1 Neuroscience C 3045; Eye and Brain Professor M. Glickstein ...
1 Neuroscience C 3045; Eye and Brain Professor M. Glickstein ...
1 Neuroscience C 3045; Eye and Brain Professor M. Glickstein ...

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
<strong>Neuroscience</strong> C <strong>3045</strong>; <strong>Eye</strong> <strong>and</strong> <strong>Brain</strong><br />
<strong>Professor</strong> M. <strong>Glickstein</strong><br />
January 2012<br />
The general problem of vision; what sorts of things can you do with your eyes? The<br />
eye, like other excellent optical devices forms an image of the world in front of it. Visual<br />
direction is probably innate. In addition there is vision for:<br />
Fine detail: Detection of a small object against a background (visual acuity). Clinical<br />
measures 20/20 [6/6] you see @ 20 feet what a normal eye sees at 20 feet. Acuity<br />
falls off in peripheral retina.<br />
Sensitivity: How much light do you need to see? Each rod can be activated by a single<br />
photon.<br />
Visual Depth <strong>and</strong> Distance Perception: How far away is an object? Which of two objects is<br />
closer?<br />
Monocular Mechanisms (e.g. Motion parallax; relative angular movement of 2 objects<br />
at different distances).<br />
Binocular Stereopsis Fusing of 2 disparate images to form a depth cue.<br />
Colour Vision: Identifying, naming, discriminating colours. How is colour processed in the<br />
brain?<br />
Form vision: Recognition of your friend's car in front of his house; Recognition of your<br />
friend's face.<br />
Movement Detection:<br />
Is an object stationary or moving?<br />
Visual guidance of your own movement.<br />
<strong>Eye</strong> movements (saccadic, pursuit; vergence).<br />
Accommodation<br />
Fingers <strong>and</strong> limbs (grasping an object in front of you)<br />
Whole body; walking along a crowded street: climbing a tree or the north fact of the<br />
Eiger.<br />
Related problem of distinguishing self-movement from movement of something in our<br />
field of view.<br />
The constancies: Why does brightness remain constant over a broad range of illumination<br />
intensities? Lump of coal in sunlight.<br />
When we consider the anatomy <strong>and</strong> physiology of the visual pathways, remember that it is<br />
these phenomena of vision that direct <strong>and</strong> focus research. How does the brain do it?<br />
1
Comparative anatomy of the vertebrate <strong>Eye</strong><br />
Discuss the classical light-microscopic view of the structure of the vertebrate eye with some<br />
general principles that apply to all vertebrates <strong>and</strong> examples of specialised structures that<br />
have evolved in one or another form.<br />
Similarity. To a great degree, all vertebrate eyes are similar. They share an almost identical<br />
imaging system (modified in special environments, such as under water).<br />
Compare with compound eye.<br />
Basic Cellular Plan The basic structural elements of the retina are similar in all vertebrates.<br />
Three nuclear layers with five or six cell-types involved in visual transduction<br />
<strong>and</strong> coding<br />
Two synaptic (plexiform) layers<br />
Evolution; Darwin’s difficulty<br />
“….if numerous gradations from a perfect <strong>and</strong> complex eye to one very<br />
imperfect <strong>and</strong> simple, each grade being useful to its possessor can be show to<br />
exist…then the difficulty… can hardly be considered real” See article by Lamb<br />
et.al. in reference list.<br />
Normal Human <strong>Eye</strong><br />
3 "Tunics" of eye<br />
A: Sclera<br />
B: Choroid <strong>and</strong> iris muscles<br />
C: Retina (<strong>and</strong> pigment epithelium)<br />
Note also focusing apparatus; cornea <strong>and</strong> lens <strong>and</strong> the optic nerve<br />
Differences. Where vertebrate eyes differ it is often related to the light environment in which<br />
the animal lives.<br />
Range of Light Intensity. Range of Physical Intensity approaches 10 12 . No sense organ<br />
could work effectively over such a range (10 3 /second action potentials).<br />
How to Code? Scale compression (Weber's law). We are sensitive to relative, not absolute<br />
light levels.<br />
How to deal with great range in light intensities<br />
1. Photomechanical Changes<br />
Physically restrict the amount of light which is allowed to reach receptors<br />
2. Parallel set of receptors (rods <strong>and</strong> cones) with different threshold <strong>and</strong> different<br />
dynamic range<br />
3. "Tuning" ganglion cells so that they report differences in light intensities within<br />
receptive field (lump of coal in sunlight looks black)<br />
2
