
You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
4. Onkogener og tumorsuppressorer<br />
– Kræftcellernes svar på speeder og bremse<br />
Dette kapitel fortæller,<br />
· hvorfor kræftceller deler sig ukontrolleret<br />
· hvad et onkogen er<br />
· hvad en tumorsuppressor er<br />
· hvad denne viden kan bruges til<br />
En kræftcelle – en slidt bil med speederen i bund<br />
og uden bremse<br />
Kræftcellen bliver ofte sammenliget med en slidt gammel bil,<br />
hvor speederen er trykket i bund, og hvor bremsen mangler.<br />
Med det billede i tankerne forestiller man sig en bil, som<br />
bare bliver ved med at køre af sted og ikke er til at stoppe.<br />
Men før vi hopper med på vognen – for det gør vi – vil vi<br />
præcisere, hvordan vi vil overføre dette billede til en kræftcelle:<br />
En kræftcelle med "speederen i bund" og "uden bremse"<br />
er en celle, som hæmningsløst kaster sig ud i den ene celledeling<br />
efter den anden. Vi vil bruge resten af kapitlet på at<br />
forklare, hvad "speederen" og "bremsen" er i cellesprog.<br />
Først vil vi se på, hvordan speederen reguleres i en normal<br />
celle, og hvordan speederen sætter sig fast i kræftceller.<br />
Dernæst vil vi se, hvordan bremsen reguleres i en normal<br />
celle, og ikke overraskende, hvordan bremsen går i stykker i<br />
kræftceller.<br />
Til sidst vil vi fortælle, hvordan speederen og bremsen i<br />
fællesskab bestemmer, om cellen skal dele sig i to. Alt dette<br />
vil samtidig forklare, hvorfor celledelinger foregår meget<br />
kontrolleret i normale celler, og hvordan kræftceller mister<br />
denne kontrol.<br />
Normale celler bestemmer ikke deres egen skæbne<br />
Historien om normale cellers speedere og bremser starter på<br />
cellens overflade. For det er ikke cellen selv, som bestemmer,<br />
om den skal dele sig eller lade være. Det er derimod<br />
signaler afleveret fra andre celler til cellens overflade, som<br />
bestemmer cellens skæbne. Hvis den overvejende del af signalerne<br />
fortæller cellen, at den skal dele sig, deler cellen sig.<br />
Hvis den overvejende del af signalerne fortæller det modsatte,<br />
nemlig at cellen skal stoppe sin deling, stopper cellen<br />
sin deling.<br />
Det vil sige, at en normal celle er en meget social celle,<br />
som kun gør de ting, den får besked på.<br />
Hvor kommer signalerne fra?<br />
Signalerne kan enten komme fra naboceller i det væv, hvor<br />
cellen befinder sig – parakrin signalering (græsk: para = ved<br />
siden af), eller også fra blodbanen, hvor signalerne kan være<br />
produceret et helt andet sted i kroppen – endokrin signalering<br />
(græsk: endon = indvendig). På den måde indgår cellerne<br />
i et tæt samarbejde, som sørger for, at hvert væv opretholder<br />
sin rette størrelse og en struktur, som er forenelig med<br />
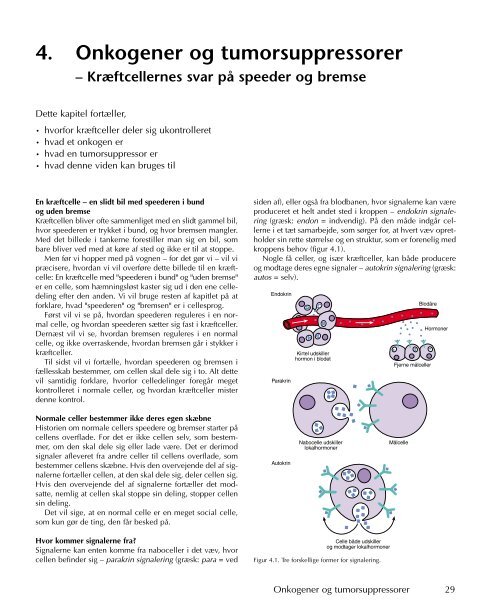
kroppens behov (figur 4.1).<br />
Nogle få celler, og især kræftceller, kan både producere<br />
og modtage deres egne signaler – autokrin signalering (græsk:<br />
autos = selv).<br />
Figur 4.1. Tre forskellige former for signalering.<br />
Onkogener og tumorsuppressorer<br />
29
Ved parakrin og autokrin signalering afgiver en celle et signalmolekyle,<br />
som kun kan påvirke de nærmeste celler – der er<br />
tale om en afstand på under 1 mm. Signalmolekylerne bliver<br />
også kaldt lokalhormoner. Lokalhormoner kommer ikke i kontakt<br />
med blodbanen, men bliver derimod afgivet til det rum,<br />
som er mellem cellerne og kaldes det ekstracellullære rum.<br />
På den måde adskiller lokalhormoner sig fra normale hormoner,<br />
som bruges ved endokrin signalering. Normale hormoner<br />
bliver lavet i kirtler og bagefter afgivet til blodbanen,<br />
hvor hormonerne rejser rundt og påvirker mange celler, som<br />
er fordelt i hele kroppen. Fokus i meget af denne bog vil<br />
være på den enkelte celle og dens nære miljø. Vi vil derfor<br />
fokusere på virkningen af lokalhormoner frem for hormoner,<br />
der påvirker cellerne via den endokrine signalering.<br />
Lokalhormoner er ikke ét bestemt signalmolekyle, som<br />
har én funktion. De er mange forskellige molekyler, som<br />
påvirker deres målceller på vidt forskellige måder. De lokalhormoner,<br />
som får deres målcelle til at dele sig, kaldes<br />
vækstfaktorer. Omvendt vil vi kalde de lokalhormoner, som<br />
bremser en celles deling, for antivækstfaktorer.<br />
Fra vækstfaktor til celledeling<br />
Nu forestiller vi os, at en celle har modtaget mange vækstfaktorer<br />
fra nabocellerne. Vækstfaktorerne binder sig til særlige<br />
receptorer på overfladen af cellen. Receptorer (latin: en<br />
som optager noget) er antennelignende proteiner, som ud over<br />
at have en ydre del, som genkender vækstfaktorerne uden for<br />
30 Bogen om kræft<br />
cellen, også har en indre del, som munder ud i cellens cytoplasma<br />
(græsk: kytos = hulrum, plasma = det formede). Receptorerne<br />
har altså en indre og en ydre del, som er forbundet<br />
med et stykke, der går gennem cellemembranen.<br />
Når vækstfaktorerne bindes til receptorerne, videresender<br />
receptorerne signalerne til andre proteiner i cytoplasmaet.<br />
De proteiner, som ligger tæt på den del af receptorerne, som<br />
er inde i cellen, sender signalerne videre til andre proteiner,<br />
som ligger dybere inde i cellen, og sådan fortsætter det i en<br />
kæde af signaler, som ender i cellens centrum, cellekernen.<br />
Inde i cellekernen svarer en type af proteiner, kaldet transkriptionsfaktorer,<br />
på det in<strong>dk</strong>omne vækstsignal ved at aktivere<br />
en gruppe af gener. Aktiveringen af generne fører til<br />
nydannede proteiner, som sørger for at føre cellen gennem<br />
sin deling.<br />
Proteinerne danner en signalkæde<br />
Proteinerne, som videresender et signal fra receptoren og<br />
ind til cellens kerne, kaldes en signalkæde (figur 4.2). Proteinerne<br />
i signalkæden er i stand til at modtage et signal fra et<br />
protein og sende det videre til et andet. Billedligt kunne det<br />
svare til en lang række af mennesker (proteiner), som i gamle<br />
dage slukkede brande ved at lade en spand fyldt med vand<br />
(signal) gå fra hånd til hånd fra brønden (receptor) til branden<br />
(cellekernen). Signalkæden findes i alle kroppens celler,<br />
men aktiveres kun, når receptorerne modtager vækstfaktorer<br />
fra en anden celle.<br />
Figur 4.2. En signalkæde, som fører til en celles deling. Vækstfaktorer afgivet fra naboceller binder sig til receptorer på målcellens overflade og starter et signal.<br />
Signalet bliver overført fra det ene protein til det andet (en signalkæde) og ender i cellekernen, hvor transkriptionsfaktorer sørger for, at de proteiner,<br />
som skal føre cellen gennem sin deling, bliver dannet.
