PDF (Lo-Res) - Smithsonian Institution Libraries
PDF (Lo-Res) - Smithsonian Institution Libraries PDF (Lo-Res) - Smithsonian Institution Libraries
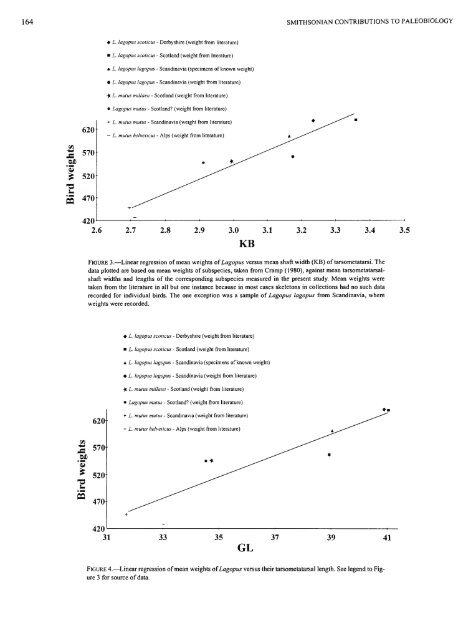
164 Vl 620 M 570 £ 520 S 470f • L. lagopus scoticus - Derbyshire (weight from literature) • L. lagopus scoticus - Scotland (weight from literature) * L. lagopus lagopus - Scandinavia (specimens of known weight) • L. lagopus lagopus - Scandinavia (weight from literature) -fc L. mutus millaisi - Scotland (weight from literature) • Lagopus mutus - Scotland? (weight from literature) -•- L. mutus mutus - Scandinavia (weight from literature) — L. mutus helveticus - Alps (weight from literature) 420 2.6 2.7 2.8 2.9 3.0 KB 3.1 .2f s SMITHSONIAN CONTRIBUTIONS TO PALEOBIOLOGY 3.2 3.3 3.4 3.5 FIGURE 3.—Linear regression of mean weights of Lagopus versus mean shaft width (KB) of tarsometatarsi. The data plotted are based on mean weights of subspecies, taken from Cramp (1980), against mean tarsometatarsalshaft widths and lengths of the corresponding subspecies measured in the present study. Mean weights were taken from the literature in all but one instance because in most cases skeletons in collections had no such data recorded for individual birds. The one exception was a sample of Lagopus lagopus from Scandinavia, where weights were recorded. 620 570 520 470 • L. lagopus scoticus - Derbyshire (weight from literature) • L. lagopus scoticus - Scotland (weight from literature) • L. lagopus lagopus - Scandinavia (specimens of known weight) • L. lagopus lagopus - Scandinavia (weight from literature) -f( L. mutus millaisi - Scotland (weight from literature) • Lagopus mutus - Scotland? (weight from literature) + L. mutus mutus - Scandinavia (weight from literature) - L. mutus helveticus - Alps (weight from literature) • * 420 31 33 35 37 GL 39 41 FIGURE 4.—Linear regression of mean weights ofLagopus versus their tarsometatarsal length. See legend to Figure 3 for source of data.