Mechanical Regulation of Light at the Receptor<br />
Pupils<br />
Tiger<br />
What's wrong with the picture?<br />
Association of slits with nocturnal life style (esp. Snakes)<br />
Round vs. slit pupils. Nocturnal retina requires exclusion of light in daylight "basking"<br />
Pigments <strong>and</strong> stray light. Stray light degrades the image, it is absorbed in cameras <strong>and</strong> in the<br />
eye by pigment layers.<br />
Monkey<br />
Pigment epithelium <strong>and</strong> choroid<br />
Pigment epithelium plays an especially important role in light <strong>and</strong> dark adaptation of lower<br />
vertebrates. Processes interdigitate with tips of receptors.<br />
Pigment epithelium (Hedgehog)<br />
Pigment epithelium is a channel for (choroidal) vascular supply for receptors. Bites off<br />
growing rod tips.<br />
Pigment migration in lower vertebrate. The processes of pigment epithelial cells may extend<br />
the length of the receptors.<br />
Image 2 hairbrushes (receptors)<br />
Melanin granules are actively migrating:<br />
Migrate outward in light back (towards cell body) in dark<br />
Dark-adapted fish eye<br />
Light adapted fish eye<br />
Augmentation of Sensitivity. Animals that live in dim light-environment sacrifice precision<br />
of an image for increased sensitivity "snapshots vs. time exposures"<br />
Suppose light goes through receptor layer without ever being absorbed. If it strikes a<br />
reflective surface the receptors have a second chance<br />
<strong>Eye</strong> shine Leopard<br />
Tapetum Lucidum<br />
(Note Absence of melanin pigment in pigment epithelium)<br />
Toxicity of Excessive Light<br />
Albinos are especially at risk. Four weeks of continuous moderate illumination; albino rat<br />
causes degeneration of receptors<br />
Normal rat retina<br />
Constant light; retinal degeneration<br />
Retinal structure <strong>and</strong> the accuracy of ophthalmic instruments. To a reasonable approximation<br />
the retina of all mammals (vertebrates) is of constant thickness.<br />
Rat<br />
Whale<br />
Receptors <strong>and</strong> inner nuclear cells are roughly equivalent in size <strong>and</strong> number; related to the<br />
fact that these retinal elements do not conduct action potentials.<br />
Receptors <strong>and</strong> inner nuclear cells convey information over a short length by graded potentials<br />
(receptors, bipolar <strong>and</strong> horizontals some amacrine)<br />
Ganglion cell carries impulses to brain. The distance varies from a few millimetres to many<br />
centimetres. Ganglion cell sizes vary greatly. There is also variability within a single retina:<br />
Size of cell body is related to fibre diameter.<br />
Consequences of finite retinal thickness for ophthalmoscope <strong>and</strong> retinoscope. Shine light in;<br />
observe reflected shadow<br />
3
Suppose light is reflected from layer other than rods <strong>and</strong> cones. For thin lens in air<br />
D = f -- 1 by definition<br />
- 2<br />
dD = -f<br />
df or error (if constant) should be a function of the inverse square<br />
of the focal length<br />
Raw data; <strong>Eye</strong> size <strong>and</strong> refraction<br />
Log plot<br />
Conclude: retinoscope uses a specular reflection from vitreous-retinal surface<br />
Similar factor applies to opthalmoscope<br />
Growth <strong>and</strong> Plasticity of <strong>Eye</strong> Size; Emmetropization<br />
The normal newborn human infant eye measures 17 mm. from cornea to retina; the adult eye<br />
24 mm.<br />
The (strange but true ) evidence that eye growth is under local visual control.<br />
Different focal planes in the bird eye<br />
Fundamental Dichotomy of Rods <strong>and</strong> Cones<br />
Schultze's dichotomy: Differences in receptor types correlated with the life of the animal:<br />
"pure rod" <strong>and</strong> "pure cone" retinas.<br />
Retina is mixed rod/cone<br />
Monkey retina 3 layers<br />
Rod/cone (nuclei). Note Difference in nuclei of rods <strong>and</strong> cones.<br />
Higher power; conical outer segments<br />
Variability in cone density in human (an monkey) retina; rods are relatively constant<br />
Peripheral retina<br />
10 0 OUT<br />
Follow colour of nuclei<br />
Edge of fovea<br />
Central fovea<br />
Other Plans; The Retinal Streak<br />
Lindsay Johnson reindeer<br />
Note 1) Pigment distribution<br />
2) Vascular differences<br />
Receptors <strong>and</strong> Visual Pigments<br />
Wavelength sensitivity of Receptors.<br />
Outer segments of rods <strong>and</strong> cones contain photopigments.<br />
1) Absorb light 2) Transduce physical signal into an electrical one:<br />
Wavelength <strong>and</strong> Absorption<br />
Rods peak absorption at 500 nm<br />