Signalkædens komponenter<br />
er kendte<br />
De seneste 20 år er kortlægningen af de signalkæder, der<br />
fører til cellens deling, gået meget stærkt. Forskere har<br />
fundet frem til mange af de proteiner, som udgør signalkæderne.<br />
De har også fundet ud af deres rækkefølge.<br />
PDGF – et eksempel på en vækstfaktor<br />
Her følger en beskrivelse af den typiske signalkæde, som<br />
de fleste vækstfaktorer bruger for at få en celle til at dele sig.<br />
Vækstfaktoren PDGF (engelsk: Platelet-Derived Growth<br />
Factor) bruges som eksempel.<br />
Der skal to til en tango<br />
Lad os forestille os en normal målcelle i bindevævet. Den<br />
producerer ikke PDGF. Det kan derimod en anden celle,<br />
som befinder sig i knoglemarven. Den producerende<br />
celle afgiver PDGF til målcellen. PDGF-molekylet er<br />
bundet til et andet PDGF-molekyle. De danner det, man<br />
kalder en dimer. Mange vækstfaktorer virker på denne<br />
måde. Dimeren binder nu ikke til én receptor, men til to,<br />
og det viser sig at være vigtigt. For det medfører, at de to<br />
receptorer bliver tvunget til at ligge fysisk tæt på hinanden.<br />
Herved får de to receptorer en let ændret struktur, og<br />
det aktiverer dem. Der skal to til en tango.<br />
Figur 4.3. Aktivering af PDGF-receptoren. PDGF-dimeren binder til to<br />
receptorer, og det bringer dem i umiddelbar nærhed af hinanden. Den<br />
tætte kontakt giver receptorerne en strukturændring, som gør den ene<br />
receptor i stand til at sætte en fosfatgruppe på den anden receptor og vice<br />
versa. Nu er receptorerne aktive, og de sender et vækstsignal ind i cellen.<br />
PDGF´s receptor – en tyrosinkinase-receptor<br />
Hvordan bliver PDGF´s receptor så aktiv? Receptoren er<br />
en såkaldt tyrosinkinase-receptor. Kinase betyder, at den<br />
kan sætte en fosfatgruppe på et andet protein. I dette tilfælde<br />
sættes fosfatgruppen på aminosyren tyrosin og derfor<br />
navnet tyrosinkinase-receptor. Kinaseaktiviteten sidder<br />
på den del af receptoren, som befinder sig i cellens indre.<br />
Kinasen bliver aktiv, når PDGF-vækstfaktoren binder sig<br />
til receptoren. Strukturændringen af receptorerne gør, at<br />
kinasen bliver aktiv og gør den i stand til at sætte en fosfat-<br />
gruppe på (fosforylere hedder det) den anden receptor. Så<br />
er receptoren aktiv og er blandt andet i stand til at fosforylere<br />
– og dermed aktivere – andre proteiner end sig selv.<br />
Fra receptor til aktive transkriptionsfaktorer<br />
Men det er faktisk ikke ved at fosforylere de nærmeste proteiner<br />
i cytoplasmaet, at den aktive receptor får sat gang<br />
i cellens deling. Et protein, Grb2, som normalt befinder<br />
sig i cellens cytoplasma, bliver tiltrukket af fosfatgruppen<br />
på den aktive receptor og binder sig derefter til receptoren.<br />
Grb2 har ikke nogen enzymaktivitet i sig selv, men er<br />
bundet til et andet protein, Sos, som har en enzymaktivitet.<br />
Ved receptoren kommer Sos i tæt kontakt med det tredje<br />
protein i rækken, Ras, som sidder på indersiden af cellemembranen.<br />
Her aktiveres Ras af Sos.<br />
Figur 4.4. Et vækstsignal, som oprindeligt starter på cellens overflade,<br />
skal igennem mange proteiner, før det når til målet i cellekernen: Fra<br />
PDGF-receptor til Grb2 til Sos til Ras til Raf til MEK til ERK til transkriptionsfaktorer.<br />
De aktive transkriptionsfaktorer sørger for, at nye proteiner<br />
bliver dannet. Disse sørger for, at cellen deler sig.<br />
Onkogener og tumorsuppressorer<br />
31
Aktiv Ras har "en kort levetid" og kræver vækstfaktor<br />
Ras befinder sig enten i en aktiv eller en inaktiv form. Ras<br />
er et guanin-nukleotid-bindende (guanin-diphosphat (GDP)<br />
eller guanin-triphosphat (GTP)) protein. I sin inaktive form<br />
binder Ras GDP, men når vækstreceptoren er aktiv, sørger<br />
Sos for at udskifte GDP-molekylet med GTP, og det gør<br />
Ras aktiv. GTP-Ras-komplekset har en kort levetid, fordi<br />
andre proteiner (såkaldte GAP-proteiner) hurtigt sørger for,<br />
at komplekset bliver omdannet til den inaktive udgave,<br />
hvor GDP er bundet til Ras. Derfor kræver det, at vækstfaktoren<br />
hele tiden er bundet til vækstreceptoren, hvis<br />
Ras skal forblive aktiv.<br />
Fra Ras til transkriptionsfaktorer: en kaskade af fosforyleringer<br />
Det aktive Ras-protein sørger for at aktivere det fjerde<br />
protein i rækken, nemlig proteinkinasen Raf. Aktiveringen<br />
af Raf er startskuddet til en kaskade af fosforyleringer,<br />
hvor den ene proteinkinase (Raf) fosforylerer den anden<br />
(MEK) – den anden (MEK) den tredje (ERK) – og det ender<br />
med, at en fosforyleret og aktiv proteinkinase (ERK) bevæger<br />
sig fra cytoplasmaet og ind i cellekernen, hvor proteinet<br />
(ERK) fosforylerer og aktiverer indtil flere DNA-bindende<br />
proteiner (transkriptionsfaktorer), som stimulerer<br />
transkription: De sørger for at aktivere de gener, som er<br />
nødvendige for celledelingen. De nydannede proteiner<br />
tager så over og sørger for at gøre cellen klar til det, det<br />
hele handler om: cellens deling (kapitel 5).<br />
I normale celler "kan man lette foden fra speederen"<br />
Normale cellers delinger er nøje reguleret af vækstfaktorerne.<br />
Hvis den overvejende del af de signaler, som cellerne<br />
modtager, indeholder vækstfaktorer, vil cellen dele sig. Hvis<br />
vækstfaktorerne forsvinder, vil cellen holde op med at dele<br />
sig.<br />
Hvis vi kort vender tilbage til bilen, kunne vækstfaktoren<br />
svare til en fod, og receptoren kunne være speederen. Hvis<br />
vækstfaktoren er til stede og binder til receptoren, vil det<br />
svare til, at foden trykker på speederen, og det får bilen til at<br />
køre (cellen deler sig). Omvendt: Hvis vækstfaktoren forsvinder,<br />
