Zemes un vides zinātnes Earth and Environment Sciences - Latvijas ...
Zemes un vides zinātnes Earth and Environment Sciences - Latvijas ... Zemes un vides zinātnes Earth and Environment Sciences - Latvijas ...
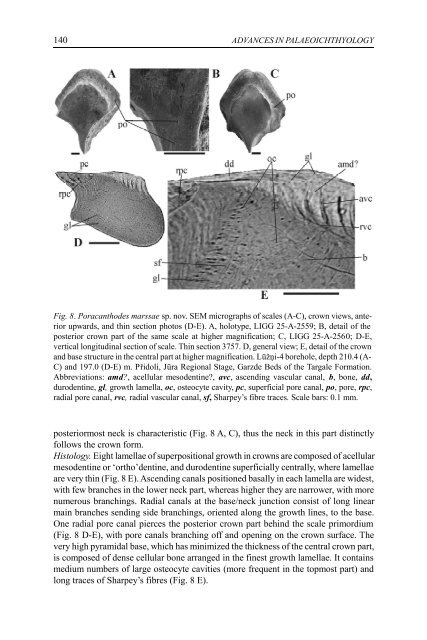
140 ADVANCES IN PALAEOICHTHYOLOGY Fig. 8. Poracanthodes marssae sp. nov. SEM micrographs of scales (A-C), crown views, anterior upwards, and thin section photos (D-E). A, holotype, LIGG 25-A-2559; B, detail of the posterior crown part of the same scale at higher magnification; C, LIGG 25-A-2560; D-E, vertical longitudinal section of scale. Thin section 3757. D, general view; E, detail of the crown and base structure in the central part at higher magnification. Lūžņi-4 borehole, depth 210.4 (A- C) and 197.0 (D-E) m. Přidoli, Jūra Regional Stage, Garzde Beds of the Targale Formation. Abbreviations: amd?, acellular mesodentine?, avc, ascending vascular canal, b, bone, dd, durodentine, gl, growth lamella, oc, osteocyte cavity, pc, superficial pore canal, po, pore, rpc, radial pore canal, rvc, radial vascular canal, sf, Sharpey’s fibre traces. Scale bars: 0.1 mm. posteriormost neck is characteristic (Fig. 8 A, C), thus the neck in this part distinctly follows the crown form. Histology. Eight lamellae of superpositional growth in crowns are composed of acellular mesodentine or ‘ortho’dentine, and durodentine superficially centrally, where lamellae are very thin (Fig. 8 E). Ascending canals positioned basally in each lamella are widest, with few branches in the lower neck part, whereas higher they are narrower, with more numerous branchings. Radial canals at the base/neck junction consist of long linear main branches sending side branchings, oriented along the growth lines, to the base. One radial pore canal pierces the posterior crown part behind the scale primordium (Fig. 8 D-E), with pore canals branching off and opening on the crown surface. The very high pyramidal base, which has minimized the thickness of the central crown part, is composed of dense cellular bone arranged in the finest growth lamellae. It contains medium numbers of large osteocyte cavities (more frequent in the topmost part) and long traces of Sharpey’s fibres (Fig. 8 E).
J. Valiukevičius. Silurian acanthodians from Lūžņi-4 141 Remarks. Crown pores were not visible on examination of scales under a binocular microscope, and they were considered possibly related to Gomphonchus boekschoteni Vergoossen (1999 c, pl. 2, figs 24-25; pl.3, figs 26-31). As for Poracanthodes marssae, most G. boekschoteni scales have large pores opening on all faces of the neck, pearshaped crowns with strengthened and elongated posterior parts, and the similar neck characters including a prominent posterior keel. The grade of the base convexity and its position in advance of the crown in P. marssae were also within the range of the G. boekschoteni morphovarieties. Discovery of pores on crowns of P. marssae precluded assignment to G. boekschoteni and supported attribution of scales to the punctatiform Poracanthodes. Occurrence. See Table. Biostratigraphical potential of acanthodians A single studied locality is insufficient to determine exact acanthodian zonality, but the Lužni-4 core fauna is interesting both in the subdivisional and correlational aspects of Silurian acanthodians in Latvia and the wider region. The series of the Ludlow and Pridoli is here acanthodian-based divided into eleven biostratigraphic intervals named by the chosen index species (Fig. 9, right column), and defined by the first occurrence of the nominative taxon. The intervals are neither interval zones nor partial range zones, because they occupy only some parts of the partial ranges of indices, until the next index appears in the sequence. This fact lowers the stratigraphic value of the interval units. However, the entry levels of the index or other first appearing taxa are significant to the regional record. The most ancient acanthodian of the Lužni-4 borehole is a tiny indeterminable nostolepid (of the putative N. striata-group?) at the depth of 458 m (see Table). The first, Nostolepis gracilis interval starts at the depth of 388 m, about 50 m below the top of the Ventspils Formation, that is coincident with the boundary of Ludlow/ Pridoli. The unit is based on only a few scales of N. gracilis at this single point, which has yielded no other taxon. The finds are significant (I have no doubts concerning their identification), because they mark the earliest occurrence of N. gracilis Gross (1947) ever defined in the Baltic. All prior data (Märss 1986, 1992, 1997, 2000; Märss et al. 1995, 1996) indicated that this species did not exist in the Ludlow, and it was introduced as the Pridolian index fossil of the Vertebrate Standard Scheme. Seemingly, its entry and the biozonal boundary must be lowered into the late Ludlow. The next, Nostolepis latvica sp. nov. interval (338.6-306.2 m) is approximately attached to the Šilale Beds of the Minija Fm. The first occurrence of the index is about 1.3 m above the lower boundary of the Beds, and its range extends to the Venzava Bedss of the Targale Fm. This basal Pridolian acanthodian association is based on the entry of Gomphonchus? minutus sp. nov. and Nostolepis cf. amplifica as the most definitive species (for full list of taxa of this and further intervals see Fig. 9). Its correlational value is yet uncertain, as all three most significant taxa have not been discovered in Lithuania. Of note, N. gracilis starts its ongoing occurrences in samples in the upper part of the interval, above 319.9 m (see Table), about 20 m over the