2) Absorption spectra of pigment in cones. In human <strong>and</strong> monkey. 3 classes by<br />
microspectrophotometry; intracellular recording.<br />
Requirements for Colour Vision<br />
Colour vision requires at least 2 classes of receptor with different wavelength subdivisions.<br />
In principle you could do it either with pigments or with filters.<br />
Oil droplets<br />
Variability in Receptor Morphology Cones <strong>and</strong> Rods may be Thin or Fat<br />
Frog; Rods <strong>and</strong> cones<br />
4
Note massive rod (also "green" rods; double cones)<br />
Now cut parallel to the inner segments<br />
Retinal mosaic in Gecko.<br />
Gecko retina<br />
Synaptic connections of the retina<br />
Structure of the eye; Cajal 1892<br />
Cajal; Mammalian retina<br />
elements: receptors contact both horizontal <strong>and</strong> bipolar cells<br />
bipolar cells contact amacrine <strong>and</strong> ganglion cells<br />
amacrine cells contact bipolars <strong>and</strong> ganglion cells<br />
ganglion cells as final common path<br />
There are two synaptic layers in the eye. Outer plexiform layer containing cone pedicles, rod<br />
spherules; bipolar <strong>and</strong> horizontal cell processes<br />
Much more complex inner plexiform layer. Many cell types in the ganglion cell layer.<br />
Laminar structure of inner plexiform layer<br />
Ground squirrel<br />
Differences in ganglion cell size<br />
Cat retina<br />
Functional differences between ganglion cells of different morphology.<br />
1. Large somas have largest-calibre axons (m cells: transient properties)<br />
2. Smaller sized (p cells)<br />
On vs. off centre ganglion cells are associated with different planes within the inner<br />
plexiform layer<br />
The conduction pathway within the eye. Axons take a varied course, some sweeping away<br />
from the fovea enroute to optic nerve (myelinated only when they reach the disc hence non<br />
opaque)<br />
Note Blue Arcs in demonstrations<br />
Monkey whole mount<br />
Convergence & Divergence within the Retina<br />
Receptors as "points"<br />
Ganglion cells as transmitters<br />
Great variability in the complexity of inner nuclear layer<br />
Convergence is generally an adaptation for sensitivity<br />
Hamster<br />
But many animals (especially birds!) have massive inner nuclear layer: an active filter<br />
Tree shrew<br />
Humans <strong>and</strong> monkeys have highly convergent peripheral retina roughly equal number<br />
of receptors/inner nuclear cells/ganglion cells near fovea<br />
Monkey fofea<br />
Why is fovea shaped as it is?<br />
Owl fovea<br />
Chameleon (Anolis)<br />
5
Two aims;<br />
Central visual pathways <strong>and</strong> visuo-motor links<br />
`Introduce the targets of the eye, <strong>and</strong> particularly the primary visual cortex<br />
Discuss the connections from visual areas of the cortex to motor areas.<br />
I will talk about the connections from the eye, the nature of the crossing at the optic chiasma,<br />
<strong>and</strong> the primary <strong>and</strong> secondary targets of the optic tracts. I will introduce these topics in a<br />
historical context; how did we learn about the primary visual cortical area?<br />
I will review the evidence that led to our recognition of a primary motor area of the cerebral<br />
cortex, <strong>and</strong> early ideas about how the visual input might be linked to the motor output for<br />
visual guidance of the arm, the legs <strong>and</strong> the fingers. There are three major ways in which the<br />
visual areas might be linked to motor areas;<br />
1) Via a series of cortico-cortical links<br />
2) By way of basal ganglia<br />
3) A major pathway from cortex to the cerebellum by way of the pontine nuclei<br />
THE EYE AND THE CENTRAL VISUAL PATHWAYS<br />
. Much of the visual system can be thought of as in a series-like organization.<br />
Slide 1: Rhesus monkey retina ; low power to show series like arrangement of<br />
layers<br />
Receptors: (Rod/cones) Inner nuclear cells: (Horizontal, bipolar, amacrine) to ganglion<br />
cells.<br />
Visual information is imaged on the photoreceptors <strong>and</strong> must be relayed via inner-nuclear<br />
layer to ganglion cells for transmission to the brain.<br />
Centre-surround organization of retinal ganglion cells <strong>and</strong> scaling of relative intensity.<br />
Within this series-like arrangement, there are at every level, parallel processing mechanisms.<br />
Easiest to see at the level of the photoreceptors themselves.<br />
Rods <strong>and</strong> cones share the image plane: process visual information in parallel.<br />
Slide 2: High power to show rod <strong>and</strong> cone nuclei<br />