vil det svare til, at foden løftes fra speederen – bilen<br />
stopper (cellen deler sig ikke).<br />
Kræftceller sætter sig ud over nabocellernes kontrol<br />
Normale celler deler sig altså kun, når de får besked på det.<br />
Her adskiller kræftceller sig fra normale celler. For kræftceller<br />
er i stand til at dele sig, selvom de ikke har fået besked<br />
på det. Det vil sige, at kræftceller kan dele sig, selvom de<br />
ikke har modtaget vækstfaktorer fra nabocellerne. På den<br />
måde sætter kræftcellerne sig ud over nabocellernes kontrol<br />
af celledelingerne. At sætte sig ud over nabocellernes kontrol<br />
er det samme som at sætte sig ud over kroppens kontrol.<br />
Og det kan være livsfarligt. Speederen har sat sig fast.<br />
32 Bogen om kræft<br />
Hvad er det, fosfat gør?<br />
Det er gennemgående for denne og andre signalkæder, at<br />
et protein bliver aktivt, når det modtager en fosfat fra et<br />
andet protein. Hvis en inaktiv kinase bliver fosforyleret,<br />
bliver den aktiv og i stand til at fosforylere andre proteiner.<br />
Hvis en transkriptionsfaktor bliver fosforyleret, gør<br />
det proteinet i stand til at binde til DNA og aktivere<br />
genet.<br />
Hvad skyldes det? Fosfat, som er stærkt negativt ladet,<br />
får proteinet til at ændre sin struktur. Den aktive del af<br />
enzymet, som normalt ligger skjult i proteinets indre, bliver<br />
blottet og dermed i stand til at udføre sin aktivitet. For<br />
eksempel består en transkriptionsfaktor af en DNA-bindende<br />
del og en regulerende del. Når fosfatgruppen bliver<br />
bundet til den regulerende del, sker der en strukturændring,<br />
som blotter den DNA-bindende del, som herefter<br />
binder sig til DNA'et.<br />
Figur 4.5. Fosforylering gør transkriptionsfaktor aktiv (blå garnnøgle).<br />
Fosforyleringen ændrer transkriptionsfaktorens struktur, så den ellers<br />
gemte DNA-bindende del blotlægges og binder til DNA.<br />
Signalkæden er forstyrret i kræftceller<br />
Hvordan kan det være, at kræftceller, som før har været normale<br />
celler, pludselig gør sig uafhængige af nabocellerne og<br />
deler sig uhæmmet? Det skyldes, at den signalkæde, som<br />
normalt styrer en celles deling, er forstyrret i kræftcellerne. I<br />
en normal celle bliver signalkæden kun "tændt", når vækstfaktorerne<br />
binder sig til receptorerne. I kræftceller derimod<br />
er signalkæden tændt hele tiden, og det uanset om der er<br />
vækstfaktorer til stede eller ej.<br />
Det kan gå galt flere steder i signalkæden<br />
En normal celles udvikling til kræftcelle kan skyldes skader,<br />
som opstår forskellige steder i signalkæden. Nogle kræftceller<br />
har f.eks. en defekt receptor, som hele tiden sender et vækstsignal<br />
ind i cellen, selvom receptoren ikke har modtaget en<br />
vækstfaktor. I andre kræftceller har indre signalproteiner fået<br />
en defekt, som gør, at de hele tiden udsender et vækstsignal.<br />
Det betyder i begge tilfælde, at cellen vil dele sig, selvom en<br />
vækstfaktor ikke har bundet sig til kræftcellens receptorer.<br />
Hvordan kan det gå så galt?<br />
For at svare på det må man først se på, hvordan en normal<br />
celle "bygger" den signalkæde, som styrer cellens deling.<br />
Signalproteinerne, der udgør signalkæden, bliver dannet ud
fra få af cellens gener. Det kan få fatale følger, hvis en celle<br />
er så uheldig, at der tilfældigt opstår en fejl – en mutation –<br />
i et af disse få gener.<br />
En mutation er startskuddet til kræft<br />
Ved mutationer kan der ske flere ting (kapitel 2). Ofte har<br />
mutationen ingen betydning, men hvis mutationen rammer<br />
genet særlig uheldigt, kan der ske en af to ting: Enten kan<br />
mutationen medføre, at kontrollen af genets aflæsning (transkription)<br />
forsvinder. Det betyder, at cellen danner alt for<br />
meget af det vækststimulerende protein. Eller også kan en<br />
mutation i genet føre til et ændret protein. Denne ændring<br />
kan medføre, at proteinet konstant udsender et vækstsignal.<br />
Begge former for fejl – alt for meget vækststimulerende protein<br />
eller en hyperaktiv udgave af vækstproteinet – kan<br />
ændre cellens opførsel. Det kan måske være startskuddet til,<br />
at en normal celle udvikler sig til en kræftcelle.<br />
Proto-onkogener bliver til onkogener<br />
Potentielt kan fejl i ét af de relativt få gener, der styrer cellens<br />
deling, være kræftfremkaldende. Men man ved ikke, om det<br />
gælder for dem alle. Den del af generne, som har vist sig at<br />
være kræftfremkaldende, når de får en uheldig mutation, kalder<br />
man proto-onkogener (græsk: proto = først/forrest, onkos<br />
= masse).<br />
Proto-onkogenerne adskiller sig ikke fra de øvrige gener,<br />
der er involveret i celledeling, og er derfor en nødvendig og<br />
naturlig del af cellens liv.<br />
Men hvis der opstår en fejl, en mutation, i et proto-onkogen,<br />
kan det føre til, at cellen deler sig ukontrolleret. Når et protoonkogen<br />
får en mutation, som gør det kræftfremkaldende,<br />
ændrer genet navn til et onkogen. Her følger nogle eksempler.<br />
Nogle kræftceller forsyner sig selv med vækstfaktorer<br />
Normalt undgår vækstfaktorproducerende celler at stimulere<br />
sig selv, fordi de ikke har de receptorer, som er nødvendige<br />
for at modtage vækstfaktorerne (figur 4.6).<br />
Omvendt er det med de celler, som modtager vækstfaktorerne<br />
og deler sig. De har kun receptorerne og ikke vækstfaktorerne.<br />
Det vil sige, at de kun deler sig, når de modtager<br />
vækstfaktorer fra andre celler. På den måde indgår cellerne<br />
i et fint samarbejde, hvor de er afhængige af hinanden (parakrin<br />
signalering).<br />
Det er netop denne fine balance, der bliver forstyrret i<br />
nogle kræftceller. De er blevet ramt af særlige mutationer i<br />
