

Memoir cover 0.tif - Ohio University College of Osteopathic Medicine
Memoir cover 0.tif - Ohio University College of Osteopathic Medicine
Memoir cover 0.tif - Ohio University College of Osteopathic Medicine

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
SOCIETY OF VERTEBRATE PALEONTOLOGY, MEMOIR 3<br />
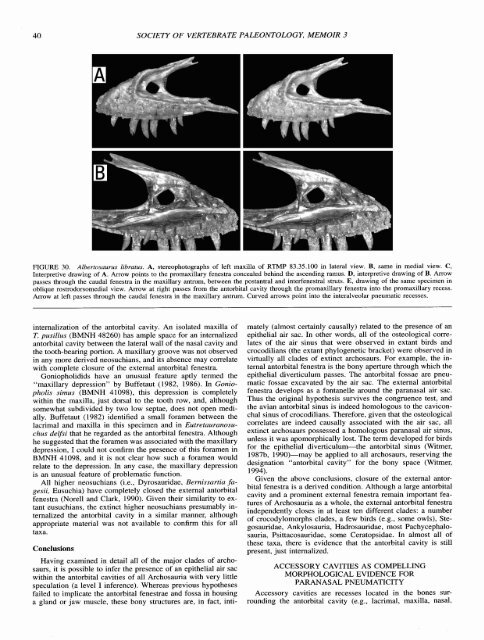
FIGURE 30. Albertosaurus libratus. A, stereophotographs <strong>of</strong> left maxilla <strong>of</strong> RTMP 83.35.100 in lateral view. B, same in medial view. C,<br />
Interpretive drawing <strong>of</strong> A. Arrow points to the promaxillary fenestra concealed behind the ascending ramus. D, interpretive drawing <strong>of</strong> B. Arrow<br />
passes through the caudal fenestra in the maxillary antrum, between the postantral and interfenestral struts. E, drawing <strong>of</strong> the same specimen in<br />
oblique rostrodorsomedial view. Arrow at right passes from the antorbital cavity through the promaxillary fenestra into the promaxillary recess.<br />
Arrow at left passes through the caudal fenestra in the maxillary antrum. Curved arrows point into the interalveolar pneumatic recesses.<br />
internalization <strong>of</strong> the antorbital cavity. An isolated maxilla <strong>of</strong><br />
T. pusillus (BMNH 48260) has ample space for an internalized<br />
antorbital cavity between the lateral wall <strong>of</strong> the nasal cavity and<br />
the tooth-bearing portion. A maxillary groove was not observed<br />
in any more derived neosuchians, and its absence may correlate<br />
with complete closure <strong>of</strong> the external antorbital fenestra.<br />
Goniopholidids have an unusual feature aptly termed the<br />
"maxillary depression" by Buffetaut (1982, 1986). In Goniopholis<br />
sirnus (BMNH 41098), this depression is completely<br />
within the maxilla, just dorsal to the tooth row, and, although<br />
somewhat subdivided by two low septae, does not open medially.<br />
Buffetaut (1982) identified a small foramen between the<br />
lacrimal and maxilla in this specimen and in Eutretauranosuchus<br />
delfsi that he regarded as the antorbital fenestra. Although<br />
he suggested that the foramen was associated with the maxillary<br />
depression, I could not confirm the presence <strong>of</strong> this foramen in<br />
BMNH 41098, and it is not clear how such a foramen would<br />
relate to the depression. In any case, the maxillary depression<br />
is an unusual feature <strong>of</strong> problematic function.<br />
All higher neosuchians (i.e., Dyrosauridae, Bernissartia fagesii,<br />
Eusuchia) have completely closed the external antorbital<br />
fenestra (Norell and Clark, 1990). Given their similarity to extant<br />
eusuchians, the extinct higher neosuchians presumably internalized<br />
the antorbital cavity in a similar manner, although<br />
appropriate material was not available to confirm this for all<br />
taxa.<br />
Conclusions<br />
Having examined in detail all <strong>of</strong> the major clades <strong>of</strong> archosaurs,<br />
it is possible to infer the presence <strong>of</strong> an epithelial air sac<br />
within the antorbital cavities <strong>of</strong> all Archosauria with very little<br />
speculation (a level I inference). Whereas previous hypotheses<br />
failed to implicate the antorbital fenestrae and fossa in housing<br />
a gland or jaw muscle, these bony structures are, in fact, inti-<br />
mately (almost certainly causally) related to the presence <strong>of</strong> an<br />
epithelial air sac. In other words, all <strong>of</strong> the osteological correlates<br />
<strong>of</strong> the air sinus that were observed in extant birds and<br />
crocodilians (the extant phylogenetic bracket) were observed in<br />
virtually all clades <strong>of</strong> extinct archosaurs. For example, the internal<br />
antorbital fenestra is the bony aperture through which the<br />
epithelial diverticulum passes. The antorbital fossae are pneumatic<br />
fossae excavated by the air sac. The external antorbital<br />
fenestra develops as a fontanelle around the paranasal air sac.<br />
Thus the original hypothesis survives the congruence test, and<br />
the avian antorbital sinus is indeed homologous to the caviconchal<br />
sinus <strong>of</strong> crocodilians. Therefore, given that the osteological<br />
correlates are indeed causally associated with the air sac, all<br />
extinct archosaurs possessed a homologous paranasal air sinus,<br />
unless it was apomorphically lost. The term developed for birds<br />
for the epithelial diverticulum-the antorbital sinus (Witmer,<br />
1987b, 1990)-may be applied to all archosaurs, reserving the<br />
designation "antorbital cavity" for the bony space (Witmer,<br />
1994).<br />
Given the above conclusions, closure <strong>of</strong> the external antorbital<br />
fenestra is a derived condition. Although a large antorbital<br />
cavity and a prominent external fenestra remain important features<br />
<strong>of</strong> Archosauria as a whole, the external antorbital fenestra<br />
independently closes in at least ten different clades: a number<br />
<strong>of</strong> crocodylomorphs clades, a few birds (e.g., some owls), Stegosauridae,<br />
Ankylosauria, Hadrosauridae, most Pachycephalosauria,<br />
Psittacosauridae, some Ceratopsidae. In almost all <strong>of</strong><br />
these taxa, there is evidence that the antorbital cavity is still<br />
present, just internalized.<br />
ACCESSORY CAVITIES AS COMPELLING<br />
MORPHOLOGICAL EVIDENCE FOR<br />
PARANASAL PNEUMATICITY<br />
Accessory cavities are recesses located in the bones surrounding<br />
the antorbital cavity (e.g., lacrimal, maxilla, nasal,