At the retinal ganglion-cell level. There are ganglion cells with grossly different structure,<br />
connections <strong>and</strong> function.<br />
Ratio of receptor to ganglion cells varies greatly<br />
Slide 3: Cat retina (note tepetum) Note differences in ganglion cell size<br />
Galgion cells axons project to brain<br />
Note large <strong>and</strong> smaller ganglion cells (m <strong>and</strong> p cells). Correlated with fibre-diameter.<br />
These different sized cells have different receptive field properties, different targets in the<br />
brain <strong>and</strong> different functions.<br />
6
Large cells (m cells); respond transiently to changes of illumination in their receptive field;<br />
<strong>and</strong> there are relatively more of them in peripheral retina.<br />
Smaller sized (p cells); respond in a sustained manner to appropriate illumination of their<br />
receptive field, have small receptive fields <strong>and</strong> are more heavily concentrated near fovea.<br />
Much smaller cells “k” cells (konio; dust) <strong>and</strong> light detection<br />
There is a smaller class of cells, the so-called “k” cells which have different receptive field<br />
properties <strong>and</strong> different structures. K cells only account for an estimated 10% of all retinal<br />
ganglion cells, but they seem to have important functions as light detectors.<br />
An interesting class of ganglion cell has been discovered relatively recently; the melanopsin<br />
containing ganglion cells. These cells may receive an input from photoreceptors, but they<br />
also have a photosensitive pigment within them. Melanopsin ganglion cells respond to a short<br />
wavelength of 484 nm peak sensitivity.<br />
“Tiling of retinal ganglion cells” <strong>and</strong> the extent of ganglion cell dendritic tree<br />
Slide 4: Cajal; Mammalian retina<br />
See reference to Dacey et. al. in reference list<br />
The three cell types cover the entire retina. Since there are about 80% p cells, 10% m cells<br />
<strong>and</strong> an estimated 10% k cells, it follows that if the entire retina is covered by the dendrites of<br />
these cells, the m <strong>and</strong> k cells must have larger dendritic territories than the p cell.<br />
Magnocellular/parvocellular division of LGN; Also interstitial cells<br />
Axons of the optic nerve<br />
Axons of the ganglion cell collect at the back of the eye after running along the inner retinal<br />
surface as unmyelinated fibres. (Why are they not yet myelinated?)<br />
Blind spot <strong>and</strong> exit zone.<br />
Slide 5: Monkey eye to show optic nerve (blue-arcs in demo)<br />
On the course of the optic nerve<br />
Optic nerve fibres proceed backwards <strong>and</strong> seem to unite in the X-shaped Optic Chiasma<br />
(Greek X = Chi). The re-sorted fibres after the chiasma are called the optic tract.<br />
At the chiasma the optic nerve fibres arising from ganglion cells in the nasal retina cross;<br />
those from the temporal retina remain uncrossed.<br />
The effect of this pattern of crossing is to project the right visual field onto the left<br />
hemisphere.<br />
Slide 6: To show location of chiasma at the ventral surface of the brain<br />
There was no agreement on what happens at the chiasma. Some believed in no<br />
decussation, some total decussation. (Isaac Newton had suggested the correct<br />
arrangement)<br />
There was always the problem of why we see singly with two eyes. Solved in different ways<br />
by different authors.<br />
Slide 7: Des Cartes; Chiasma <strong>and</strong> pineal<br />
Slide 8: Hemi decussation; John Taylor; Frankfort 1750<br />
Left Visual Field is projected onto the right side of the brain.<br />
On the consequences of lesions 1) before the chiasma 2) of the crossing fibres in the chiasma<br />
3) of the optic tract <strong>and</strong> beyond.<br />
HEMI-AN-OPIAS<br />
7
Homonymous (right or left)<br />
Heteronymous (bitemporal or binasal)<br />
Mnemonic: a person (with optic tract), visual radiations, or complete unilateral cortical<br />
lesion sees only on the side of the lesion.<br />
Partial decussation in dogs, cats <strong>and</strong> horses. How far lateral is the eye? Principal of Visual<br />
Field Representation in mammals.<br />
Slide 9: Tree shrew to show laterality of the eyes<br />
Slide 10: Tree Shrew; LGN <strong>and</strong> optic tract<br />
On the targets of the optic tract<br />
The targets in terms of relative numbers of fibres in man are:<br />
Lateral Geniculate Nucleus (LGN). The major target in humans<br />
Superior Colliculus<br />
Pretectal Nucleus; role in pupillary control<br />
Accessory optic system<br />
Hypothalamus particularly the suprachiasmatic nucles. Role in diurnal cycling.<br />