nogle proto-onkogener, som gør cellerne i stand til at producere<br />
deres egne vækstfaktorer. Det sker f.eks. ved sarkomer-<br />
og bindevævskræft. Normalt deler bindevævsceller og<br />
hjerneceller sig kun, når de modtager vækstfaktorer fra naboceller.<br />
Men i kræftudgaven af disse celler har proto-onkogenerne,<br />
som koder for vækstfaktorerne, fået nogle uheldige<br />
mutationer, som gør dem i stand til at producere deres egne<br />
vækstfaktorer. På den måde vil kræftcellerne konstant modtage<br />
signal til at dele sig.<br />
Andre kræftceller deler sig uden vækstfaktor<br />
Men kræftceller kan også sætte sig uden for naboernes kontrol<br />
ved at "lade som om", at de har modtaget vækstfaktorer<br />
fra nabocellerne.<br />
Konstant aktive receptorer<br />
Lad os starte på cellens overflade. Vækstreceptorerne er normalt<br />
kun aktive, når de modtager en vækstfaktor fra nabocellen.<br />
Men i mange kræftformer fyrer receptoren konstant et signal<br />
ind i cellen, selvom den ikke har bundet en vækstfaktor.<br />
For meget vækstreceptor<br />
Epidermal growth factor receptor (EGF-R) er et eksempel på<br />
en receptor, som er overudtrykt i mave-, hjerne- og brystkræft.<br />
Det vil sige, at kræftcellerne indeholder mange flere<br />
EGF-receptorer på deres overflade end normale celler. EGFreceptoren,<br />
som findes i mange forskellige celletyper, får cellerne<br />
til at dele sig, når de modtager deres specifikke vækstfaktor<br />
(EGF-vækstfaktor). Den unaturligt store mængde af receptorer<br />
på kræftcellernes overflade kan have flere følger.<br />
For eksempel gør det cellen særlig følsom over for få vækstfaktorer.<br />
Det vil sige, at cellen deler sig, bare den ser "skyggen"<br />
af en vækstfaktor, som i en normal situation ikke ville<br />
få en celle til at dele sig (figur 4.7).<br />
Mere overraskende er det måske, at det store antal receptorer<br />
også gør dem i stand til at aktivere sig selv spontant<br />
uden tilstedeværelsen af vækstfaktorer (figur 4.8). Det vil sige,<br />
at receptoren bliver ved med at sende et vækstsignal ind i<br />
cellerne, selvom ingen har bedt den om det. Hvordan kan<br />
det lade sig gøre? Den mest almindelige forklaring er, at det<br />
store antal receptorer på cellens overflade øger risikoen for,<br />
at to receptorer støder sammen. Dette sammenstød vil så<br />
ligne almindelig receptoraktivering: Vækstfaktoren bringer<br />
to receptorer i umiddelbar nærhed af hinanden, som øjeblikkelig<br />
gør receptoren aktiv.<br />
Figur 4.6. Normale celler deler sig kun, når de får<br />
besked på det fra andre celler. Nogle kræftceller kan<br />
både producere og modtage egne vækstfaktorer.<br />
Kræftcellen kan give sig selv besked på, at den skal<br />
dele sig.<br />
Onkogener og tumorsuppressorer<br />
33
Figur 4.7. En normal celle deler sig ikke, hvis der kun er lidt vækstfaktor til<br />
stede. Nogle kræftceller har unaturligt mange receptorer, som gør dem i<br />
stand til at "fange" selv små mængder af vækstfaktor.<br />
Figur 4.8. Nogle kræftceller overudtrykker receptorer for vækstfaktorer. De<br />
mange receptorer på kræftcellers overflade aktiveres spontant og uden tilstedeværelse<br />
af vækstfaktorer.<br />
Onkogener kan også kode for defekte proteiner<br />
Den føromtalte EGF-receptor findes også i en anden kræftfremkaldende<br />
udgave. Denne receptor har mistet den ydre<br />
del af receptoren, som normalt findes på cellens overflade.<br />
Det gør receptoren ude af stand til at genkende og binde<br />
vækstfaktoren. Umiddelbart skulle man tro, at det ville give<br />
34 Bogen om kræft<br />
anledning til en inaktiv receptor, som ikke kan udsende et<br />
vækstsignal. Men receptoren er langtfra inaktiv. For manglen<br />
på receptorens ydre del får spontant den tilbageblevne del af<br />
receptoren til at forbinde sig med en anden receptor, som<br />
også mangler yderdelen. Denne ændring betyder, at den<br />
indre del af receptoren begynder at udsende et konstant<br />
vækstsignal (figur 4.9).<br />
En deletionsmutation<br />
Hvordan kan EGF-receptoren miste den del, som genkender<br />
vækstsignalet? Hvis man ser på onkogen-udgaven af EGFreceptoren,<br />
mangler den et stort stykke af det DNA, som i<br />
proto-onkogen-udgaven koder for den ydre del af receptoren.<br />
En sådan mutation kaldes en deletionsmutation. Når<br />
DNA-koden bliver omsat til protein, vil den ydre del af receptoren<br />
mangle, og det giver sig udslag i en konstant aktiv<br />
receptor.<br />
Ras amok i cellens indre<br />
Det er ikke kun på cellens overflade, kræftceller undgår kontrol.<br />
Signalkæden kan også være forstyrret i cellens indre. Et<br />
eksempel er Ras-onkogenet.<br />
Proteinet, som det normale Ras-gen (proto-onkogenet) koder<br />
for, befinder sig i cellens cytoplasma, hvor det er hæftet<br />
fast på indersiden af cellens membran ved hjælp af fedtstofmolekyler.<br />
Ras-proteinerne modtager signalerne, som kommer<br />
ind fra cellens receptorer og sender dem videre til andre<br />
proteiner, som ligger længere nede i signalkæden (figur 4.4,<br />
side 31 og 4.10, side 35).<br />
Men proteinet, som det muterede Ras-gen (onkogener)<br />
koder for, er en strukturelt ændret udgave af Ras-proteinet,<br />
som bliver ved med at sende et signal dybere ind i cellen,<br />
selvom receptorerne ikke har modtaget en vækstfaktor. Det<br />
Figur 4.9. EGF-receptoren får<br />
kun normale celler til at dele<br />
sig, hvis de binder EGF. Nogle<br />
kræftceller har en defekt EGFreceptor,<br />
som hele tiden fortæller,<br />
at kræftcellen skal dele sig.<br />
Og det uanset om der er vækstfaktor<br />
til stede eller ej. Læg<br />
mærke til, at kræftcellen mangler<br />
et stort stykke DNA (deletionsmutation).<br />
Det omdanner et<br />
proto-onkogen til et onkogen.
