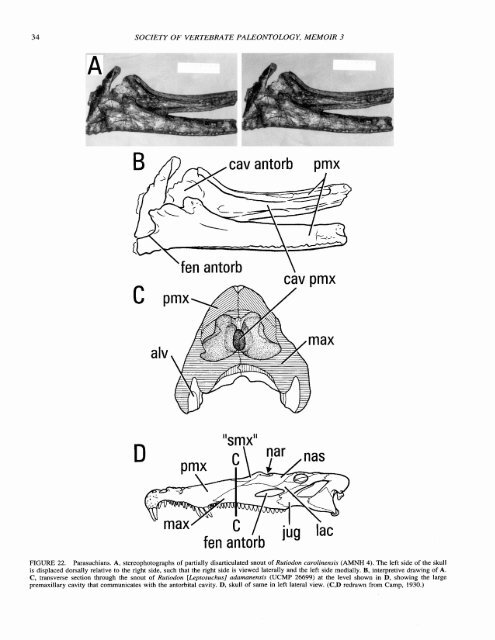
34 SOCIETY OF VERTEBRATE PALEONTOLOGY, MEMOIR 3 cav antorb pmx 'fen antorb \ cav pmx "smx" fen antorb JuY FIGURE 22. Parasuchians. A, stereophotographs <strong>of</strong> partially disarticulated snout <strong>of</strong> Rutiodon carolinensis (AMNH 4). The left side <strong>of</strong> the Skull is displaced dorsally relative to the right side, such that the right side is viewed laterally and the left side medially. B, interpretive drawing <strong>of</strong> A. C, transverse section through the snout <strong>of</strong> Rutiodon [LRprosuchus] adamanensis (UCMP 26699) at the level shown in D, showing the large premaxillary cavity that communicates with the antorbital cavity. D. skull <strong>of</strong> same in left lateral view. (C,D redrawn from Camp, 1930.)
WITMERANTORBITAL ( 7AVITY OF ARCHOSA URS 35 fen antorb int suborb lac pier FIGURE 23. Postosuchus kirkpatricki, facial skeleton. A, left lateral view. B, ventral view. (Redrawn from Chatterjee, 1985.) by an extensive, well-marked antorbital fossa (Fig. 23). In G. stipanicicorum, however, there is no fossa on the lateral surface <strong>of</strong> the jugal ramus <strong>of</strong> the lacrimal or ventrally on the body <strong>of</strong> the maxilla. Desmatosuchus haplocerus (TTUP 9023; see also Case, 1922) is also an exception in that the fossa does not extend ventrally onto the maxilla. The course <strong>of</strong> the nasolacrimal canal is known in the stagonolepidids D. haplocerus (TTUP 9023) and Stagonolepis robertsoni (Fig. 9A,B; Walker, 1961) where it clearly opens orbitally between the lacrimal and prefrontal. The rostra1 course <strong>of</strong> the duct is a little less clear. It probably did not extend to the naris as suggested by Walker (1961) but rather opened into the choanal recess medial to the ascending ramus <strong>of</strong> the maxilla (e.g., S. robertsoni, BMNH R4787), passing dorsomedially around the antorbital cavity. The nasolacrimal canal is present in Postosuchus kirkpatricki (TTUP 9000; Chatterjee, 1985), extending from orbit to antorbital cavity, but it is not clear if it runs within the lacrimal or in the lacrimoprefrontal suture. Crurotarsi: Crocodylomorpha-The general conformation <strong>of</strong> the facial bones in basal crocodylomorphs does not differ substantially from that observed in most other archosaurs, although the trend for reduction and enclosure <strong>of</strong> the antorbital cavity is already evident (see section on facial trends below). The basal sphenosuchians Terrestrisuchus gracilis (Crush, 1984), Saltoposuchus connectens (Huene, 1921 ; Sereno and Wild, 1992), and Pseudhesperosuchus jachaleri (Bonaparte, 1972) retain a relatively large antorbital cavity that excavates a deep antorbital fossa on the extensive medial laminae <strong>of</strong> the maxilla and lacrimal. Thus, the internal antorbital fenestra, bor- dered by the lacrimal and maxilla, is relatively long and low and is much smaller than the external fenestra. A well-developed supralveolar lamina provides a sharp ventral border to the fenestra, and the palatine partially floors the antorbital cavity. The internal antorbital fenestra opens directly opposite the choana within the nasal cavity, as in all sphenosuchians (Figs. 1, 24). The course <strong>of</strong> the nasolacrimal canal is known best in Sphenosuchus acutus where it passes dorsomedially over the antorbital cavity completely within the lacrimal, just internal to the antorbital fossa, to open medially above the dorsal portion <strong>of</strong> the internal antorbital fenestra; the duct presumably continued to the choana along a low ridge on