Lateral Geniculate Nucleus in turn relays visual information to the cortex.<br />
The Laminar Organization of the LGN.<br />
Slide 11: Human LGN (Recall centre surround receptive fields of LGN cells<br />
Strict segregation of input, but perefectly aligned. Problem of the blind spot.<br />
Phenomenon of transneuronal atrophy (Minkowski, 1920)<br />
Slide 12: LGN: Normal monkey<br />
Slide 13: Trans-neuronal atrophy<br />
Numbering from ventral to dorsal 1-6<br />
Contralateral eye projects to layers 1, 4 <strong>and</strong> 6<br />
Ipsilateral eye projects to layers 2, 3 <strong>and</strong> 5<br />
Note important distinction of<br />
Magnocellular layers (1 <strong>and</strong> 2) <strong>and</strong><br />
Parvocellular layers (3, 4, 5, <strong>and</strong> 6) laminae<br />
K cell projection to cells within the interlaminar fibre layers.<br />
Receptive fields of cells in the LGN<br />
Magnocellular layers relay m cells; achromatic; transient<br />
Parvocellular layers relay p cells colour-opponent; sustained<br />
K cells as light detectors; other properties<br />
Slide 14: Human colliculus; parasagittal<br />
The superior colliculus<br />
The superior colliculus receives a direct input from the optic tract <strong>and</strong> from the cerebral<br />
cortex.<br />
Importantly involved in the control of saccadic eye movements.<br />
Lesions of superior colliculus produce a transient deficit in initiating eye movements. Large<br />
lesion of the entire colliculus leads to a transient fixed gaze; which recovers in a few days.<br />
Lesions of Area 8 of the cerebral cortex (the "frontal eye fields") also produce a temporary<br />
deficit in initiating eye movements.<br />
Combined bilateral lesions of both lead to a permanent <strong>and</strong> severe deficit in initiating eye<br />
movements.<br />
8
Pretectal Nuclei <strong>and</strong> the pathway for the pupillary reflex<br />
Direct <strong>and</strong> consensual pupillary reflexes illumination of the eye causes pupillary constriction<br />
Retinal ganglion cells (certain small cells) Pretectal nuclei<br />
Edinger Westphal nucleus in midbrain <strong>and</strong> thence via NIII to Ciliary ganglion controlling<br />
Radial muscles of the iris.<br />
Melanopsin ganglion cells as another input to pupillary control.<br />
THE GENICULO-CORTICAL PROJECTION<br />
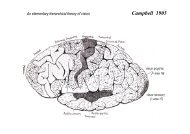
Slide 15: Brodmann, 1905; Monkey cortical areas, lateral<br />
Slide 16: Brodmann, 1905; Medial view<br />
Slide 17: Human Weil stain to show stripe of Gennari<br />
Segregation of the input from the left <strong>and</strong> right eye in the visual cortex; ocular dominance<br />
columns.<br />
We saw that projections from the left <strong>and</strong> right eyes are segregated in their connections to<br />
LGN. This segregation is maintained at the first cortical synapse (in lamina 4 of cortex). The<br />
segregation shows up in both histological <strong>and</strong> physiological experiments.<br />
Inject one eye of a monkey with a labelled amino acid. Taken up <strong>and</strong> transported to LGN.<br />
Some labelled protein taken up by neurons across the synapse <strong>and</strong> transported by LGN<br />
neurons to the cortex. Terminals of left & right eye run in parallel stripes of about 0.5 mm<br />
each on the cortex within layer IV.<br />
Slide 17: Ocular dominance columns; Hubel<br />
If one eye is masked in infancy vision is severely impaired in the deprived eye, the ocular<br />
dominance columns become asymmetric; <strong>and</strong> few cells can be activated by the deprived eye.<br />
"Take over" is failure to retract terminals.<br />
On the normal growth of the eye in development<br />
Slide 18: Human eye; neonate <strong>and</strong> adult<br />
Slide 19: Wiesel <strong>and</strong> Raviola ; Normal <strong>and</strong> form-deprived eye<br />
The form-deprived eyegrows much longer, <strong>and</strong> is seriously myopic<br />
Cells in more superficial <strong>and</strong> deep layers of cortex begin the process of fusing the input from<br />
the two eyes;<br />
Cytochrome oxidase "patches" or “blobs”in primary visual cortex<br />
Livingston <strong>and</strong> Hubel showed that colour information is targeted on the cytochrome oxidase<br />
patches. But nocturnal primates who lack colour vision also have the cytochrome oxidase<br />
patches.<br />
Multiple visual areas<br />
Slide 20: Brodmann; lateral view of monkey brain<br />
Slide 21: Brodmann; Medial view of monkey brain<br />
Is the striate cortex the sole target of LGN axons? Old evidence form the cat, recent<br />
evidence in monkeys. Macular sparing “blindsight” <strong>and</strong> other odds <strong>and</strong> ends<br />