vil sige, at kræftcellen på sin overflade kan ligne en normal<br />
celle med normale velfungerende receptorer, mens det inde<br />
i cellen står anderledes til. Her findes der et protein, som bliver<br />
ved med at fortælle, at cellen skal dele sig, selvom cellen<br />
ikke har fået besked på det udefra.<br />
Man har fundet disse hyperaktive udgaver af Ras, som<br />
ikke er til at stoppe, i omkring en fjerdedel af alle kræfttilfælde.<br />
I en af de mest velstuderede kræftformer – tyktarmskræft<br />
– har hver anden patient en muteret udgave af<br />
proto-onkogenet i deres kræftceller, og det understreger<br />
genets centrale rolle i kræftudviklingen. Mange kræftforskere<br />
tror, at patienter med samme sygdom, men som ikke har en<br />
Ras-mutation, må have mutationer i de proteiner, som Ras<br />
enten modtager vækstsignalet fra eller sender vækstsignalet<br />
videre til. Det vil i sidste ende give samme resultat. Kræftcellerne<br />
deler sig ukontrolleret.<br />
Det kan også gå galt i cellekernen.<br />
Det er ikke kun i cellens cytoplasma, hyperaktive proteiner<br />
får kræftceller til at dele sig ukontrolleret. Det er de såkaldte<br />
Myc-onkogener et bevis på. Myc-onkogener koder for<br />
transkriptionsfaktorer, som normalt kun dannes på en vækstfaktors<br />
kommando. Transkriptionsfaktorerne sørger for at<br />
aktivere de gener, som er med til at drive cellen frem til sin<br />
deling. Men i mange typer af kræft er niveauet af Myc-transkriptionsfaktorer<br />
hele tiden meget højt – selv i fravær af<br />
vækstfaktorer. Det vil sige, at Myc-transkriptionsfaktorerne<br />
sørger for, at der hele tiden bliver produceret proteiner, som<br />
sørger for, at cellen deler sig, og det selvom ingen har bedt<br />
dem om det (figur 4.11).<br />
Onkogener og tumorsuppressorer<br />
Figur 4.10. Hvis en normal<br />
celle modtager en vækstfaktor,<br />
går Ras-proteinet fra en inaktiv<br />
til en aktiv form (fra Ras-GDP til<br />
Ras-GTP) og sender på den<br />
måde vækstsignalet dybere ind<br />
i cellen. I en fjerdedel af alle<br />
kræfttilfælde findes Ras-proteinet<br />
konstant i sin aktive form<br />
(Ras-GTP) og udsender derfor et<br />
vækstsignal, uanset om der er<br />
vækstfaktor til stede eller ej.<br />
Figur 4.11. Myc-transkriptionsfaktorer<br />
dannes kun på en<br />
vækstfaktors kommando, og<br />
transkriptionsfaktorerne sørger<br />
herefter for, at de proteiner, som<br />
skal føre cellen gennem sin<br />
deling, bliver produceret. I<br />
mange kræftceller er Myc-transkriptionsfaktorerne<br />
til stede<br />
hele tiden, uanset om der er<br />
vækstfaktor til stede eller ej.<br />
Det betyder også, at der konstant<br />
vil være en produktion af<br />
de proteiner, som fører cellen<br />
gennem sin deling.<br />
35
Hvordan finder man<br />
et onkogen?<br />
Normale celler i en cellekultur i laboratoriet er meget<br />
afhængige af at vokse på en fast overflade som f.eks. den,<br />
man ser i petriskåle, mens kræftceller i kultur ikke behøver<br />
noget at binde sig til for at kunne dele sig. Når normale<br />
celler kommer i kontakt med hinanden, holder de<br />
op med at dele sig. Det gør kræftceller ikke. De fortsætter<br />
med at dele sig, og er der ikke plads ved siden af,<br />
begynder de at vokse oven på hinanden (figur 4.12).<br />
Figur 4.12. To petriskåle set fra siden. Den øverste indeholder normale<br />
celler: De vokser kun i et enkelt lag og stopper med at dele sig, hvis de<br />
kommer i kontakt med hinanden (kontakthæmning). Den nederste indeholder<br />
kræftceller: De kan vokse i flere lag (kræver ikke fast overflade),<br />
og de holder ikke op med at dele sig, selvom de kommer i kontakt med<br />
hinanden (undgår kontakthæmning).<br />
Denne forskel satte en amerikansk forskergruppe sig for at<br />
udnytte i deres søgen efter onkogener. De håbede, at de<br />
måske kunne få nogle af de normale celler til at opføre sig<br />
som kræftceller, hvis de overførte gener fra kræftceller til<br />
normale celler. Sagt med andre ord: De håbede, at den normale<br />
celle ville få kræftlignende egenskaber, hvis den modtog<br />
et onkogen fra kræftcellerne. Når en normal celle bliver<br />
til en kræftcelle, siger man, at den transformerer (figur 4.13).<br />
En næsten normal celle<br />
Nu var ideen skabt. Som celler brugte forskerne særlige<br />
fibroblaster (latin: fibra = fiber, græsk: blastos = kim (celler,<br />
som danner bindevæv)) fra mus, som i kultur opfører<br />
sig som normale celler med en undtagelse: De kunne dele<br />
sig uendeligt. Man siger, at de er immortaliserede, gjort<br />
udødelige.<br />
DNA fra kræftsvulst til immortaliserede celler<br />
Med disse celler i petriskålen gik forskerne videre i deres<br />
forsøgsrække. Deres næste skridt var at isolere DNA fra<br />
en menneskelig kræftsvulst, og med en særlig teknik<br />
overførte de DNA'et til de immortaliserede celler i petriskålene.<br />
Efter at være opbevaret et par dage ved 37° C<br />
kiggede forskerne igen på cellerne.<br />
36 Bogen om kræft<br />
Figur 4.13. Sådan kan man finde et onkogen.<br />
Enkelte celler transformerer<br />
Ved første øjekast så det ud, som om der ikke var sket<br />
noget. Men da forskerne så nærmere efter, var der steder,<br />
hvor celler var begyndt at vokse i flere lag, og disse celler<br />
blev med tiden til store synlige kolonier af mange celler.<br />
Enkelte af de immortaliserede celler havde altså fået kræftlignende<br />
egenskaber.<br />
Som at finde en nål i en høstak?<br />
Efter at have gravet en koloni af celler ud af petriskålen<br />
var forskerne selvfølgelig interesseret i at vide, hvad det<br />
var for et stykke DNA, cellerne havde modtaget. Men<br />
hvordan skulle man kunne adskille og finde frem til det<br />
"fremmede" stykke DNA blandt musecellernes eget DNA?<br />
Umiddelbart kunne det virke som at skulle finde en nål i<br />
en høstak. Men her var forskerne snedige.
"Nålen" var et enkelt gen<br />
I mus og menneskers DNA er der nemlig en bestemt<br />
sekvens, som går igen og igen med korte mellemrum, og<br />
som ikke koder for et protein (sekvenserne er nemlig ikke<br />
gener). Sekvensen i mus adskiller sig fra sekvensen i mennesket,<br />
og ved hjælp af en elegant teknik kunne de amerikanske<br />
forskere isolere det humane DNA. De fandt ud af, at<br />
det var et enkelt gen i det DNA, de havde isoleret, som var<br />
i stand til at gøre de immortaliserede celler til kræftceller.<br />
Det er Ras<br />
Det "transformerende" gen var isoleret, og så stod der tilbage<br />
at finde ud af, hvad det var for et. De fandt ud af<br />
genets nukleotidsekvens. Sekvensen fortalte, at det var<br />
genet for Ras-protein, som vi har beskrevet tidligere.<br />
En lille ændring med store konsekvenser<br />
Ras-genet, som stammede fra kræftcellerne, var stort set<br />
identisk med Ras-genet i normale celler. Med én undtagelse:<br />
Nukleinsyre nr. 35 var i stedet for at være guanin<br />
(G) udskiftet med thymin (T). Man kalder det en punktmutation,<br />
når en nukleinsyre er udskiftet med en anden<br />
(figur 4.14).<br />
Denne lille ændring havde store konsekvenser for genets<br />
egenskab. Hvis forskere tog den normale udgave af<br />
genet (proto-onkogenet) ud fra en normal celle og "puttede"<br />
det ind i en ny normal celle, ville cellen ikke blive til en<br />
kræftcelle. Men hvis de tog den ændrede udgave af genet<br />
(onkogenet) fra kræftcellerne, ville den immortaliserede<br />
celle blive til en kræftcelle. Og det kun på grund af en<br />
lille ændring. Hvorfor?<br />
Punktmutationen gør Ras konstant aktiv<br />
Punktmutationen, hvor et G blev udskiftet med et T, medfører,<br />
at codonet, som normalt koder for aminosyren glycin,<br />
bliver til aminosyren valin. Den senere tids forskning<br />
har vist, at det får fatale følger for Ras-proteinets aktivitet.<br />
Normalt er Ras skiftevis inaktiv eller aktiv alt efter, om<br />
proteinet binder GDP (inaktiv) eller GTP (aktiv). Men<br />
med den ene ændrede aminosyre vil Ras forblive i den<br />
aktive form, hvor den binder GTP. Det vil sige, at Ras<br />
konstant sender et vækstsignal ind i cellen (se figur 4.10).<br />
Ikke kun et "uden for kroppen"-fænomen<br />
Selvom det er meget praktisk at arbejde med normale<br />
celler og kræftceller uden for kroppen, er det vigtigt at<br />
undersøge, om det samme sker inde i kroppen. De amerikanske<br />
