1 Menneskets genom
1 Menneskets genom
1 Menneskets genom

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
18209 01.fm7 Page 47 Friday, March 3, 2006 12:37 PM<br />
Tabel 1.8 Eksempler på epigenetiske modifikationer<br />
til regulation af gen-ekspressionen.<br />
Alternativ transkription og processering<br />
alternativ splejsning meget hyppig<br />
alternativ anvendelse af promotorer almindelig<br />
alternativ polyadenylering almindelig<br />
RNA-editering<br />
Genomiske modifikationer<br />
sjælden<br />
metylering/demetylering af DNA meget hyppig<br />
acetylering/deacetylering af histon<br />
RNA-interferens<br />
meget hyppig<br />
nedbrydning af mRNA almindelig<br />
motorer og differentiel RNA-processering og<br />
der kan således ud fra et relativt lille antal gener<br />
dannes et stort antal forskellige isoformer af<br />
genprodukter. Disse opdagelser har ændret den<br />
klassiske definition af et gen, og den simple relation<br />
»ét gen – ét polypeptid« er ikke mere<br />
gældende.<br />
Mange gener har to eller flere alternative promotorer<br />
hvilket kan resultere i dannelsen af forskellige<br />
isoformer af genprodukter med forskellige<br />
egenskaber. Et af de bedst undersøgte gener,<br />
hvor differentiel promotor anvendes, er<br />
Genomisk regulation af gen-ekspressionen<br />
det store dystrofin-gen (DMD), som består af<br />
79 exons fordelt på 2,4 Mb. Mindst 8 forskellige<br />
promotorer er beskrevet (Figur 1.31), og de<br />
anvendes til celletypespecifik ekspression af<br />
dystrofin-genet. Atter andre isoformer af dystrofin-genet<br />
skyldes alternativ splejsning.<br />
Omkring halvdelen af menneskets gener har<br />
primære transkripter der undergår alternativ<br />
splejsning, hvor forskellige exon-kombinationer<br />
bliver inkluderet i det færdige transkript<br />
ved RNA-processering (Figur 1.17 og 1.18).<br />
Det har vist sig at visse transkripter har bestemte<br />
exon-kombinationer i forskellige væv. De forskellige<br />
isoformer i de forskellige væv giver<br />
mange muligheder for ændrede funktionelle<br />
egenskaber, men detaljeret viden herom mangler<br />
endnu. Alternativ splejsning har vist sig at<br />
være en af de vigtigste mekanismer bag dannelsen<br />
af forskellige isoformer.<br />
Alternativ polyadenylering er en anden almindelig<br />
måde til dannelse af isoformer. Et eksempel<br />
herpå er alternativ polyadenylering af<br />
calcitonin-transkriptet, hvilket resulterer i<br />
vævsspecifik ekspression af to isoformer (Figur<br />
1.32). Calcitonin er et cirkulerende Ca 2+ -homøostatisk<br />
hormon som produceres i gl. thyroidea.<br />
Det calcitonin-gen-relaterede peptid<br />
L C M P R CNS S G<br />
L1 C1 M1 P1<br />
0 500 1000 1500 2000<br />
2 5 10 15 20 30 40 45 50 55 60 70 79<br />
Dp427 Dp260 Dp140 Dp116 Dp71<br />
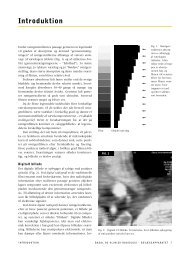
Figur 1.31 Mindst 8 forskellige promotorer benyttes til cellespecifik ekspression af dystrofin-genet. Positionen af de<br />
8 alternative promotorer er vist øverst: L i lymfocytter, C i hjernebarken (cortex cerebri), M i muskel, P i Purkinjefibre,<br />
R i retina, CNS i central nervesystemet, S i schwannske celler, G for generel promotor. Exons er nummererede 1-79.<br />
Notér at promotorvalget afgør hvilken exon der bliver den første i det pågældende transkript (L1, C1, M1, P1, R1 osv.).<br />
Dp427, Dp260, Dp140, Dp116, Dp71 refererer til størrelsen af færdige peptid i kDa, eksempelvis 427 kDa for Dp427.<br />
kb<br />
47