

genomewide characterization of host-pathogen interactions by ...
genomewide characterization of host-pathogen interactions by ...
genomewide characterization of host-pathogen interactions by ...

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
Maren Depke<br />
Introduction<br />
contains all properties for its own insertion into membranes and therefore needs no assistance<br />
<strong>by</strong> proteins like chaperones (Menestrina et al. 2001). Other cytolytic toxins <strong>of</strong> S. aureus are betahemolysin<br />
(hlb), gamma-hemolysin (hlgA, hlgB, hlgC), delta-hemolysin (hld), and the groups <strong>of</strong><br />
leukocidins (luk). The Panton-Valentine leukocidin (PVL, lukF-PV and lukS-PV), a long-known and<br />
well-studied bicomponent toxin, depends in its pore-forming activity on two components, which<br />
first form themselves a heterodimer (Kaneko/Kamio 2004). Also the other leukocidins and Hlg are<br />
bicomponent lysins. Hlg can form either the AB toxin or the CB toxin depending on the<br />
association <strong>of</strong> the subunits to heterodimers.<br />
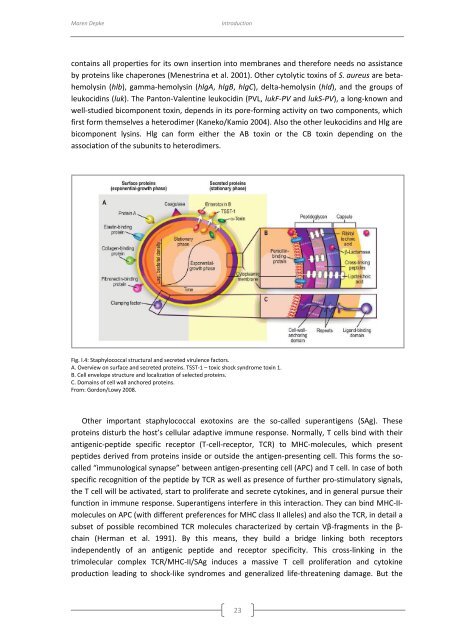
Fig. I.4: Staphylococcal structural and secreted virulence factors.<br />
A. Overview on surface and secreted proteins. TSST-1 – toxic shock syndrome toxin 1.<br />
B. Cell envelope structure and localization <strong>of</strong> selected proteins.<br />
C. Domains <strong>of</strong> cell wall anchored proteins.<br />
From: Gordon/Lowy 2008.<br />
Other important staphylococcal exotoxins are the so-called superantigens (SAg). These<br />
proteins disturb the <strong>host</strong>’s cellular adaptive immune response. Normally, T cells bind with their<br />
antigenic-peptide specific receptor (T-cell-receptor, TCR) to MHC-molecules, which present<br />
peptides derived from proteins inside or outside the antigen-presenting cell. This forms the socalled<br />
“immunological synapse” between antigen-presenting cell (APC) and T cell. In case <strong>of</strong> both<br />
specific recognition <strong>of</strong> the peptide <strong>by</strong> TCR as well as presence <strong>of</strong> further pro-stimulatory signals,<br />
the T cell will be activated, start to proliferate and secrete cytokines, and in general pursue their<br />
function in immune response. Superantigens interfere in this interaction. They can bind MHC-IImolecules<br />
on APC (with different preferences for MHC class II alleles) and also the TCR, in detail a<br />
subset <strong>of</strong> possible recombined TCR molecules characterized <strong>by</strong> certain Vβ-fragments in the β-<br />
chain (Herman et al. 1991). By this means, they build a bridge linking both receptors<br />
independently <strong>of</strong> an antigenic peptide and receptor specificity. This cross-linking in the<br />
trimolecular complex TCR/MHC-II/SAg induces a massive T cell proliferation and cytokine<br />
production leading to shock-like syndromes and generalized life-threatening damage. But the<br />
23