forskere brugte mus for at se, om deres elegante<br />
metode også ville give et lignende resultat i en levende<br />
organisme. Hvis de puttede de immortaliserede celler ind<br />
i musen – det kalder man at inokulere (latin: in = ind i,<br />
oculus = øje) eller at pode – fandt de ingen ændring af musens<br />
tilstand. Men hvis de derimod podede musen med<br />
immortaliserede celler transformeret med Ras (den onkogene<br />
udgave), så de, at musen udviklede kræftsvulster<br />
der, hvor de havde sprøjtet cellerne ind.<br />
Det viser, at der er en sammenhæng mellem det, man ser<br />
uden for kroppen (in vitro – latin for: i glas), og det, man ser<br />
inde i kroppen (in vivo – latin for: i det levende). Onkogenet<br />
(Ras i dette eksempel) giver immortaliserede celler kræftlignende<br />
egenskaber in vitro. Hvis man putter disse celler ind<br />
i mus, dannes der kræftsvulster. Det er vigtigt altid at sammenholde<br />
de resultater, man får uden for kroppen (in vitro),<br />
med, hvad der sker i et levende system (in vivo).<br />
En helt normal celle vil gå i stå<br />
De immortaliserede celler, som amerikanerne brugte, var<br />
ikke helt normale – de var nemlig udødelige. De vil være<br />
i stand til at dele sig i det uendelige, hvis bare man sørger<br />
for, at der er plads, og de ikke kommer i kontakt med<br />
hinanden.<br />
Denne ændring har vist sig at være vigtig for at finde<br />
et onkogen. Hvis man putter et Ras-onkogen ind i en helt<br />
normal celle, ser man noget andet. I stedet for at dele sig<br />
uhæmmet går cellen i stå. Derfor kan vi fastslå, at et onkogen<br />
alene ikke er nok til at give cellen kræftfremkaldende<br />
egenskaber. Konklusionen er, at der skal mere end<br />
én ændring til at gøre en normal celle til en kræftcelle.<br />
Figur 4.14. Ras-genet (onkogenet), som forskerne fandt i kræftceller, var stort set identisk med Ras-genet (proto-onkogenet) i normale celler med én<br />
undtagelse: Nukleinsyre nr. 35 var i stedet for at være guanin (G) udskiftet med thymin (T). Det får den konsekvens, at når genet oversættes til protein,<br />
er aminosyre nr. 12, glycin, udskiftet til fordel for valin. Den lille ændring får fatale følger. Ras-proteinet vil konstant være aktivt (kan ikke inaktiveres)<br />
og vil konstant udsende et vækstsignal.<br />
Onkogener og tumorsuppressorer<br />
37
"Det er ikke kun speederen, der bestemmer<br />
bilens hastighed"<br />
Indtil nu har vi fokuseret på cellens normale "speeder" –<br />
proto-onkogenet – eller "speederen, der har sat sig fast" –<br />
onkogenet – som den eneste faktor, der bestemmer, om cellen<br />
skal dele sig eller lade være. Oversætter man det til bilsprog,<br />
svarer det til, at det kun er bilens speeder, som bestemmer,<br />
om bilen skal køre eller stoppe. Men ligesom i en<br />
bil findes der også en "bremse" i cellerne, som kan få cellerne<br />
til at stoppe delingerne, selvom speederen er trykket i bund.<br />
Så for at cellerne skal kunne dele sig ukontrolleret, skal de<br />
også af med bremsen.<br />
Antivækstfaktorer sætter også gang i en signalkæde<br />
Når man kigger ned på en bils speeder og bremse, kan man<br />
godt sige, at de ligner hinanden. Men trykker man på dem<br />
hver især, er virkningen vidt forskellig. Det billede kan man<br />
også overføre til en celle. For ligesom vækstsignalet starter<br />
med, at vækstfaktorer binder til receptorer på cellens overflade,<br />
gør antivækstfaktorer det samme. Når antivækstfaktorer<br />
bliver afgivet fra en nabocelle, binder de sig til særlige<br />
receptorer (antivækstreceptorer) på cellens overflade. Denne<br />
binding aktiverer receptorerne og sender antivækstsignalet<br />
videre gennem en kæde af andre proteiner. Antivækstsignalet<br />
ender, ligesom vækstsignalet gør det, i cellekernen,<br />
hvor nye proteiner bliver dannet. Men i stedet for at få cellen<br />
til at dele sig sørger de nye proteiner for, at cellen stopper<br />
sin deling.<br />
En mutation er ikke nok<br />
Det vil sige, at selvom celler har fået en uheldig mutation,<br />
som omdanner et proto-onkogen til et onkogen (speederen<br />
sætter sig fast), vil antivækstfaktorer stadig være i stand til at<br />
stoppe cellens ukontrollable delinger. For at opnå evnen til<br />
at dele sig ukontrolleret skal cellerne også undgå denne barriere.<br />
Nu er det ikke sådan, at cellerne er snedige og udtænker<br />
en fiks måde at gøre det på. For celler tænker ikke, og de har<br />
heller ikke nogen vilje. Nej, det er endnu en uheldig mutation<br />
i en af de celler, som med tiden er blevet til flere og i<br />
forvejen har en mutation i et proto-onkogen, som gør, at antivækstsignalet<br />
bliver sat ud af kraft.<br />
En ny tilfældig mutation ødelægger bremsen<br />
Hvordan kan det ske? Det sker ved, at en tilfældig mutation<br />
rammer et af de gener (tumorsuppressorgener), som koder<br />
for et protein, der er en del af antivækstsignalkæden. Mutationen<br />
ødelægger proteinets funktion eller forhindrer, at det<br />
bliver dannet. Det vil sige, at proteinet ikke kan modtage<br />
eller videresende antivækstsignalet, og derfor er det ude af<br />
stand til at stoppe cellens deling. Populært kunne man sige,<br />
at cellen har mistet sin bremse og nu kan dele sig uhæmmet.<br />
"Svulstundertrykkere"<br />
Det er ikke ét bestemt gen, som skal udsættes for en øde-<br />
38 Bogen om kræft<br />
læggende mutation, for at en celle mister sin bremse. Der er<br />
mange gener, som med særlige mutationer fører til det samme<br />
resultat. Disse gener går under navnet tumorsuppressorgener.<br />
Som navnet antyder, er de "svulstundertrykkere", og<br />
det vil sige, at generne er i stand til at redde cellerne fra at<br />
blive til kræftceller.<br />
TGF-β – en af cellens bremser<br />
Et eksempel på en tumorsuppressor-signalkæde er den, som<br />
antivækstfaktoren transforming growth factor β (TGF-β) bruger,<br />
når den binder til sin respektive receptor, TGF-β-receptoren.<br />
Men selvom det er et godt eksempel, har antivækstfaktoren<br />
et dårligt navn, og her tænkes på "growth factor".<br />
For i stedet for at sætte cellen i gang med at dele sig, som en<br />
klassisk "growth factor" gør det, gør den det modsatte. Når<br />
antivækstfaktoren binder til sin receptor, sender den et signal<br />
ind i cellen om, at den ikke skal dele sig.<br />
En bremsende signalkæde med få stationer<br />
Ofte ser man, at et signal fra en receptor skal igennem mange<br />
stationer, før det ender i cellekernen. Det vil sige, at der<br />
er mange proteiner involveret i at modtage og sende signalet<br />
videre. Men i TGF-β’s tilfælde ser det ud til, at der er meget<br />
få stationer (figur 4.15).<br />
TGF-β-receptoren er en kinase, og den aktiverende binding<br />
af TGF-β-antivækstfaktoren gør receptoren i stand til at<br />
sætte fosfatgrupper på to proteiner, Smad2 og Smad3, som<br />
ligger i receptorens umiddelbare nærhed. Det giver de to proteiner<br />
en strukturændring, som gør dem i stand til at gå sammen<br />
med et tredje protein, Smad4. Herefter bevæger proteinkomplekset<br />
bestående af Smad2, Smad3 og Smad4 sig fra<br />
cytoplasmaet og ind i cellekernen, hvor det binder sig til en<br />
transkriptionsfaktor og aktiverer den. Den aktive transkriptionsfaktor<br />
sørger for, at særlige gener bliver aflæst og gjort til<br />
protein. Dannelsen af proteinet, p15, er særlig vigtig i denne<br />
sammenhæng, fordi proteinet er i stand til at stoppe cellens<br />
deling.<br />
Kræftceller mister bremsen<br />
Men tilfældige mutationer i de tumorsuppressor-gener, som<br />
koder for proteinerne i TGF-β´s signalkæde, kan sætte den<br />
bremsende effekt ud af kraft. Her følger eksempler på, hvordan<br />
det i kræftceller kan gå galt forskellige steder i signalkæden.<br />
Mavekræftceller uden antivækstreceptor<br />
I nogle former for mavekræft har en tilfældig mutation ramt<br />
det gen, som koder for TGF-β-receptoren. Mutationen gør, at<br />
genet pludselig bliver inaktivt og derfor ikke omsat til protein.<br />
Fra at have TGF-β-receptorer på cellens overflade vil cellen<br />
pludselig stå uden. Det vil sige, at der på cellens overflade<br />
ikke er nogen receptor til at tage imod antivækstsignalet, og<br />
cellen vil derfor ignorere beskeden fra nabocellen (se figur<br />
4.16 og kræftcelle A i figur 4.17).