the medial surface <strong>of</strong> the ascending ramus <strong>of</strong> the maxilla (Walker, 1990). The position <strong>of</strong> the orbital aperture <strong>of</strong> the canal in Terrestrisuchus gracilis (Crush, 1984), Saltoposuchus connectens (Sereno and Wild, 1992), and Dibothrosuchus elaphros (IVPP V7907; Fig. 24) implies a similar course in these animals. Basal crocodyliforms (protosuchians) continue the trend <strong>of</strong> reduction <strong>of</strong> the antorbital cavity and external antorbital fenestra. In the forms for which data are available, the internal antorbital fenestra is a relatively small opening bounded by the lacrimal and maxilla (Fig. 25A). It clearly opens medially into the nasal cavity, and it can be seen to be opposite the choana in Protosuchus richardsoni (MCZ 6727, UCMP 130860, AMNH 3024; Crompton and Smith, 1980; Clark, 1986), an unnamed protosuchid (UCMP 97638; Clark, 1986), Orthosuchus stormbergi (Nash, 1975), Gobiosuchus kielanae (Osmblska, 1972; Fig. 25B,C), and Hoplosuchus kayi (CM 11361). The antorbital cavity is floored medially by the palatine in at least Gobiosuchus kielanae (Osm6lska, 1972; Fig. 25B,C). The antorbital fossa is not extensive in any <strong>of</strong> these taxa, but tends to excavate the lacrimal and maxilla caudally, dorsally, and rostrally. It is not clear if the apparently prominent maxillary fossa <strong>of</strong> Hemiprotosuchus leali (Bonaparte, 1972) or the "accessory antorbital depression" <strong>of</strong> Platyognathus hsui (Wu and Sues, 1996) are associated with the antorbital cavity. The course <strong>of</strong> the nasolacrimal canal has been described only for Protosuchus richardsoni, where it extends within the lacrimal from the orbit, dorsomedially over the antorbital cavity, to open into the caudodorsal margin <strong>of</strong> the internal antorbital fenestra (Clark, 1986). Further reduction <strong>of</strong> the antorbital cavity and closure <strong>of</strong> the external antorbital fenestra occurs more than once in Mesoeucrocodylia, how many times depending on the cladogram adopted (see below). Before surveying this taxon, one <strong>of</strong> the osteological correlates, the position <strong>of</strong> the choana, requires clarification. The most obvious synapomorphy <strong>of</strong> Mesoeucrocodylia involves the formation <strong>of</strong> a bony nasopharyngeal duct such that the opening <strong>of</strong> the airway is diverted caudally (Huxley, 1875; Langston, 1973; Benton and Clark, 1988; Clark, 1994; Busbey, 1995). This opening usually is referred to as the "choana." However, as mentioned above for extant crocodilians, the rostral end <strong>of</strong> the nasopharyngeal duct is the primary choana, homologous to the choana <strong>of</strong> other archosaurs (Witmer, 1995b), and the caudal end <strong>of</strong> the duct is a mesoeucrocodilian neomorph, the secondary choana. Thus, in the following discussion, explicit reference will be to only the primary choana. The basal mesoeucrocodilian clade (Buffetaut, 1982; Benton and Clark, 1988; Wu, Li, and Li, 1994), Thalattosuchia, represents the earliest instance <strong>of</strong> extensive reduction <strong>of</strong> the antorbital cavity. The basal thalattosuchian Pelagosaurus typus (Fig. 26; BMNH 32599) resembles teleosaurids such as Steneosaurus spp. (FMNH UC 402, CM 360, MCZ 1063, many BMNH specimens) in having a very small, slitlike external antorbital fenestra between the maxilla and lacrimal with very little to no development <strong>of</strong> an antorbital fossa (Fig. 26A); in some specimens the fenestra may even be absent (see Westphal,