9
VISUAL GUIDANCE OF MOVEMENT<br />
1. Some general problems: What do you want to move?<br />
Slide 1) YO YO MA PLAYING THE CELLO<br />
Slide 2) Bouree from Bach’s 3d suite for unaccompanied cello<br />
Demo Catching a small object<br />
Slide 3) (MONKEY REACHING FOR A PELLET<br />
Arms, legs etc. (catching a cricket ball/baseball)<br />
Fingers <strong>and</strong> wrist (threading a needle)<br />
<strong>Eye</strong>s: On the categories of voluntary eye movements conjugate vs vergence<br />
Smooth pursuit vs saccadic<br />
Why has so much attention been paid to eye movements?<br />
General problem of load compensation in limbs;<br />
<strong>Eye</strong>s always weigh the same<br />
How are visual areas connected to motor areas of the brain.<br />
Visual cortex to motor cortex; (superior colliculus to red nucleus).<br />
Three possible routes: 1) Cortico-cortical circuits. 2) Basal Ganglia, 3) Cerebellar<br />
Side look into history. The discovery of the motor cortex<br />
Slide 4: FRITSCH AND HITZIG<br />
Electrical stimulation<br />
Slide 5: MOTOR CORTEX<br />
The first attempt to identify the visual area led to an error.<br />
FERRIER: "CAUSING BLINDNESS"<br />
Ferrier's mistake was gently pointed out to him by Hermann Munk<br />
MUNK: OCCIPITAL LESIONS CAUSES HEMIANOPIA<br />
SALOMON HENSCHEN “HEMIANOPISCHE FÄLLE”<br />
“KEINE HEMIANOPSIE”<br />
By the end of the nineteenth century, the identity <strong>and</strong> location of the primary visual<br />
<strong>and</strong> motor areas of the human <strong>and</strong> monkey cerebral cortex were clear. Less was understood<br />
about further processing of the visual signal.<br />
BRODMANN LATERAL VIEW<br />
Note Area 18 <strong>and</strong> 19; "Association areas"<br />
BRODMANN MEDIAL VIEW<br />
Parietal Lobe Lesions Cause Visuo Motor Impairment<br />
Why did Ferrier make the mistakes that he did? In his earliest experiments, Ferrier operated<br />
in unsterile conditions. Because of the risk of infections, he killed the monkeys three days<br />
after they were operated on. He later joined the faculty of the Medical School at King's <strong>and</strong><br />
learned sterile operative technique from his colleague, Joseph Lister.<br />
His animals could now live for weeks, months or years. In the second edition of his book,<br />
Functions of the <strong>Brain</strong>, he modified his statement<br />
FERRIER: "TEMPORARY COMPLETE ...."<br />
Angular gyrus <strong>and</strong> parietal lobe<br />
BALINTS CASE<br />
BALINTS DESCRIPTION<br />
What is the Circuit between Visual cortex <strong>and</strong> Motor Cortex?<br />
10
BERNSTEIN 1900<br />
The discovery of the motor cortex. Fritsch & Hitzig's Experiment<br />
The discovery of the visual cortex. Ferrier's Mistake: Munk<br />
The "obvious" nature of cortico-cortical links.<br />
If cortico-cortical circuits are essential, the first link must be by way of the parietal lobe<br />
.<br />
Special problem of integration between the hemispheres (backh<strong>and</strong>ed shot in tennis).<br />
The evidence (or lack of it) for the role of cortico-cortical fibres.<br />
. Section of corpus callosum does not produce obvious deficits in bimanual<br />
coordination or sensory guidance of movement;<br />
Direct cuts of cortico-cortical fibres do not block visual guidance of the h<strong>and</strong> <strong>and</strong> arm.<br />
(Myers, Sperry & McCurdy)<br />
MYERS ET AL 1963<br />
But see Kuyper's interpretation<br />
.<br />
The Corticoponto-cerebellar pathway.<br />
Corticopontine (<strong>and</strong> tectopontine) fibres:<br />
HUMAN BASIS PEDUNCULI<br />
How to trap them<br />
Basis pedunculi - (pyramidal tract + corticocular fibres)<br />
Corticopontine fibres<br />
HUMAN MEDULLA<br />
PONS<br />
MATANO, STEPHAN<br />
How Does the cerebral Cortex Connect to the Pontine Nuclei?<br />
Two related questions;<br />
1) Which Cells<br />
2) Which Areas<br />
Inject pontine nuclei with a retrograde tracer; Identify labelled cortical cells<br />
TREE SHREW<br />
MONKEY: SOURCE OF CORTICO PONTINE VISUAL FIBRES<br />
VAN ESSEN EXTRA-STRIATE VISUAL AREAS<br />
NUMBER OF VISUAL AREAS AS A FUNCTION OF TIME<br />
Distiction of parietal <strong>and</strong> temporal lobe directed cortical connections<br />
1) Layer V pyramidal cells.<br />
2) The differential distribution from extrastriate visual areas in the monkey.<br />
(Superior Colliculus; Pretectum <strong>and</strong> LGN)<br />
11
Pontocerebellar Pathway<br />
Cerebellar afferent fibres<br />
Mossy vs. climbing fibres<br />
3 peduncles<br />
HUMAN PONS; SAGITTAL SECTION<br />
MIDDLE PEDUNCLE<br />
middle > (inferior <strong>and</strong> superior)<br />
all (well nearly all) from n. pontis.<br />