Kræftceller uden Smad2 eller 4 mister også bremsen<br />
Det tyder dog på, at det i langt højere grad er de tumorsuppressorer,<br />
der "gemmer" sig i cellens indre, som bliver udsat<br />
for ødelæggende mutationer. Både Smad2 og Smad4 er i<br />
mange forskellige kræftformer enten defekte, eller også er<br />
deres respektive gener blevet inaktiveret, så proteinerne ikke<br />
bliver dannet. Det gør kræftcellerne ude af stand til at videresende<br />
antivækstsignalet fra receptoren, og det vil sige, at<br />
de proteiner, som Smad2 og Smad4 i samarbejde styrer dannelsen<br />
af, og som skulle sørge for at stoppe cellens deling,<br />
ikke bliver lavet (kræftcelle B i figur 4.17). Med andre ord:<br />
Kræftcellen får lov til at dele sig ukontrolleret, fordi to<br />
tumorsuppressorer, enten Smad2 eller Smad4, af forskellige<br />
årsager er sat ud af funktion.<br />
Signalkædens slutprodukt savnes<br />
I nogle kræftceller er det ikke selve signalkæden, det er gået<br />
ud over, men derimod slutproduktet af et antivækstsignal –<br />
på figur 4.15 er slutproduktet p15. Normalt ender signalkæden<br />
med, at nye proteiner bliver dannet på antivækstsignalets<br />
kommando – deriblandt p15 – som stopper cellen i sin<br />
deling. Men i mange forskellige kræftformer har kræftcellerne<br />
mistet p15-genet, og det vil sige, at p15-proteinet ikke bliver<br />
dannet (kræftcelle C i figur 4.17). I de tilfælde vil der ikke<br />
være noget til at stoppe cellens deling, og den vil derfor kunne<br />
dele sig uhæmmet.<br />
Onkogener og tumorsuppressorer<br />
Figur 4.15. Uden TGF-β er<br />
receptoren og de proteiner, som<br />
er ansvarlige for at videresende<br />
antivækstsignalet, inaktive. Hvis<br />
cellen modtager TGF-β fra en<br />
anden celle, bliver receptoren<br />
aktiv. Receptoren aktiverer derefter<br />
de "sovende" proteiner,<br />
som ender i cellekernen, hvor<br />
nydannet p15-protein sørger for<br />
at stoppe cellens deling.<br />
Figur 4.16. Hvis en normal<br />
celle modtager antivækstfaktorer<br />
fra en nabocelle, stopper<br />
den sin deling (parakrin signalering).<br />
Nogle kræftceller mangler<br />
receptorerne, som skal tage<br />
imod antivækstfaktorerne. På<br />
den måde ignorerer kræftcellerne<br />
beskeden og fortsætter ufortrødent<br />
deres deling.<br />
39
Figur 4.17. Tre eksempler på, hvordan kræftceller kan miste deres bremse. Det første billede viser, hvordan bremsen virker i en normal celle: antivækstfaktoren<br />
TGF-β’s binding til receptoren får Smad2, Smad3 og Smad4 til at danne et Smad-kompleks. Smad-komplekset virker som en transkriptionsfaktor og sørger<br />
for syntese af p15-proteinet, som bremser cellens deling. De næste tre billeder viser, hvordan kræftceller på tre forskellige måder kan miste deres bremse<br />
og med det samme resultat: Celledelingerne kan ikke stoppes. I kræftcelle A har cellen mistet TGF-β-receptoren og kan derfor ikke modtage antivækstsignalet.<br />
I kræftcelle B mangler cellen Smad4. Det vil sige, at Smad-komplekset vil mangle en enhed (Smad4), og Smad-komplekset kan derfor ikke udføre<br />
sin funktion. I kræftcelle C mangler p15-genet, og det vil sige, at p15-proteinet ikke kan dannes på antivækstfaktorens kommando.<br />
Kan man genoprette balancen?<br />
Det ultimative mål med kræftforskning er selvfølgelig at<br />
finde ud af, hvordan man kan komme af med sygdommen.<br />
Man kunne f.eks. forestille sig, at den efterhånden meget<br />
detaljerede viden om, hvad det er, der får kræftcellen til at<br />
40 Bogen om kræft<br />
miste kontrollen over sine delinger, kunne bruges i en behandlingssammenhæng.<br />
Sagt med andre ord: Vil det være<br />
muligt at genskabe balancen i kræftcellerne – enten ved at<br />
"lette" cellen for de vedvarende signaler, som fortæller, at<br />
cellen skal dele sig – eller ved at genskabe modvægten i<br />
Figur 4.18.<br />
Normale celler: skal/skal ikke<br />
Normale celler modtager altså signaler, som fortæller cellen, at den skal<br />
dele sig, og andre signaler, som fortæller det modsatte, at den skal lade<br />
være. Udfaldet afhænger af, hvilket signal der er stærkest. Billedligt kunne<br />
man illustrere det med en gammel købmandsvægt, hvor vækstfaktorerne<br />
blev placeret på den ene tallerken og antivækstfaktorerne på den anden.<br />
Hvis der er flere vækstfaktorer end antivækstfaktorer, ville vækstfaktorerne<br />
"veje" mest, og cellen ville dele sig. Omvendt: Hvis der var flere antivækstfaktorer<br />
end vækstfaktorer, ville vægten tippe til antivækstfaktorernes<br />
fordel, og cellen ville ikke dele sig.<br />
Kræftceller: skal<br />
I kræftceller er vægten ude af balance, allerede før "vægt-lodderne" bliver<br />
lagt på. Her hælder vægten til den side, som fortæller, at cellen skal dele<br />
sig. Kræftcellen har defekte proteiner, som hele tiden fortæller, at cellen<br />
skal dele sig, selvom den ikke har modtaget vækstfaktorer. Samtidig er<br />
defekte proteiner ude af stand til at videresende antivækstsignaler, og de er<br />
derfor ikke en modvægt til vækstfaktorerne. Denne ubalance giver sig<br />
udslag i, at cellerne deler sig ukontrolleret.