- Page 89 and 90: O.A. Lebedev. A new tetrapod from R
- Page 91 and 92: O.A. Lebedev. A new tetrapod from R
- Page 93 and 94: O.A. Lebedev. A new tetrapod from R
- Page 95 and 96: O.A. Lebedev. A new tetrapod from R
- Page 97 and 98: O.A. Lebedev. A new tetrapod from R
- Page 99 and 100: ACTA UNIVERSITATIS LATVIENSIS, 2004
- Page 101 and 102: E. Lukševičs, I. Zupiņš. Sedime
- Page 103 and 104: E. Lukševičs, I. Zupiņš. Sedime
- Page 105 and 106: E. Lukševičs, I. Zupiņš. Sedime
- Page 107 and 108: E. Lukševičs, I. Zupiņš. Sedime
- Page 109 and 110: E. Lukševičs, I. Zupiņš. Sedime
- Page 111 and 112: E. Lukševičs, I. Zupiņš. Sedime
- Page 113 and 114: E. Lukševičs, I. Zupiņš. Sedime
- Page 115 and 116: E. Lukševičs, I. Zupiņš. Sedime
- Page 117 and 118: E. Lukševičs, I. Zupiņš. Sedime
- Page 119 and 120: E. Lukševičs, I. Zupiņš. Sedime
- Page 121 and 122: J. Valiukevičius. Silurian acantho
- Page 123 and 124: J. Valiukevičius. Silurian acantho
- Page 125 and 126: J. Valiukevičius. Silurian acantho
- Page 127 and 128: J. Valiukevičius. Silurian acantho
- Page 129 and 130: J. Valiukevičius. Silurian acantho
- Page 131 and 132: J. Valiukevičius. Silurian acantho
- Page 133 and 134: J. Valiukevičius. Silurian acantho
- Page 135 and 136: J. Valiukevičius. Silurian acantho
- Page 137 and 138: J. Valiukevičius. Silurian acantho
- Page 139: J. Valiukevičius. Silurian acantho
- Page 143 and 144: J. Valiukevičius. Silurian acantho
- Page 145 and 146: J. Valiukevičius. Silurian acantho
- Page 147 and 148: J. Valiukevičius. Silurian acantho
- Page 149 and 150: V. Pernegre, V. Dupret. Biostratigr
- Page 151 and 152: V. Pernegre, V. Dupret. Biostratigr
- Page 153 and 154: V. Pernegre, V. Dupret. Biostratigr
- Page 155 and 156: V. Pernegre, V. Dupret. Biostratigr
- Page 157 and 158: V. Pernegre, V. Dupret. Biostratigr
- Page 159 and 160: Ž. Žigaite. New telodont from Tuv
- Page 161 and 162: Ž. Žigaite. New telodont from Tuv
- Page 163 and 164: Ž. Žigaite. New telodont from Tuv
- Page 165 and 166: Ž. Žigaite. New telodont from Tuv
140<br />
ADVANCES IN PALAEOICHTHYOLOGY<br />
Fig. 8. Poracanthodes marssae sp. nov. SEM micrographs of scales (A-C), crown views, anterior<br />
upwards, <strong>and</strong> thin section photos (D-E). A, holotype, LIGG 25-A-2559; B, detail of the<br />
posterior crown part of the same scale at higher magnification; C, LIGG 25-A-2560; D-E,<br />
vertical longitudinal section of scale. Thin section 3757. D, general view; E, detail of the crown<br />
<strong>and</strong> base structure in the central part at higher magnification. Lūžņi-4 borehole, depth 210.4 (A-<br />
C) <strong>and</strong> 197.0 (D-E) m. Přidoli, Jūra Regional Stage, Garzde Beds of the Targale Formation.<br />
Abbreviations: amd?, acellular mesodentine?, avc, ascending vascular canal, b, bone, dd,<br />
durodentine, gl, growth lamella, oc, osteocyte cavity, pc, superficial pore canal, po, pore, rpc,<br />
radial pore canal, rvc, radial vascular canal, sf, Sharpey’s fibre traces. Scale bars: 0.1 mm.<br />
posteriormost neck is characteristic (Fig. 8 A, C), thus the neck in this part distinctly<br />
follows the crown form.<br />
Histology. Eight lamellae of superpositional growth in crowns are composed of acellular<br />
mesodentine or ‘ortho’dentine, <strong>and</strong> durodentine superficially centrally, where lamellae<br />
are very thin (Fig. 8 E). Ascending canals positioned basally in each lamella are widest,<br />
with few branches in the lower neck part, whereas higher they are narrower, with more<br />
numerous branchings. Radial canals at the base/neck j<strong>un</strong>ction consist of long linear<br />
main branches sending side branchings, oriented along the growth lines, to the base.<br />
One radial pore canal pierces the posterior crown part behind the scale primordium<br />
(Fig. 8 D-E), with pore canals branching off <strong>and</strong> opening on the crown surface. The<br />
very high pyramidal base, which has minimized the thickness of the central crown part,<br />
is composed of dense cellular bone arranged in the finest growth lamellae. It contains<br />
medium numbers of large osteocyte cavities (more frequent in the topmost part) <strong>and</strong><br />
long traces of Sharpey’s fibres (Fig. 8 E).