NUMBER 89 165 18.5 -- 18 - 17.5 17 'i 16.5 16 15.5 15 •- 14.5 14 52 • A A 5 5 A x 5 ° A A ° • o o • • A O • • • % • A X A a »A ° * A • 54 56 58 60 62 Greatest length 5x £ • • 64 66 68 • L. lagopus scoticus - Derbyshire • L. lagopus scoticus - Scotland A L. lagopus lagopus - Scandinavia x L. lagopus lagopus - Russia x L. lagopus brevirostris o Lagopus mutus millaisi- Scotland o Lagopus mutus (Scotland?) A Lagopus mutus islandorum - Iceland D L mutus mutus - Scandinavia 0 L. mutus mutus - Russia A L. mutus helveticus - Alps 5 La Colombiere 1 Pin Hole Cave FIGURE 5.—ScattergTam of humerus length of Lagopus lagopus and Lagopus mutus versus proximal width. certain species. Studies such as that on the pygmy shrew Sorex minutus Linnaeus in northern Europe have shown that where two ecologically similar taxa occur in sympatry their sizes will be more divergent than when in allopatry (Malmquist, 1985). This does not appear to affect Lagopus today, and it could not affect the change in size seen through time because these changes are independent of sympatry or allopatry. Lagopus lagopus and L. mutus are presumably not ecologically similar enough for character displacement to take place. The most often-quoted hypothesis to account for change in body size during the Quaternary is that of climate and, in particular, temperature, which is the mechanism often invoked to account for Bergmann's Rule. Many Pleistocene mammals from glacial episodes were larger than today, and certain authors have suggested that thermoregulation is the causal mechanism (Davis, 1981). Other paleontologists and biologists, however, have agreed that this mechanism has been applied where it may not be appropriate, and that the subject is a much more complex one (Lister, 1992). A counterargument proposed by Guthrie (1984, 1990) and Geist (1986) is that it is not the climate that directly affects an animal's size but the consequences of the length and quality of the plant growing season, which in turn are affected by climate. The vegetational environment, called steppe-tundra or mammoth-steppe, has been described as very productive on the basis of the large herbivores it supported (Guthrie, 1990). The vegetation was a mosaic of high diversity, although predominated by grassland. It should be noted, however, that some palynologists have disagreed with the concept of the mammoth-steppe. They believe the vegetation was poor, a polar desert, based on the apparently low pollen influx at the time. The idea that the vegetational environment was a rich steppe-tundra has recently been expanded by Lister and Sher (1995), who have suggested that the steppe-tundra vegetation relied on a climatic regime that has vanished. They pointed out that detailed climatic records, such as studies of the Greenland ice cores, have shown that the Holocene is distinct from the late Pleistocene in having unusually stable conditions. Pleistocene climatic instability may have allowed the mosaic vegetation of the steppe-tundra to persist. Once this climatic regime ceased to exist, the megafauna, which relied so heavily on the vegetation type the climate supported, changed along with it. Some animals became extinct, like the giant deer Megaloceros giganteus (Blumenbach) and the woolly rhinoceros Coelodonta antiquitatis (Blumenbach), or locally extinct, like the lion Panthera leo Linnaeus and spotted hyena Crocuta crocuta Erxleben (Stuart, 1991). Others underwent a reduction in body size, such as the fox Vulpes vulpes Linnaeus and wild boar Sus scrofa Linnaeus (Davis, 1981). It is, therefore, an attractive hypothesis that certain
- Page 124 and 125: 114 SMITHSONIAN CONTRIBUTIONS TO PA
- Page 126 and 127: 116 SMITHSONIAN CONTRIBUTIONS TO PA
- Page 128 and 129: 118 SMITHSONIAN CONTRIBUTIONS TO PA
- Page 130 and 131: 120 SMITHSONIAN CONTRIBUTIONS TO PA
- Page 132 and 133: 122 SMITHSONIAN CONTRIBUTIONS TO PA
- Page 135 and 136: The Middle Pleistocene Avifauna of
- Page 137: NUMBER 89 Accordi, B. 1962. La grot
- Page 140 and 141: 130 FIGURE 1.—Map showing locatio
- Page 142 and 143: 132 Cranium Mandibula Scapula Clavi
