

Autofluorescent Proteins with Excitation in the Optical ... - Tsien
Autofluorescent Proteins with Excitation in the Optical ... - Tsien
Autofluorescent Proteins with Excitation in the Optical ... - Tsien

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
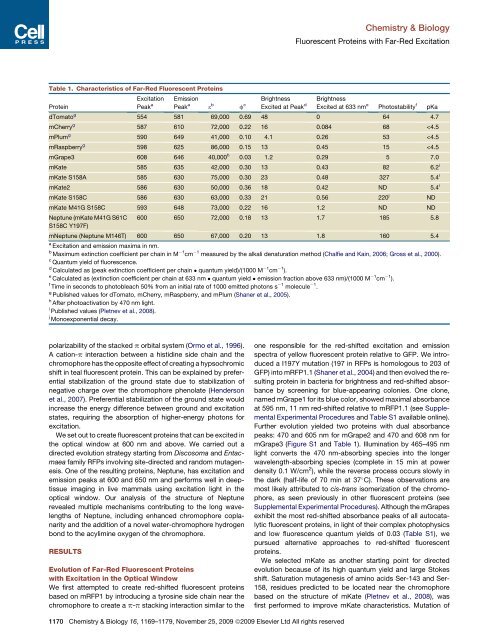
Table 1. Characteristics of Far-Red Fluorescent <strong>Prote<strong>in</strong>s</strong><br />
polarizability of <strong>the</strong> stacked p orbital system (Ormo et al., 1996).<br />
A cation-p <strong>in</strong>teraction between a histid<strong>in</strong>e side cha<strong>in</strong> and <strong>the</strong><br />
chromophore has <strong>the</strong> opposite effect of creat<strong>in</strong>g a hypsochromic<br />
shift <strong>in</strong> teal fluorescent prote<strong>in</strong>. This can be expla<strong>in</strong>ed by preferential<br />
stabilization of <strong>the</strong> ground state due to stabilization of<br />
negative charge over <strong>the</strong> chromophore phenolate (Henderson<br />
et al., 2007). Preferential stabilization of <strong>the</strong> ground state would<br />
<strong>in</strong>crease <strong>the</strong> energy difference between ground and excitation<br />
states, requir<strong>in</strong>g <strong>the</strong> absorption of higher-energy photons for<br />
excitation.<br />
We set out to create fluorescent prote<strong>in</strong>s that can be excited <strong>in</strong><br />
<strong>the</strong> optical w<strong>in</strong>dow at 600 nm and above. We carried out a<br />
directed evolution strategy start<strong>in</strong>g from Discosoma and Entacmaea<br />
family RFPs <strong>in</strong>volv<strong>in</strong>g site-directed and random mutagenesis.<br />
One of <strong>the</strong> result<strong>in</strong>g prote<strong>in</strong>s, Neptune, has excitation and<br />
emission peaks at 600 and 650 nm and performs well <strong>in</strong> deeptissue<br />
imag<strong>in</strong>g <strong>in</strong> live mammals us<strong>in</strong>g excitation light <strong>in</strong> <strong>the</strong><br />
optical w<strong>in</strong>dow. Our analysis of <strong>the</strong> structure of Neptune<br />
revealed multiple mechanisms contribut<strong>in</strong>g to <strong>the</strong> long wavelengths<br />
of Neptune, <strong>in</strong>clud<strong>in</strong>g enhanced chromophore coplanarity<br />
and <strong>the</strong> addition of a novel water-chromophore hydrogen<br />
bond to <strong>the</strong> acylim<strong>in</strong>e oxygen of <strong>the</strong> chromophore.<br />
RESULTS<br />
<strong>Excitation</strong><br />
Peak a<br />
Emission<br />
Peak a<br />
Evolution of Far-Red Fluorescent <strong>Prote<strong>in</strong>s</strong><br />
<strong>with</strong> <strong>Excitation</strong> <strong>in</strong> <strong>the</strong> <strong>Optical</strong> W<strong>in</strong>dow<br />
We first attempted to create red-shifted fluorescent prote<strong>in</strong>s<br />
based on mRFP1 by <strong>in</strong>troduc<strong>in</strong>g a tyros<strong>in</strong>e side cha<strong>in</strong> near <strong>the</strong><br />
chromophore to create a p-p stack<strong>in</strong>g <strong>in</strong>teraction similar to <strong>the</strong><br />
3 b<br />
f c<br />
Brightness<br />
Excited at Peak d<br />
Brightness<br />
Excited at 633 nm e<br />
Prote<strong>in</strong><br />
pKa<br />
dTomato g<br />
554 581 69,000 0.69 48 0 64 4.7<br />
mCherry g<br />
587 610 72,000 0.22 16 0.084 68