Contralateral <strong>and</strong> ipsilateral projection in the middle cerebellar peduncle: Possible solution to<br />
some old problems.<br />
INFERIOR OLIVE (HUMAN)<br />
Climbing fibre input to the cerebellum. (J. Simpson)<br />
Accessory optic system dorsal cap of the inferior olivary nucleus. Flocculus <strong>and</strong> adjacent<br />
flocculus.<br />
On the difference between mossy fibre <strong>and</strong> climbing fibre visual afferents to the cerebellum.<br />
Some behavioural evidence for cerebellar role in visuo motor coordination <strong>and</strong> plasticity <strong>and</strong><br />
some unresolved questions.<br />
1. Plasticity of the VOR<br />
2. Saccadic Adaptation<br />
2. Prism adaptation<br />
3. Nictitating membrane conditioning<br />
References<br />
<strong>Eye</strong> <strong>and</strong> Central Visual pathways<br />
Classic work on the comparative anatomy of the eye is:<br />
Walls,G. (1942) The Vertebrate <strong>Eye</strong> <strong>and</strong> its Adaptive radiation Bloomfield Hills; Cranbrook<br />
A good general source for sensory function<br />
Barlow, H., <strong>and</strong> Mollon, J. (1983) The senses Cambridge<br />
A recent massive reference :<br />
Chalupa, L., <strong>and</strong> Werner, J. (2003) The Visual <strong>Neuroscience</strong>s Cambridge, MIT Press<br />
9:<br />
Dacey DM, Packer OS. (2003), Colour coding in the primate retina: diverse cell types <strong>and</strong><br />
cone-specific circuitry. Curr Opin Neurobiol. 2003 Aug;13(4):421-7. Review.<br />
Functions beginning to catch up with Cajal’s anatomy.<br />
Lamb et. A;.(2007), Evolution of the vertebrate eye: opsins , photoreceptors, retibna <strong>and</strong> eye<br />
cup. Nature <strong>Neuroscience</strong> Reviews; 8 . 960-975.<br />
Evidenvce of evolution beginning to reassure Darwin<br />
For the central visual pathways, any good neuroscience text.<br />
<strong>Eye</strong> Movements<br />
Robinson, D.A. (1981) Control of eye movements in (V. Brooks, Ed.). American<br />
Physiological Society H<strong>and</strong>book of Physiology Section 1. The Nervous System, Volume II<br />
Motor Cortical, Ch. 28, 1275-1320.<br />
Carpenter, R.H.S. (1988) Movements of the <strong>Eye</strong>s. London: Pion.<br />
Lee, R.J., <strong>and</strong> Zee, D.S. (1999) The Neurology of <strong>Eye</strong> Movements New York; Oxford U<br />
12
ENTOPTIC PHENOMENA, VISUAL PHENOMENA AND EYE ANATOMY<br />
The pupil as a first line of defence against excessive light.<br />
DEMONSTRATION 1. PUPIL SHAPE; SIZE CHANGE<br />
Make a triangular shape with the tips of your fingers; bring the triangle closer to your eye:<br />
What happens to the shape of the triangle?<br />
Now open <strong>and</strong> close the other eye. Observe your own (consensual) Pupillary Reflex.<br />
The pupillary reflex has been well studied quantitatively. If we arbitrarily increase gain of<br />
the pupillary reflex.<br />
DEMONSTRATION 2. OSCILLATING PUPIL<br />
Shine penlight just on the edge of the pupil. Pupil will oscillate in size.<br />
In general with poor focus as in uncorrected myopia size of blur circles is related to the size<br />
of pupil. Therefore decreasing pupil size causes improved resolution with sufficient ambient<br />
light.<br />
DEMONSTRATION 3. REFRACTIVE DISORDER/PINHOLE<br />
Chose a myopic subject: Find the far point of distinct vision by seeing how far they can see<br />
the tip of a pin clearly. Place text just beyond far point so as to be unreadable. Now view<br />
through narrow pinhole.<br />
ABBERATIONS AND DIFFRACTION<br />
The pupil plays a complex role in relation to image quality <strong>and</strong> hence to acuity. Three major<br />
physical factors are related to pupil size - can a point of light be imaged as a point on the<br />
retina?<br />
Spherical Aberration<br />
In any simple spherical lens the edges of the lens refract more than the centre. Therefore a<br />
distant point can not be imaged as a point.<br />
(Human cornea is actually somewhat flatter at edge; consider effects of hard contact lenses<br />
with large pupils).<br />
Diffraction<br />
Points of light are actually imaged as concentric bright/dark rings. Diffraction effect is<br />
greater with smaller pupils.<br />
Chromatic Aberration<br />
Short wave length light is refracted more than long wave lengths.<br />
Therefore given a white light, different wavelengths will be imaged at different depths.<br />
DEMONSTRATION 4. BLUE-FREE ZONE IN CENTRAL FOVEA<br />
DIRECTIONALITY OF VISION<br />
Visual world is imaged on the retina as in a camera (left-right; up-down inversion).<br />
Yet we locate directions accurately relative to our position in space.<br />
DEMONSTRATION 5. PRESSURE PHOSPHENES<br />