- Page 144 and 145: 134 SMITHSONIAN CONTRIBUTIONS TO PA
- Page 146 and 147: 136 SMITHSONIAN CONTRIBUTIONS TO PA
- Page 149 and 150: Seabirds and Late Pleistocene Marin
- Page 151 and 152: NUMBER 89 141 METHODS Only strictly
- Page 153 and 154: NUMBER 89 143 FIGURE 2.—Area of s
- Page 155 and 156: NUMBER 89 145 FIGURE 4.—Area of s
- Page 157 and 158: NUMBER 89 147 FIGURE 6.—Area of s
- Page 159 and 160: NUMBER 89 149 FIGURE 8.—Area of s
- Page 161 and 162: NUMBER 89 151 FIGURE 10.—Area of
- Page 163 and 164: NUMBER 89 153 FIGURE 12.—Area of
- Page 165 and 166: NUMBER 89 155 the period studied. T
- Page 167: NUMBER 89 157 Walker, C.A., G.M. Wr
- Page 170 and 171: 160 SMITHSONIAN CONTRIBUTIONS TO PA
- Page 172 and 173: 162 SMITHSONIAN CONTRIBUTIONS TO PA
- Page 176 and 177: 166 birds, such as the two species
- Page 178 and 179: 168 SMITHSONIAN CONTRIBUTIONS TO PA
- Page 180 and 181: 170 cional Autonoma de Mexico, for
- Page 182 and 183: 172 SMITHSONIAN CONTRIBUTIONS TO PA
- Page 184 and 185: 174 ated with this specimen, see Mi
- Page 187 and 188: The Fossil Record of Condors (Cicon
- Page 189 and 190: NUMBER 89 179 FIGURE 2.—Geographi
- Page 191 and 192: NUMBER 89 181 FIGURE 5.—Vulturida
- Page 193 and 194: NUMBER 89 183 FIGURE 7.—Referred
- Page 195 and 196: Two New Fossil Eagles from the Late
- Page 197 and 198: NUMBER 89 187 TABLE 1.—Measuremen
- Page 199 and 200: NUMBER 89 189 carpal trochlea relat
- Page 201 and 202: NUMBER 89 191 FIGURE 4.—Holotypic
- Page 203 and 204: NUMBER 89 193 We compared the parat
- Page 205 and 206: NUMBER 89 195 FIGURE 6.—Distribut
- Page 207 and 208: NUMBER 89 197 the Florida State Mus
- Page 209 and 210: A New Genus of Dwarf Megapode (Gall
- Page 211 and 212: NUMBER 89 201 lis hypotarsi along t
- Page 213 and 214: NUMBER 89 203 The fossil is larger
- Page 215 and 216: NUMBER 89 205 Clark, George A., Jr.
- Page 217 and 218: A New Genus and Species of the Fami
- Page 219 and 220: NUMBER 89 209 son with other known
- Page 221 and 222: NUMBER 89 211 FIGURE 1.—Argornis
- Page 223 and 224: NUMBER 89 213 AM AL AM AL AM AL AM
164<br />
Vl<br />
620<br />
M 570<br />
£ 520<br />
S 470f<br />
• L. lagopus scoticus - Derbyshire (weight from literature)<br />
• L. lagopus scoticus - Scotland (weight from literature)<br />
* L. lagopus lagopus - Scandinavia (specimens of known weight)<br />
• L. lagopus lagopus - Scandinavia (weight from literature)<br />
-fc L. mutus millaisi - Scotland (weight from literature)<br />
• Lagopus mutus - Scotland? (weight from literature)<br />
-•- L. mutus mutus - Scandinavia (weight from literature)<br />
— L. mutus helveticus - Alps (weight from literature)<br />
420<br />
2.6 2.7 2.8 2.9 3.0<br />
KB<br />
3.1<br />
.2f<br />
s<br />
SMITHSONIAN CONTRIBUTIONS TO PALEOBIOLOGY<br />
3.2 3.3 3.4 3.5<br />
FIGURE 3.—Linear regression of mean weights of Lagopus versus mean shaft width (KB) of tarsometatarsi. The<br />
data plotted are based on mean weights of subspecies, taken from Cramp (1980), against mean tarsometatarsalshaft<br />
widths and lengths of the corresponding subspecies measured in the present study. Mean weights were<br />
taken from the literature in all but one instance because in most cases skeletons in collections had no such data<br />
recorded for individual birds. The one exception was a sample of Lagopus lagopus from Scandinavia, where<br />
weights were recorded.<br />
620<br />
570<br />
520<br />
470<br />
• L. lagopus scoticus - Derbyshire (weight from literature)<br />
• L. lagopus scoticus - Scotland (weight from literature)<br />
• L. lagopus lagopus - Scandinavia (specimens of known weight)<br />
• L. lagopus lagopus - Scandinavia (weight from literature)<br />
-f( L. mutus millaisi - Scotland (weight from literature)<br />
• Lagopus mutus - Scotland? (weight from literature)<br />
+ L. mutus mutus - Scandinavia (weight from literature)<br />
- L. mutus helveticus - Alps (weight from literature)<br />
• *<br />
420<br />
31 33 35 37<br />
GL<br />
39 41<br />
FIGURE 4.—Linear regression of mean weights ofLagopus versus their tarsometatarsal length. See legend to Figure<br />
3 for source of data.