form af et nyt antivækstsignal. Begge tilfælde ville – på hver<br />
deres måde – kunne bremse kræftcellens ukontrollable delinger.<br />
Lad det være sagt med det samme: Der er ikke nogle af de<br />
etablerede behandlinger, man bruger i dag, som gør brug af<br />
denne nye viden. Men kræftforskere og læger over hele verden<br />
arbejder intenst på at udvikle nye behandlinger, som<br />
Kan man slukke for Ras?<br />
Mange forskere har været tiltrukket af ideen om at "slukke"<br />
for de proteiner, som er produkter af onkogener, og som<br />
konstant fortæller cellen, at den skal dele sig. Hvis man<br />
kunne slukke for disse proteiner, kunne man måske bremse<br />
kræftcellers ukontrollable delinger.<br />
En fjerdedel af alle kræfttilfælde har Ras-onkogenet<br />
I denne sammenhæng har Ras længe været i forskernes<br />
søgelys, fordi Ras-genet udmærker sig ved at være det<br />
proto-onkogen blandt alle kræftformer, som hyppigst er<br />
udsat for en kræftfremkaldende mutation. 25 pct. af alle<br />
kræfttilfælde indeholder en muteret udgave af Ras-genet.<br />
Mutationen betyder, at Ras-proteinet konstant er i sin aktive<br />
form og derfor udsender et vedvarende signal om, at<br />
cellen skal dele sig. Hvis man så – teoretisk set – var i<br />
stand til at slukke for det konstant aktive Ras, ville kræftcellen<br />
også miste det vedvarende signal om at dele sig,<br />
og så ville kræftcellen gå i stå.<br />
Hvordan kan man så slukke for Ras-proteinet? Et potentielt<br />
svar på dette spørgsmål fik man, da forskere begyndte<br />
at forstå, hvordan Ras-proteinet bliver lavet i cellen.<br />
Det var samtidig startskuddet til udviklingen af en ny<br />
Figur 4.19. Før behandling: Ved at sætte en farnesylgruppe på Ras-proteinet<br />
sørger farnesyl-transferasen for, at Ras ender på sin rette plads,<br />
nemlig på indersiden af cellemembranen. Herfra (og kun herfra) udsender<br />
proteinet et konstant vækstsignal, som ender med, at cellen deler sig.<br />
sætter ind lige netop der, hvor kræftcellerne adskiller sig fra<br />
normale celler. Hvis man kigger forskerne over skulderen og<br />
ser, hvad der foregår i laboratorierne, behøver gennembruddet<br />
ikke have lange udsigter. Det er "Kan man slukke for Ras"<br />
herunder et klart eksempel på.<br />
kræftbehandling, som i skrivende stund bliver afprøvet i<br />
videnskabelige forsøg med patienter.<br />
Fedtmolekyle hæfter Ras fast til cellemembranen<br />
Forskerne fandt ud af, at når proteinet er færdiglavet ud<br />
fra sit mRNA, findes det ikke på sin rette plads på indersiden<br />
af cellemembranen, men derimod i cellens cytoplasma.<br />
Det gør Ras-proteinet ude af stand til at udføre<br />
sin funktion. Det "umodne" Ras-protein kræver nemlig at<br />
få påsat et fedtmolekyle (en farnesylgruppe), som gør proteinet<br />
i stand til at "hæfte" sig fast på cellemembranens<br />
inderside. Det kalder man at farnesylere, og enzymet,<br />
som er ansvarlig for påsætningen, er en farnesyl-transferase,<br />
som katalyserer reaktionen mellem Ras og farnesylgruppen.<br />
Dur Ras uden fedtmolekyle?<br />
Forskerne fik en genial ide: Hvis de kunne udvikle et stof,<br />
som var i stand til at hæmme farnesyl-transferasen (og dermed<br />
påsætningen af farnesyl på Ras-proteinet), ville Rasproteinet<br />
aldrig nå frem til cellemembranen og derfor<br />
være ude af stand til at udføre sin funktion. Det lykkedes,<br />
og forskerne udviklede hurtigt indtil flere stoffer, som var<br />
i stand til at hæmme farnesyl-transferasens aktivitet.<br />
Efter behandling: Hæmmeren gør farnesyl-transferasen ude af<br />
stand til at sætte farnesyl på Ras. Ras-proteinet kommer derfor ikke<br />
til membranen og kan derfor ikke sætte et vækstsignal i gang.<br />
Kræftcellen deler sig ikke.<br />
Onkogener og tumorsuppressorer<br />
41
Det virker i celler uden for kroppen<br />
Med den bedste farnesyl-transferase-hæmmer i hånden,<br />
begav forskerne sig ud på den lange vej, som skulle til for<br />
at vurdere, om stoffet havde den rette virkning. I cellekulturer<br />
(in vitro) så de, at hæmmeren var i stand til at forhindre<br />
Ras i at nå frem til cellemembranen.<br />
I det næste trin så de, at hæmmeren var i stand til at<br />
forhindre, at normale celler fik kræftlignende egenskaber,<br />
når de modtog en ellers kræftfremkaldende udgave af<br />
Ras-proteinet. Disse resultater opmuntrede dem til at gå<br />
videre i forsøgsrækken. De begyndte at teste hæmmeren<br />
på mus.<br />
Det virker i mus<br />
I første omgang så de på, om hæmmeren var i stand til at<br />
hindre kræftsvulster i at blive dannet. De sprøjtede celler,<br />
som indeholdt en kræftfremkaldende udgave af Ras, ind<br />
under huden på mus. Og de så, hvad de gerne ville se:<br />
De mus, som ikke modtog hæmmeren, udviklede kræftsvulster,<br />
hvor cellerne var sprøjtet ind. De mus, som<br />
modtog hæmmeren, dannede ikke kræftsvulster.<br />
42 Bogen om kræft<br />
En målrettet behandling uden bivirkninger<br />
I de omtalte museforsøg fik de også en ide om, at behandlingen<br />
var meget målrettet. Brugen af hæmmeren<br />
havde ikke nogen virkning på de normale celler, og det<br />
tyder på, at behandlingen kun rammer kræftcellerne. Her<br />
adskiller behandlingen sig fra de traditionelle behandlinger<br />
– kemo eller stråler – hvor de normale celler også bliver<br />
ramt. Det giver bivirkninger, der gør, at man ofte må stoppe<br />
behandlingen i utide.<br />
Fra mus til mennesker<br />
Disse forsøg har opmuntret forskerne til at afprøve hæmmeren<br />
på kræftpatienter med fremskreden sygdom, hvor<br />
kræftcellerne indeholder muteret Ras. Patienterne indtager<br />
hæmmeren i pilleform flere gange om dagen i en længere<br />
periode. Samtidig har nye forsøg i mus vist, at hvis man<br />
kombinerer behandlingen med kemoterapi, bliver resultatet<br />
endnu bedre. Tiden vil vise, om man kan opnå de<br />
samme resultater hos mennesker.<br />
Figur 4.20. Mus, som ikke modtager hæmmer af farnesyl-transferase,<br />
udvikler kræftsvulster der, hvor kræftcellerne sprøjtes ind. Mus, som<br />
modtager hæmmer af farnesyl-transferase, udvikler ikke kræftsvulster.<br />
Ikke nok med at de var i stand til at hæmme dannelsen af kræftsvulster<br />
– forskerne så også, at hæmmeren kunne få allerede eksisterende<br />
kræftsvulster til at stoppe deres vækst og blive mindre.