Press gently with a cotton swab on the cornea of your eye. Can you see a pale flash of light?<br />
Where does it seem to come from?<br />
Schlodtmann's Experiment <strong>and</strong> the innate nature of visual direction.<br />
(Keep in mind the orderly relationship between retinal position <strong>and</strong> visual direction: How is<br />
it that we can adapt to optical disruption of direction such as in inverting or lateral-displacing<br />
prisms?)<br />
RETINAL VESSELS AND THE STABILIZED IMAGE<br />
Retinal receptors are supplied by way of the choroidal circulation <strong>and</strong> the pigment<br />
epithelium.<br />
Inner nuclear <strong>and</strong> ganglion cells by way of central retinal artery.<br />
Why don't you see these vessels since they lie in the path of the light?<br />
13
Probably because their shadows are stabilized on the retina.<br />
Therefore if we illuminate eccentrically, we might see them.<br />
DEMONSTRATION 6. PURKINJE VESSEL FIGURE<br />
Illuminate eye through the sclera with a penlight. It is now possible to see an important point<br />
about the position of retinal vessels relative to the fovea.<br />
DEMONSTRATION 7. AFTER IMAGE IN FOVEA WITH VESSELS<br />
Rods have lower thresholds <strong>and</strong> different spectral sensitivity than cones. Purkinje observed<br />
the relative brightness of the rose <strong>and</strong> its leaves in his garden.<br />
DEMONSTRATION 8. RED AND BLUE: BRIGHTNESS IN DIFFERENT LEVELS<br />
OF ILLUMINATION<br />
SPEED OF RESPONSE<br />
Cones work more rapidly: Activate ganglion cells sooner <strong>and</strong> the message reaches the brain<br />
sooner. Even within the photopic range, lower illumination levels lead to slower response of<br />
receptors <strong>and</strong> ganglion cells.<br />
DEMONSTRATION 9. REACTION-TIME UNDER CONDITIONS OF DIM AND<br />
BRIGHT ILLUMINATION<br />
Two illusions induced by differences in latency of response.<br />
DEMONSTRATION 10. FLUTTERING HEARTS<br />
A more subtle effect can be seen binocularly.<br />
DEMONSTRATION 11. PULFRICH EFFECT<br />
Object moves back <strong>and</strong> forth<br />
Seen as a disparity by the two eyes. Therefore interpreted as depth hence<br />
elliptical orbit.<br />
PERSISTENCE OF ROD AFTER IMAGES<br />
DEMONSTRATION 12. FLASH AFTER 5 MINUTES DARK ADAPTATION<br />
CENTRAL COURSE OF OPTIC FIBRES WITHIN RETINA<br />
Ganglion cells fires <strong>and</strong> axon conducts centrally. First along<br />
retinal surface <strong>and</strong> thence, through to the brain.<br />
Transmission via unmyelinated optic nerve fibres.<br />
DEMONSTRATION 13. BLUE ARCS<br />
(1) Why is it shaped as it is?<br />
(2) Why coloured blue?<br />
STABILITY OF THE VISUAL WORLD: EFFERENCE COPY AND THE ILLUSION OF<br />
MOVEMENT<br />
When we sweep our eyes over the scene in front of us it appears to remain stable. Why? Not<br />
necessarily because it is stable.<br />
DEMONSTRATION 14. JIGGLING YOUR EYE WORLD MOVES<br />
Helmholtz Interpretation: Signal from brain nulls apparent movements of the world when we<br />
voluntarily move our eyes.<br />
(Note Not brought into play during vestibular induced movements).<br />
THERE EXIST A PARALLEL SET OF EXPERIMENTS WHICH REINFORCE THE<br />
VIEW<br />
Suppose you have a stable image on your retina <strong>and</strong> will the eyes to move. What happens?<br />
Conversely, if you jiggle your eye, the after-image remains stable.<br />
DEMONSTRATION 15. ACTIVE AND PASSIVE MOVEMENT OF THE EYE WITH<br />
AN AFTER IMAGE ON FOVEA<br />
EYE MOVEMENTS AND VISUAL CONTROL OF MOVEMENT: CHANGING THE<br />
14
IMAGE AND THE MODIFICATION OF VISUALLY INDUCED RESPONSES<br />
The function of vision is to guide movement. Either the whole body, individual limbs, or the<br />
eyes themselves.<br />
Some systematics about movements of the eye.<br />
DEMONSTRATION 16. SMOOTH PURSUIT VS. SACCADIC MOVEMENT<br />
Observe your partner 1) following a smoothly moving target<br />
2) trying to replicate that movement<br />
Only saccade is under voluntary control.<br />
DEMONSTRATION 17. ACCOMMODATION AND VERGENCE MOVEMENT<br />
(pupil size decreases)<br />
Ask subject to view a distant target: observe eyes <strong>and</strong> pupil as you bring it closer to their<br />
face.<br />
VESTIBULO-OCULAR REFLEX<br />
DEMONSTRATION 18. SHAKE HAND WITH HEAD STILL; SHAKE HEAD WITH<br />
HAND STILL<br />
also READ TEXT WHILE SHAKING HEAD OR BODY<br />
LATERALLY DISPLACING PRISM<br />
DEMONSTRATION 19.<br />
Point at target Adapt: Mispoint when prism removed.<br />
Argument that effect is Not Visual (1) Doesn't transfer between arms<br />
(2) Transfer to pointing to a sound source<br />
Vision in conflict with propioception.<br />
DEMONSTRATION 20. VISUAL CAPTURE<br />
15