- Page 1400:
UREDINIOMYCETE YEASTS 667 Fig 24.4
- Page 1404:
UREDINIOMYCETE YEASTS 669 Fig 24.6
- Page 1408:
USTILAGINOMYCETE YEASTS 671 Fig 24.
- Page 1412:
25 Anamorphic fungi (nematophagous
- Page 1416:
NEMATOPHAGOUS FUNGI 675 We owe much
- Page 1420:
NEMATOPHAGOUS FUNGI 677 Fig 25.3 Ar
- Page 1424:
NEMATOPHAGOUS FUNGI 679 Fig 25.5 Da
- Page 1428:
NEMATOPHAGOUS FUNGI 681 Fig 25.7 Ne
- Page 1432:
NEMATOPHAGOUS FUNGI 683 (group 1),
- Page 1436:
AQUATIC HYPHOMYCETES (INGOLDIAN FUN
- Page 1440:
AQUATIC HYPHOMYCETES (INGOLDIAN FUN
- Page 1444:
AQUATIC HYPHOMYCETES (INGOLDIAN FUN
- Page 1448:
AQUATIC HYPHOMYCETES (INGOLDIAN FUN
- Page 1452:
AQUATIC HYPHOMYCETES (INGOLDIAN FUN
- Page 1456:
AQUATIC HYPHOMYCETES (INGOLDIAN FUN
- Page 1460:
AERO-AQUATIC FUNGI 697 Table 25.3.
- Page 1464:
AERO-AQUATIC FUNGI 699 Fig 25.22 Pr
- Page 1468:
AERO-AQUATIC FUNGI 701 means of dis
- Page 1472:
REFERENCES 703 Aist, J. R. & Israel
- Page 1476:
REFERENCES 705 Arnold, D. L., Blake
- Page 1480:
REFERENCES 707 Barr, D. J. S. (1987
- Page 1484:
REFERENCES 709 Beakes, G. W. & Gloc
- Page 1488:
REFERENCES 711 Beuchat, L. R. (1995
- Page 1492:
REFERENCES 713 Bourett, T. M., Czym
- Page 1496:
REFERENCES 715 Brown, J. S., Whan,
- Page 1500:
REFERENCES 717 Callac, P. (1995). B
- Page 1504:
REFERENCES 719 Castlebury, L. A., R
- Page 1508:
REFERENCES 721 Chiu, S. W. & Moore,
- Page 1512:
REFERENCES 723 Cooke, L. R. & Littl
- Page 1516:
REFERENCES 725 Culvenor, C. C., Bec
- Page 1520:
REFERENCES 727 Degousée, N., Gupta
- Page 1524:
REFERENCES 729 Dissing, H. (1986).
- Page 1528:
REFERENCES 731 Edgar, J. A., Frahn,
- Page 1532:
REFERENCES 733 Falk, S. P., Gadoury
- Page 1536:
REFERENCES 735 Foster, S. J. & Fitt
- Page 1540:
REFERENCES 737 Gauger, W. L. (1975)
- Page 1544:
REFERENCES 739 Haptoglossa heteromo
- Page 1548:
REFERENCES 741 Griffith, J. M., Dav
- Page 1552:
REFERENCES 743 Hampson, M. C. (1988
- Page 1556:
REFERENCES 745 Heath, M. C. & Skala
- Page 1560:
REFERENCES 747 Hohl, H. R. & Iselin
- Page 1564:
REFERENCES 749 Huang, B., Li, Z. G.
- Page 1568:
REFERENCES 751 Ingold, C. T. & Zobe
- Page 1572:
REFERENCES 753 Jiang, J., Stephenso
- Page 1576:
REFERENCES 755 Kendrick, B., ed. (1
- Page 1580:
REFERENCES 757 Ko, W. H. (1980). Ho
- Page 1584:
REFERENCES 759 concern: cytological
- Page 1588:
REFERENCES 761 Latunde-Dada, A. O.,
- Page 1592:
REFERENCES 763 Lingappa, B. T. (195
- Page 1596:
REFERENCES 765 Luttrell, E. S. (198
- Page 1600:
REFERENCES 767 Markovich, N. A. & K
- Page 1604:
REFERENCES 769 Menge, J. A. (1984).
- Page 1608:
REFERENCES 771 Molina, R., Trappe,
- Page 1612:
REFERENCES 773 Moss, M. O. & Long,
- Page 1616:
REFERENCES 775 inoperculaten Discom
- Page 1620:
REFERENCES 777 Obermayer, W. & Poel
- Page 1624:
REFERENCES 779 Ott, S., Meier, T. &
- Page 1628:
REFERENCES 781 Percudani, R., Trevi
- Page 1632:
REFERENCES 783 Pommer, E.-H. (1995)
- Page 1636:
REFERENCES 785 Raper, J. R. (1939).
- Page 1640:
REFERENCES 787 Reynolds, D. R. (197
- Page 1644:
REFERENCES 789 Roncal, T., Cordobé
- Page 1648:
REFERENCES 791 Sánchez, C. (2004).
- Page 1652:
REFERENCES 793 K. A. Powell, A. Ren
- Page 1656:
REFERENCES 795 Silliker, M. E., Mon
- Page 1660:
REFERENCES 797 Solla, A. & Gil, L.
- Page 1664:
REFERENCES 799 Stensrud, Ø., Hywel
- Page 1668:
REFERENCES 801 Swann, E. C. & Taylo
- Page 1672:
REFERENCES 803 Thines, E., Weber, R
- Page 1676:
REFERENCES 805 Uchida, W., Matsunag
- Page 1680:
REFERENCES 807 Verstrepen, K. J., D
- Page 1684:
REFERENCES 809 Waters, H., Butler,
- Page 1688:
REFERENCES 811 Weeks, R. J., Padhye
- Page 1692:
REFERENCES 813 Willetts, H. J. & Bu
- Page 1696:
REFERENCES 815 Xu, J.-T. & Mu, C. (
- Page 1700:
Index Page numbers with images are
- Page 1704:
INDEX 819 ascospore-delimiting memb
- Page 1708:
INDEX 821 Candida parapsilosis 227,
- Page 1712:
INDEX 823 Cordyceps capitata 362 Co
- Page 1716:
INDEX 825 Erysiphe polygoni 402 Ery
- Page 1720:
INDEX 827 hemicellulose 528 Hemilei
- Page 1724:
INDEX 829 Leveillula taurica 407 Le
- Page 1728:
INDEX 831 mycosporine-alanine 387 m
- Page 1732:
INDEX 833 Phallus ravenelii 591 Pha
- Page 1736:
INDEX 835 Puccinia coronata 476, 61
- Page 1740:
INDEX 837 scolytid beetles 366 Scop
- Page 1744:
INDEX 839 thallic conidiogenesis 30
- Page 1748:
INDEX 841 yeasts 3; see Archiascomy